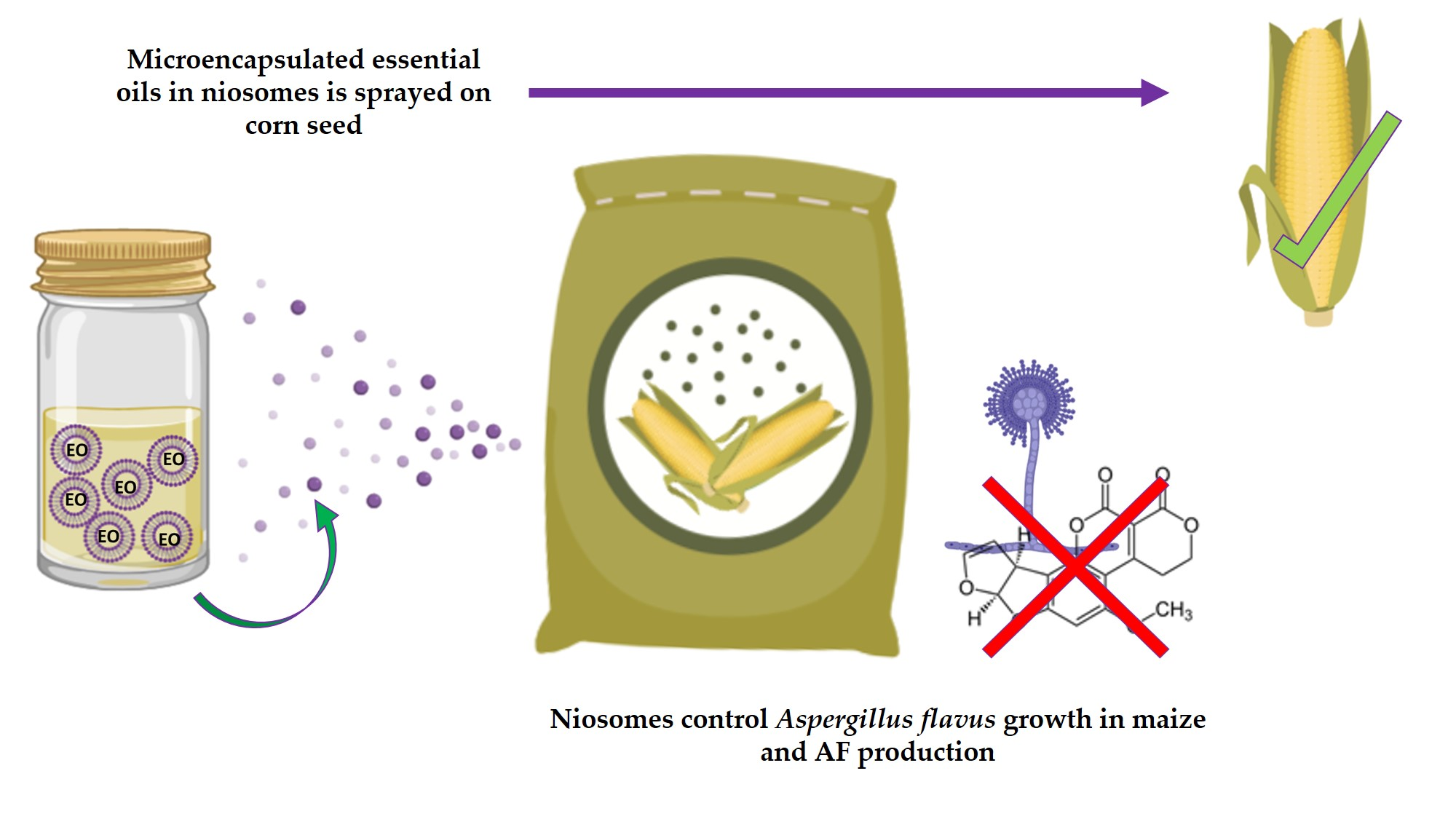
A Novel Niosome-Encapsulated Essential Oil Formulation to Prevent Aspergillus flavus Growth and Aflatoxin Contamination of Maize Grains During Storage




Abstract
:
1. Introduction
2. Results
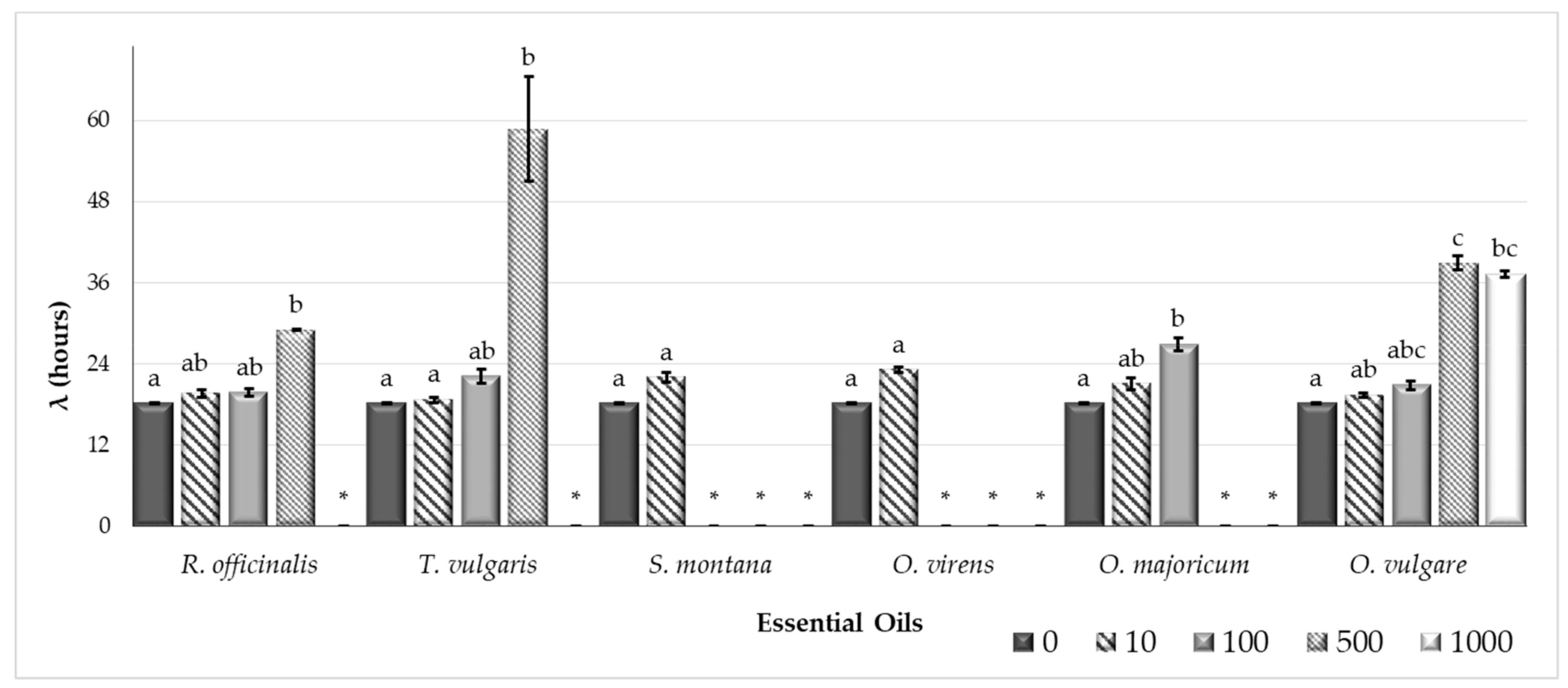
2.1. The Efficacy of Plant Essential Oils Against Fungal Growth and Mycotoxin Production
2.2. Techniques for the Application of Essential Oils to Prevent Fungal Growth and Mycotoxin Production
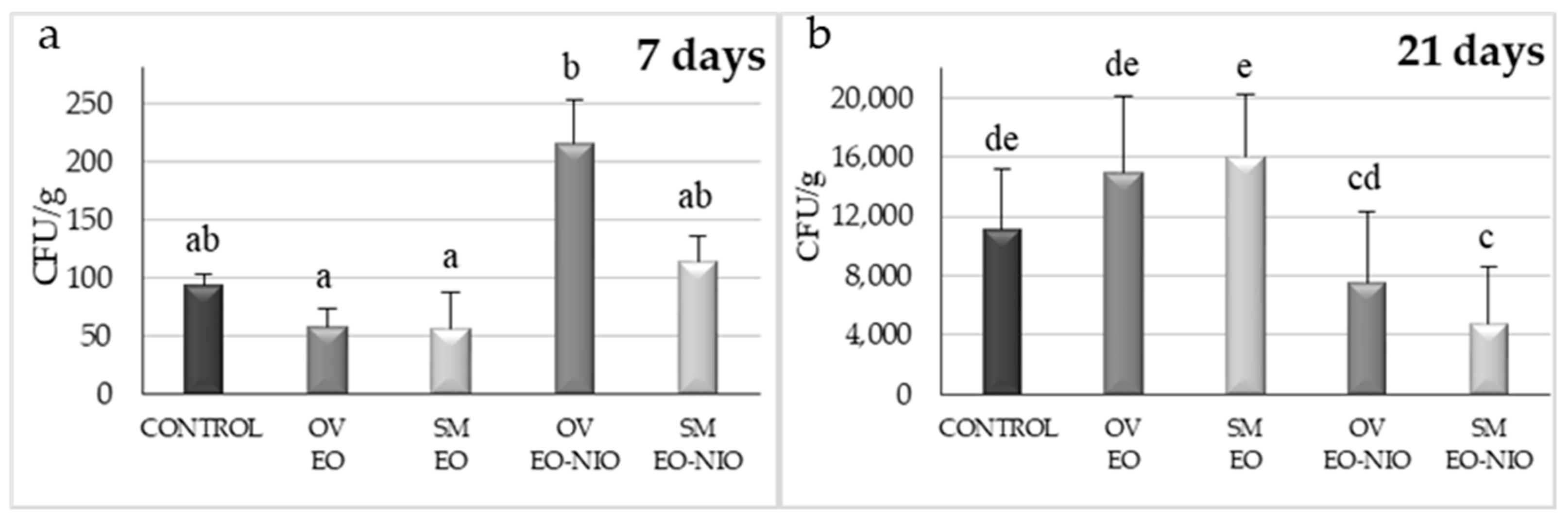
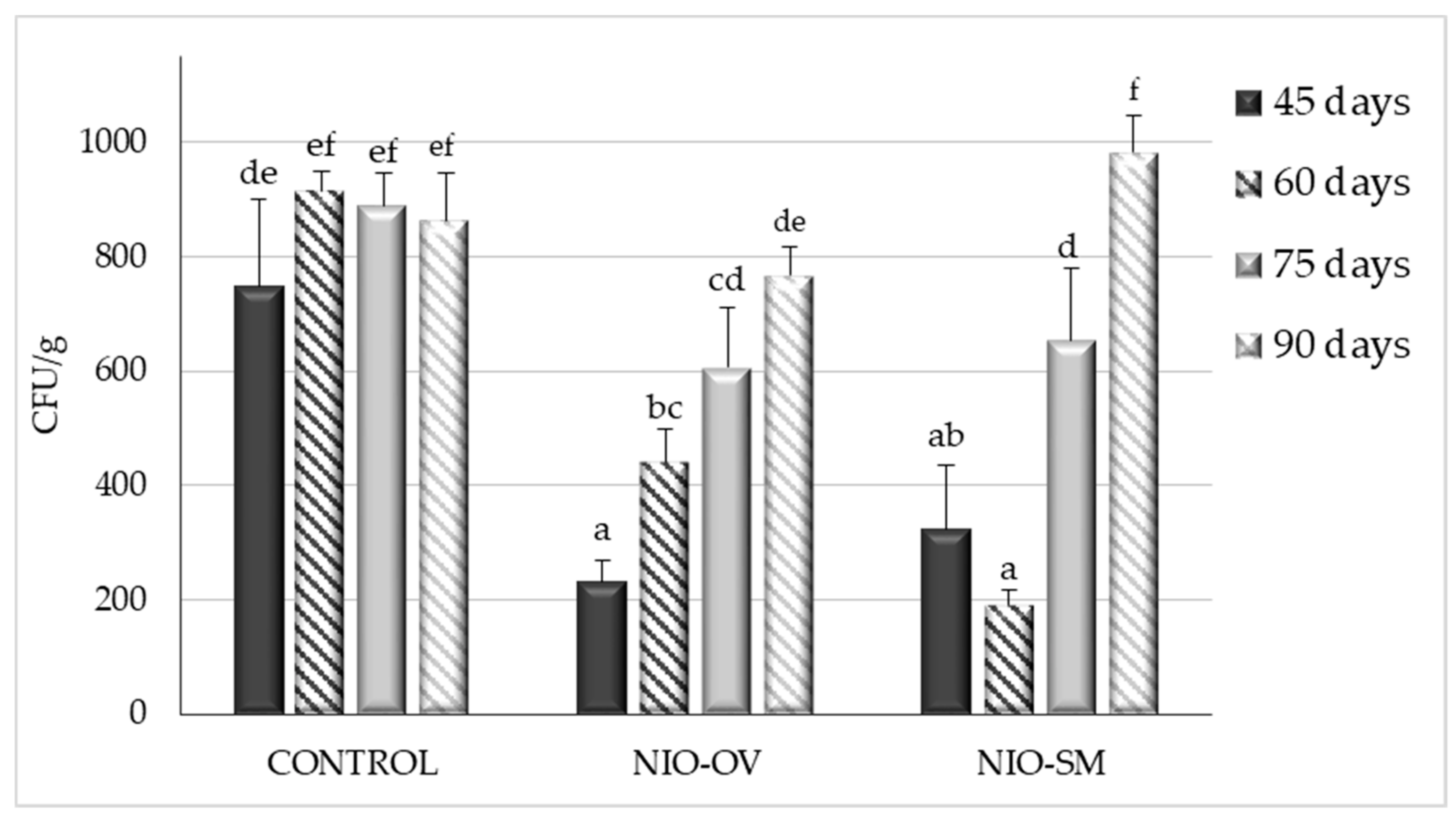
2.2.1. Small-Scale Assay
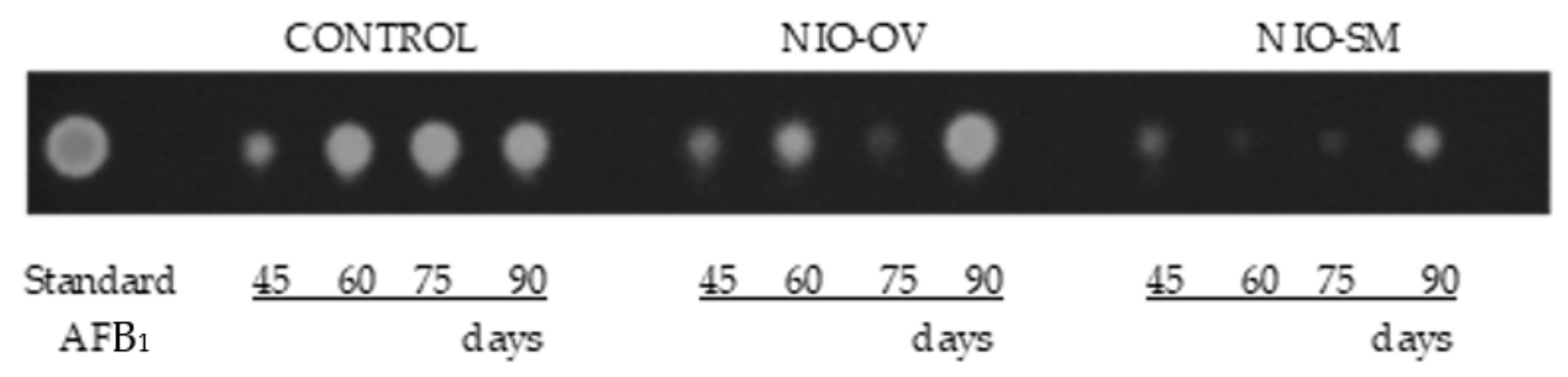
2.2.2. Polypropylene Woven Bags Assays
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Fungal Strains and Essential Oils
5.2. Effectiveness of Plant Essential Oils on Fungal Growth and Aflatoxin Production
5.3. Effect of Satureja Montana and Origanum Virens Essential Oils Encapsulated in Niosomes on Fungal Growth and Aflatoxin Contamination
5.3.1. Procedure for Microencapsulation of Essential Oils
5.3.2. Effect of Niosome-Encapsulated Essential Oils on Fungal Growth and Aflatoxin Production on Maize Grains
Small-Scale Assays
Polypropylene Woven Bag Assays
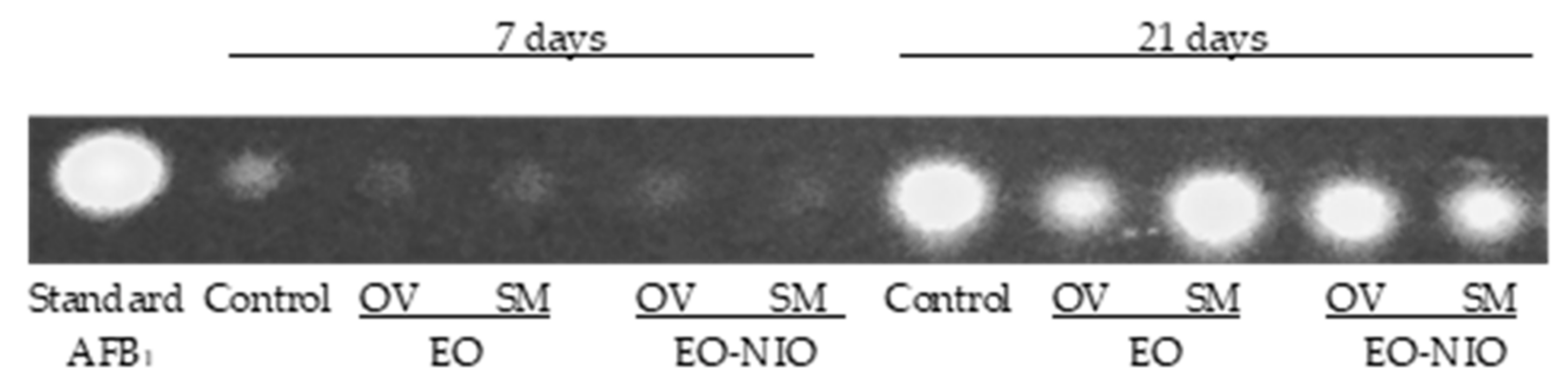
5.4. Detection of Mycotoxins
5.4.1. Detection of Mycotoxins by High Performance Liquid Chromatography (HPLC)
5.4.2. Detection of Mycotoxins by Thin Layer Chromatography
5.5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kensler, T.W.; Roebuck, B.D.; Wogan, G.N.; Groopma, J.D. Aflatoxin: A 50-year odyssey of mechanistic and translational toxicology. Toxicol. Sci. 2011, 120, S25–S48. [Google Scholar] [CrossRef] [PubMed]
- Alshannaq, A.; Yu, J.H. Occurrence, toxicity, and analysis of major mycotoxins in food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Stacy, S.L.; Kensler, T.W. Global risk assessment of aflatoxins in maize and peanuts: Are regulatory standards adequately protective? Toxicol. Sci. 2013, 135, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Gil-Serna, J.; Vázquez, C.; Patiño, B. Mycotoxins/Toxicology. In Encyclopedia of Food Microbiology; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Food and Agriculture Organization of the United Nations, Statistic Division. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 18 September 2019).
- Battilani, P.; Toscano, P.; van der Fels-Klerx, H.J.; Jeggieri, M.C.; Brera, C.; Rortais, A.; Goumperis, T.; Robinson, T. Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci. Rep. 2016, 6, 24328. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Regulation N° 165/2010 amending Regulation (EC) No 1881/2006 setting maximum levels for certain contaminants in foodstuffs as regards aflatoxins. Off. J. Eur. Union 2010, 50, 8–12. [Google Scholar]
- Hussein, H.S.; Brasel, J.M. Toxicity, metabolism, and impact of mycotoxins on humans and animals. Toxicology 2001, 167, 101–134. [Google Scholar] [CrossRef]
- Chulze, S. Strategies to reduce mycotoxin levels in maize during storage: A review. Food Addit. Contam. 2010, 27, 651–657. [Google Scholar] [CrossRef]
- Lagogianni, C.S.; Tsisigiannis, D.I. Effective chemical management for prevention of aflatoxins in maize. Phytopathol. Mediterr. 2018, 57, 186–197. [Google Scholar]
- Ji, C.; Fan, J.; Zhao, L. Review on biological degradation of mycotoxins. Anim. Nutr. 2016, 2, 127–133. [Google Scholar] [CrossRef]
- Da Cruz Cabral, L.; Pinto, V.F.; Patriarca, A. Application of plant derived compounds to control fungal spoilage and mycotoxin production in foods. Int. J. Food Microbiol. 2013, 166, 1–14. [Google Scholar] [CrossRef]
- Pandey, A.K.; Kumar, P.; Singh, P.; Tripathi, N.N.; Bajpai, V.K. Essential oils: Sources of antimicrobials and food preservatives. Front. Microbiol. 2017, 7, 2161. [Google Scholar] [CrossRef] [PubMed]
- Kumar-Dwivedy, A.; Kumar, M.; Updhyay, N.; Prakash, B.; Kishore-Dubey, N. Plant essential oils against food borne fungi and mycotoxins. Curr. Opin. Food Sci. 2016, 11, 16–21. [Google Scholar] [CrossRef]
- Da Silva, N.; Polis, L.; Faggion, J.; Yumie, C.; Galerani, S.A.; Grespan, R.; Botiao, S.; Augusto, C.; Abreu, B.A.; Machinski, M. Antifungal activity and inhibition of fumonisin production by Rosmarinus officinalis L. essential oil in Fusarium verticillioides (Sacc.) Nirenberg. Food Chem. 2015, 166, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Esper, R.H.; Gonçalez, E.; Marques, M.O.M.; Felicio, R.C.; Felicio, J.D. Potential of essential oils for protection of grains contaminated by aflatoxin produced by Aspergillus flavus. Front. Microbiol. 2014, 5, 269. [Google Scholar] [CrossRef] [PubMed]
- Kedia, A.; Kumar-Dwivedy, A.; Kumar-Jha, D.; Dubey, N.K. Efficacy of Mentha spicata essential oil in suppression of Aspergillus flavus and aflatoxin contamination in chickpea with particular emphasis to mode of antifungal action. Protoplasma 2016, 253, 647–653. [Google Scholar] [CrossRef]
- Ribeiro-Santos, R.; Andrade, M.; Sanches-Silva, A. Application of encapsulated essential oils as antimicrobial agents in food packaging. Food Sci. 2017, 14, 78–84. [Google Scholar] [CrossRef]
- Mäes, C.; Bouquillon, S.; Fauconnier, M.L. Encapsulation of essential oils for the development of biosourced pesticides with controlled release: A review. Molecules 2019, 24, 2539. [Google Scholar] [CrossRef]
- Donsí, F.; Annunziata, M.; Sessa, M.; Ferrari, G. Nanoencapsulation of essential oils to enhance their antimicrobial activity in foods. LWT Food Sci. Technol. 2011, 44, 1908–1914. [Google Scholar] [CrossRef]
- Nesci, A.; Passone, M.A.; Barra, P.; Girardi, N.; García, D.; Etcheverry, M. Prevention of aflatoxin contamination in stored grains using chemical strategies. Curr. Opin. Food Sci. 2016, 11, 56–60. [Google Scholar] [CrossRef]
- Da Silva, P.T.; Fries, L.L.M.; de Menezes, C.R.; Holken, A.T.; Schwan, C.L.; Wigmann, E.F.; Bastos, J.D.O.; da Silva, C.D.B. Microencapsulation: Concepts, mechanisms, methods and some applications in food technology. Cienc. Rural 2014, 44, 1304–1311. [Google Scholar] [CrossRef]
- Prakash, B.; Kujur, A.; Yadav, A.; Kumar, A.; Singh, P.P.; Dubey, N.K. Nanoencapsulation: An efficient technology to boost the antimicrobial potential of plant essential oils in food system. Food Control 2018, 89, 1–11. [Google Scholar] [CrossRef]
- Amoabediny, G.; Haghiralsadat, F.; Naderinezhad, S.; Helder, M.N.; Kharanaghi, E.A.; Arough, J.M.; Zandieh-Doulabi, B. Overview of preparation methods of polymeric and lipid-based (niosome, solid lipid, liposome) nanoparticles: A comprehensive review. Int. J. Polym. Mater. Polym. Biomater. 2018, 67, 383–400. [Google Scholar] [CrossRef]
- Císarová, M.; Tancinová, D.; Medo, J.; Kacaniová, M. The in vitro effect of selected essential oils on the growth and mycotoxin production of Aspergillus species. J. Environ. Sci. Health Part B 2016, 51, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Perczak, A.; Gwiazdowska, D.; Marchwinska, D.; Jus, K.; Gwiazdowski, R.; Waskiewicz, A. Antifungal activity of selected essential oils against Fusarium culmorum and F. graminearum and their secondary metabolites in wheat seeds. Arch. Microbiol. 2019, 201, 1085–1097. [Google Scholar] [CrossRef]
- Wang, L.; Jiang, N.; Wang, D.; Wang, M. Effects of essential oil citral on the growth, mycotoxin biosynthesis and transcriptomic profile of Alternaria alternate. Toxins 2019, 11, 553. [Google Scholar] [CrossRef]
- Prakash, B.; Kedia, A.; Mishra, P.K.; Dubey, N.K. Plant essential oils as food preservatives to control moulds, mycotoxin contamination and oxidative deterioration of agri-food commodities—Potentials and challenges. Food Control 2015, 47, 381–391. [Google Scholar] [CrossRef]
- Abbaszadeh, S.; Sharifzadeh, A.; Shokri, H.; Khosravi, A.R.; Abbaszadeh, A. Antifungal efficacy of thymol, carvacrol, eugenol and menthol as alternative agents to control the growth of food-relevant fungi. J. Mycol. Med. 2014, 24, 51–56. [Google Scholar] [CrossRef]
- Gao, T.; Zhou, H.; Zhow, W.; Hu, L.; Chen, J.; Shi, Z. The fungicidal activity of thymol against Fusarium graminearum via inducing lipid peroxidation and disrupting ergosterol biosynthesis. Molecules 2016, 21, 770. [Google Scholar] [CrossRef]
- Zeisig, R.; Cammerer, B. Liposomes in the food industry. In Nano-and Microencapsulation for Foods; Vilstrup, P., Ed.; Leatherhead: London, UK, 2001; pp. 101–109. [Google Scholar]
- Maina, A.W.; Wagacha, J.M.; Mwaura, F.B.; Muthomi, J.W.; Woloshuk, C.P. Postharvest practices of maize farmers in Kaiti district, Kenya and the impact of hermetic storage on populations of Aspergillus spp. and aflatoxin contamination. J. Food Res. 2016, 5, 53–66. [Google Scholar] [CrossRef]
- Perricone, M.; Arace, E.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Bioactivity of essential oils: A review on their interaction with food components. Front. Microbiol. 2015, 6, 76. [Google Scholar] [CrossRef]
- González-Salgado, N.; González-Jaén, M.T.; Vázquez, C.; Patiño, B. Highly sensitive PCR-based detection method specific for Aspergillus flavus in wheat flour. Food Addit. Contam. 2008, 25, 758–764. [Google Scholar] [CrossRef] [PubMed]
- Bragulat, M.R.; Abarca, M.L.; Cabañes, F.J. An easy screening method for fungi producing ochratoxin A in pure culture. Int. J. Food Microbiol. 2001, 71, 139–144. [Google Scholar] [CrossRef]
- Bernáldez-Rey, M.V. Desarrollo de Métodos de RT-PCR en Tiempo Real Para la Cuantificación de Mohos Toxigénicos Viables en Alimentos. Ph.D. Thesis, University of Extremadura, Badajoz, Spain, 2016. [Google Scholar]
- Scott, P.M.; Lawrence, J.W.; van Walbeek, W. Detection of mycotoxins by thin-layer chromatography: Application to screening of fungal extracts. Appl. Microbiol. 1970, 20, 839–842. [Google Scholar] [PubMed]
- Gimeno, A.; Martins, M.L. Rapid thin layer chromatography determination of patulin, citrinin, and aflatoxin in apples and pears, and their juices and jams. J. Assoc. Off. Anal. Chem. 1983, 66, 85–91. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EOs | μg/mL | AFB1 (µg/g agar) | AFB2 (µg/g agar) | AFG1 (µg/g agar) | AFG2 (µg/g agar) |
|---|---|---|---|---|---|
| R. officinalis | 0 | 10.754 ± 0.925 c | 0.201 ± 0.021 c | 0.485 ± 0.055 c | 0.088 ± 0.014 b |
| 10 | 5.205 ± 1.033 bc | 0.1 ± 0.022 bc | 0.213 ± 0.045 bc | ND a | |
| 100 | 5.223 ± 0.171 abc | 0.11 ± 0.006 bc | 0.216 ± 0.012 abc | ND a | |
| 500 | 1.09 ± 0.152 ab | 0.017 ± 0.002 ab | 0.058 ± 0.007 ab | ND a | |
| 1000 | ND a | ND a | ND a | ND a | |
| T. vulgaris | 0 | 10.754 ± 0.925 b | 0.201 ± 0.021 b | 0.485 ± 0.055 b | 0.088 ± 0.014 b |
| 10 | 7.04 ± 0.977 b | 0.122 ± 0.019 ab | 0.256 ± 0.059 ab | ND a | |
| 100 | 5.994 ± 0.554 ab | 0.117 ± 0.019 ab | 0.264 ± 0.069 ab | ND a | |
| 500 | ND a | ND a | ND a | ND a | |
| 1000 | ND a | ND a | ND a | ND a | |
| S. montana | 0 | 10.754 ± 0.925 b | 0.201 ± 0.021 b | 0.485 ± 0.055 b | 0.088 ± 0.014 b |
| 10 | 8.27 ± 0.686 ab | 0.151 ± 0.011 ab | 0.314 ± 0.029 ab | ND a | |
| 100 | ND a | ND a | ND a | ND a | |
| 500 | ND a | ND a | ND a | ND a | |
| 1000 | ND a | ND a | ND a | ND a | |
| O. virens | 0 | 10.754 ± 0.925 b | 0.201 ± 0.021 bc | 0.485 ± 0.055 b | 0.088 ± 0.014 b |
| 10 | 10.52 ± 1.334 b | 0.245 ± 0.039 c | 0.508 ± 0.065 b | ND a | |
| 100 | 0.033 ± 0.044 ab | 0.003 ± 0 ab | 0.004 ± 0.002 ab | ND a | |
| 500 | ND a | ND a | ND a | ND a | |
| 1000 | ND a | ND a | ND a | ND a | |
| O. majoricum | 0 | 10.754 ± 0.925 b | 0.201 ± 0.021 ab | 0.485 ± 0.055 b | 0.088 ± 0.014 b |
| 10 | 10.939 ± 0.21 b | 0.234 ± 0.008 b | 0.611 ± 0.057 b | ND a | |
| 100 | 7.999 ± 0.628 ab | 0.186 ± 0.022 ab | 0.375 ± 0.028 ab | ND a | |
| 500 | 0.003 ± 0 a | ND a | ND a | ND a | |
| 1000 | 0.008 ± 0.008 a | ND a | ND a | ND a | |
| O. vulgare | 0 | 10.754 ± 0.925 c | 0.201 ± 0.021 b | 0.485 ± 0.055b | 0.088 ± 0.014 b |
| 10 | 10.143 ± 0.86 bc | 0.192 ± 0.015 b | 0.475 ± 0.056 b | ND a | |
| 100 | 7.867 ± 0.409 abc | 0.168 ± 0.015 ab | 0.466 ± 0.08 | ND | |
| 500 | 0.738 ± 0.08 ab | 0.012 ± 0.002 a | 0.043 ± 0.003 | ND | |
| 1000 | 0.298 ± 0.068 a | 0.004 ± 0.003 a | 0.014 ± 0.004 | ND |
| ZETASICER | NANOSIGHT | ||||
|---|---|---|---|---|---|
| PDI | Z-AVERAGE (nm) | POTENCIAL-ʐ (mV) | Size (nm) | CONCENTRATION (Particle/mL) | |
| O. virens | 0.251 ± 0.019 | 156.2 ± 3.9 | −14.5 ± 0.5 | 142.4 ± 1.0 | (2.96 ± 0.12) × 1014 |
| S. montana | 0.251 ± 0.011 | 153.3 ± 2.8 | −14.6 ± 2.3 | 140.6 ± 3.8 | (1.86 ± 0.07) × 1014 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Díaz, M.; Patiño, B.; Vázquez, C.; Gil-Serna, J. A Novel Niosome-Encapsulated Essential Oil Formulation to Prevent Aspergillus flavus Growth and Aflatoxin Contamination of Maize Grains During Storage. Toxins 2019, 11, 646. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110646
García-Díaz M, Patiño B, Vázquez C, Gil-Serna J. A Novel Niosome-Encapsulated Essential Oil Formulation to Prevent Aspergillus flavus Growth and Aflatoxin Contamination of Maize Grains During Storage. Toxins. 2019; 11(11):646. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110646
Chicago/Turabian StyleGarcía-Díaz, Marta, Belén Patiño, Covadonga Vázquez, and Jessica Gil-Serna. 2019. "A Novel Niosome-Encapsulated Essential Oil Formulation to Prevent Aspergillus flavus Growth and Aflatoxin Contamination of Maize Grains During Storage" Toxins 11, no. 11: 646. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11110646