Bioprospecting Sponge-Associated Marine Cyanobacteria to Produce Bioactive Compounds

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Molecular Characterization of Sponge-Associated Cyanobacteria Strains
2.2. PCR Screening for PKS and NRPS Encoding Genes
2.3. Cyanotoxins
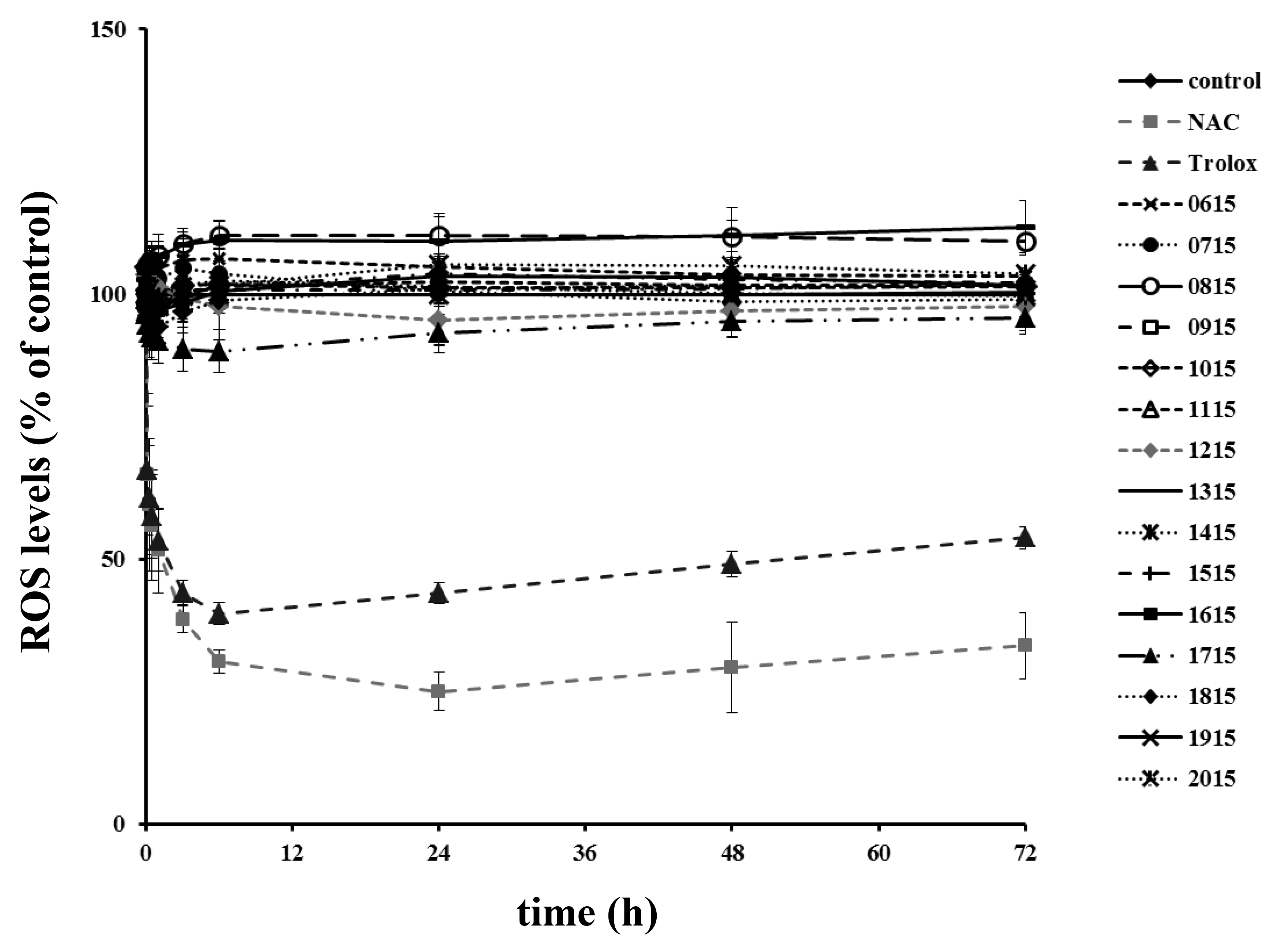
2.4. Antimicrobial Activity
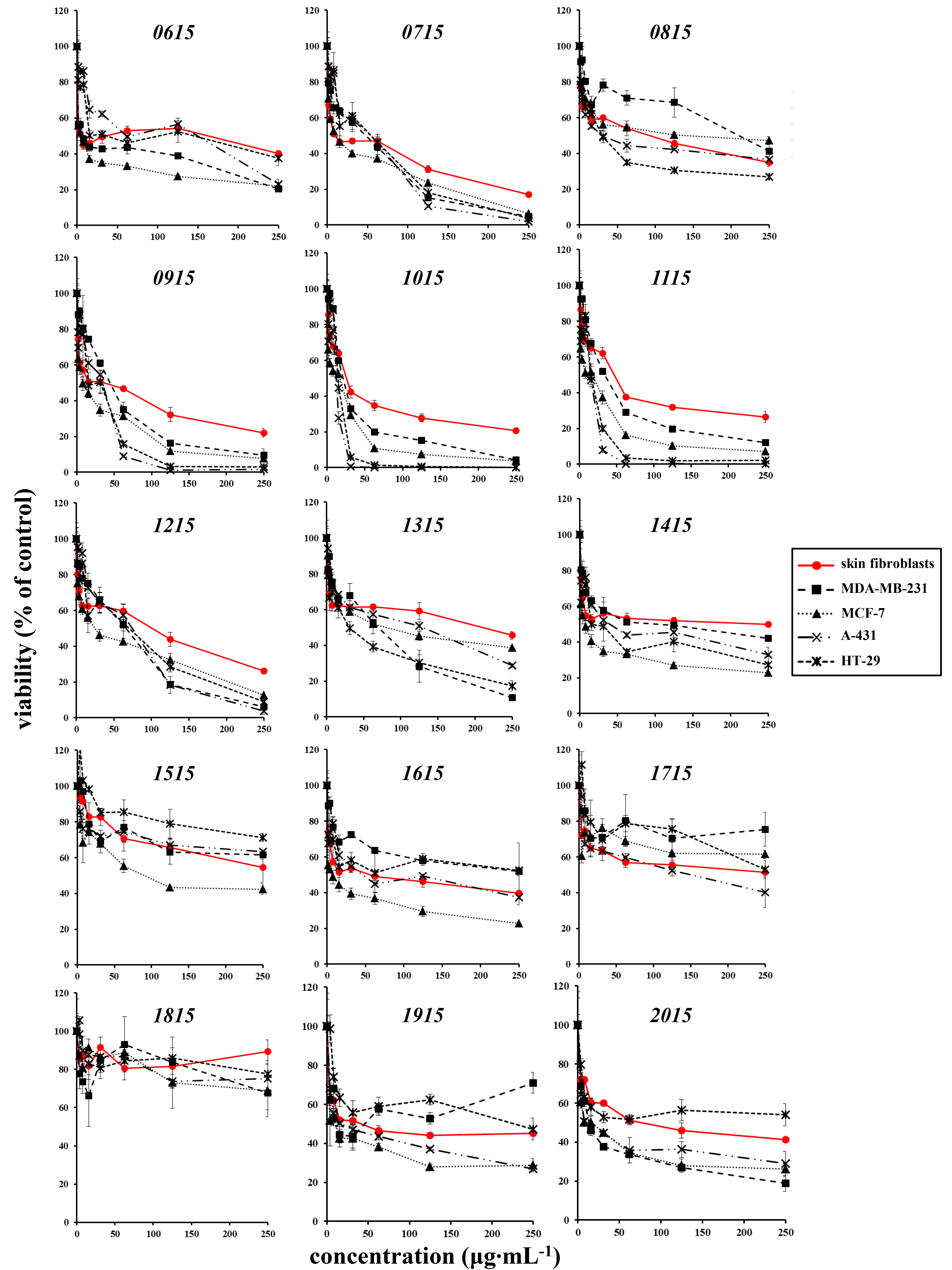
2.5. Effects on Cell Lines
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cyanobacterial Isolates and Culture Conditions
5.2. DNA Extraction, PCR Amplification and Sequencing
5.3. Phylogenetic Analysis
5.4. Cyanotoxin Analysis Using LC-MS/MS
5.5. Extract Preparation for Assays
5.6. Agar Disc Diffusion Assay
5.7. Human Cell Lines and Cell Culture Conditions
5.8. Cytotoxicity Assay
5.9. Estimation of the Antioxidant Potential
5.10. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2013, 37, 23–43. [Google Scholar] [CrossRef]
- Nunnery, J.K.; Mevers, E.; Gerwick, W.H. Biologically active secondary metabolites from marine cyanobacteria. Curr. Opin. Biotechnol. 2010, 21, 787–793. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Parihar, P.; Singh, M.; Bajguz, A.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Uncovering Potential Applications of Cyanobacteria and Algal Metabolites in Biology, Agriculture and Medicine: Current Status and Future Prospects. Front. Microbiol. 2017, 8, 515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.T. Pharmaceutical agents from filamentous marine cyanobacteria. Drug Discov. Today 2013, 18, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Gugger, M.; Sivonen, K.; Fewer, D.P. Natural Product Biosynthetic Diversity and Comparative Genomics of the Cyanobacteria. Trends Microbiol. 2015, 23, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Mi, Y.; Zhang, J.; He, S.; Yan, X. New peptides isolated from marine cyanobacteria, an overview over the past decade. Mar. Drugs 2017, 15, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, M.; Costa-Rodrigues, J.; Fernandes, M.H.; Barros, P.; Vasconcelos, V.; Martins, R. Marine cyanobacteria compounds with anticancer properties: A review on the implication of apoptosis. Mar. Drugs 2012, 10, 2181–2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, M.D.R.; Costa, M. Marine cyanobacteria compounds with anticancer properties: Implication of apoptosis. In Handbook of Anticancer Drugs from Marine Origin; Kim, S.-K., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 621–647. ISBN 9783319071442. [Google Scholar]
- Deng, C.; Pan, B.; O’Connor, O.A. Brentuximab Vedotin. Clin. Cancer Res. 2013, 19, 22–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, D.G.; Duggan, P.S.; Jackson, O. Cyanobacterial symbioses. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: Dordrech, Germany, 2012; pp. 593–636. ISBN 0306468557. [Google Scholar]
- Mehbub, M.; Lei, J.; Franco, C.; Zhang, W. Marine Sponge Derived Natural Products between 2001 and 2010: Trends and Opportunities for Discovery of Bioactives. Mar. Drugs 2014, 12, 4539–4577. [Google Scholar] [CrossRef] [Green Version]
- Thomas, T.; Moitinho-Silva, L.; Lurgi, M.; Björk, J.R.; Easson, C.; Astudillo-García, C.; Olson, J.B.; Erwin, P.M.; López-Legentil, S.; Luter, H.; et al. Diversity, structure and convergent evolution of the global sponge microbiome. Nat. Commun. 2016, 7, 11870. [Google Scholar] [CrossRef] [Green Version]
- Thomas, T.; Rusch, D.; DeMaere, M.Z.; Yung, P.Y.; Lewis, M.; Halpern, A.; Heidelberg, K.B.; Egan, S.; Steinberg, P.D.; Kjelleberg, S. Functional genomic signatures of sponge bacteria reveal unique and shared features of symbiosis. ISME J. 2010, 4, 1557–1567. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, U.; Piel, J.; Degnan, S.M.; Taylor, M.W. Genomic insights into the marine sponge microbiome. Nat. Rev. Microbiol. 2012, 10, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Konstantinou, D.; Gerovasileiou, V.; Voultsiadou, E.; Gkelis, S. Sponges-cyanobacteria associations: Global diversity overview and new data from the Eastern Mediterranean. PLoS ONE 2018, 13, e0195001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konstantinou, D.; Voultsiadou, E.; Panteris, E.; Zervou, S.K.; Hiskia, A.; Gkelis, S. Leptothoe, a new genus of marine cyanobacteria (Synechococcales) and three new species associated with sponges from the Aegean Sea. J. Phycol. 2019, 55, 882–897. [Google Scholar] [CrossRef]
- Regueiras, A.; Alex, A.; Pereira, S.; Costa, M.S.; Antunes, A.; Vasconcelos, V. Cyanobacterial diversity in the marine sponge Hymeniacidon perlevis from a temperate region (Portuguese coast, Northeast Atlantic). Aquat. Microb. Ecol. 2017, 79, 259–272. [Google Scholar] [CrossRef]
- Pagliara, P.; Caroppo, C. Cytotoxic and antimitotic activities in aqueous extracts of eight cyanobacterial strains isolated from the marine sponge Petrosia ficiformis. Toxicon 2011, 57, 889–896. [Google Scholar] [CrossRef]
- Perkerson, R.B.; Johansen, J.R.; Kovácik, L.; Brand, J.; Kaštovský, J.; Casamatta, D.A. A unique pseudanabaenalean (cyanobacteria) genus Nodosilinea gen. nov. based on morphological and molecular data. J. Phycol. 2011, 47, 1397–1412. [Google Scholar] [CrossRef]
- Dadheech, P.K.; Mahmoud, H.; Kotut, K.; Krienitz, L. Haloleptolyngbya alcalis gen. et sp. nov., a new filamentous cyanobacterium from the soda lake Nakuru, Kenya. Hydrobiologia 2012, 691, 269–283. [Google Scholar] [CrossRef]
- Papendorf, O.; Konig, G.M.; Wright, A.D. Hierridin B and 2,4-dimethoxy-6-heptadecyl-phenol, secondary metabolites from the cyanobacterium Phormidium ectocarpi with antiplasmodial activity. Phytochemistry 1998, 49, 2383–2386. [Google Scholar] [CrossRef]
- Freitas, S.; Martins, R.; Costa, M.; Leão, P.N.; Vitorino, R.; Vasconcelos, V.; Urbatzka, R. Hierridin B Isolated from a Marine Cyanobacterium Alters VDAC1, Mitochondrial Activity, and Cell Cycle Genes on HT-29 Colon Adenocarcinoma Cells. Mar. Drugs 2016, 14, 158. [Google Scholar] [CrossRef] [Green Version]
- Leão, P.N.; Costa, M.; Ramos, V.; Pereira, A.R.; Fernandes, V.C.; Domingues, V.F.; Gerwick, W.H.; Vasconcelos, V.M.; Martins, R. Antitumor Activity of Hierridin B, a Cyanobacterial Secondary Metabolite Found in both Filamentous and Unicellular Marine Strains. PLoS ONE 2013, 8, e69562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calteau, A.; Fewer, D.P.; Latifi, A.; Coursin, T.; Laurent, T.; Jokela, J.; Kerfeld, C.A.; Sivonen, K.; Piel, J.; Gugger, M. Phylum-wide comparative genomics unravel the diversity of secondary metabolism in Cyanobacteria. BMC Genom. 2014, 15, 977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, Â.; Gaifem, J.; Ramos, V.; Glukhov, E.; Dorrestein, P.C.; Gerwick, W.H.; Vasconcelos, V.M.; Mendes, M.V.; Tamagnini, P. Bioprospecting Portuguese Atlantic coast cyanobacteria for bioactive secondary metabolites reveals untapped chemodiversity. Algal Res. 2015, 9, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Shih, P.M.; Wu, D.; Latifi, A.; Axen, S.D.; Fewer, D.P.; Talla, E.; Calteau, A.; Cai, F.; Tandeau de Marsac, N.; Rippka, R.; et al. Improving the coverage of the cyanobacterial phylum using diversity-driven genome sequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 1053–1058. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.; Barua, A.; Ruvindy, R.; Savela, H.; Ajani, P.A.; Murray, S.A. The genetic basis of toxin biosynthesis in dinoflagellates. Microorganisms 2019, 7, 222. [Google Scholar] [CrossRef] [Green Version]
- Gkelis, S.; Panou, M.; Konstantinou, D.; Apostolidis, P.; Kasampali, A.; Papadimitriou, S.; Kati, D.; Di Lorenzo, G.M.; Ioakeim, S.; Zervou, S.K.; et al. Diversity, cyanotoxin production, and bioactivities of cyanobacteria isolated from freshwaters of greece. Toxins 2019, 11, 436. [Google Scholar] [CrossRef] [Green Version]
- Gkelis, S.; Fernández Tussy, P.; Zaoutsos, N. Isolation and preliminary characterization of cyanobacteria strains from freshwaters of Greece. Open Life Sci. 2015, 10, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Shishido, T.K.; Jokela, J.; Humisto, A.; Suurnäkki, S.; Wahlsten, M.; Alvarenga, D.O.; Sivonen, K.; Fewer, D.P. The biosynthesis of rare homo-amino acid containing variants of microcystin by a benthic cyanobacterium. Mar. Drugs 2019, 17, 271. [Google Scholar] [CrossRef] [Green Version]
- Zervou, S.K.; Christophoridis, C.; Kaloudis, T.; Triantis, T.M.; Hiskia, A. New SPE-LC-MS/MS method for simultaneous determination of multi-class cyanobacterial and algal toxins. J. Hazard. Mater. 2017, 323, 56–66. [Google Scholar] [CrossRef]
- Meriluoto, J.; Spoof, L.; Codd, G.A. Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; ISBN 1119068681. [Google Scholar]
- Choi, H.; Engene, N.; Smith, J.E.; Preskitt, L.B.; Gerwick, W.H. Crossbyanols A–D, toxic brominated polyphenyl ethers from the hawai’ian bloom-forming cyanobacterium leptolyngbya crossbyana. J. Nat. Prod. 2010, 73, 517–522. [Google Scholar] [CrossRef] [Green Version]
- Zainuddin, E.N.; Jansen, R.; Nimtz, M.; Wray, V.; Preisitsch, M.; Lalk, M.; Mundt, S. Lyngbyazothrins A–D, antimicrobial cyclic undecapeptides from the cultured cyanobacterium Lyngbya sp. J. Nat. Prod. 2009, 72, 2080. [Google Scholar] [CrossRef] [Green Version]
- Martins, R.F.; Ramos, M.F.; Herfindal, L.; Sousa, J.A.; Skærven, K.; Vasconcelos, V.M. Antimicrobial and cytotoxic assessment of marine cyanobacteria—Synechocystis and Synechococcus. Mar. Drugs 2008, 6, 1–11. [Google Scholar] [CrossRef]
- Swain, S.S.; Paidesetty, S.K.; Padhy, R.N. Antibacterial, antifungal and antimycobacterial compounds from cyanobacteria. Biomed. Pharmacother. 2017, 90, 760–776. [Google Scholar] [CrossRef]
- Engene, N.; Rottacker, E.C.; Kaštovský, J.; Byrum, T.; Choi, H.; Ellisman, M.H.; Komárek, J.; Gerwick, W.H. Moorea producens gen. nov., sp. nov. and Moorea bouillonii comb. nov., tropical marine cyanobacteria rich in bioactive secondary metabolites. Int. J. Syst. Evol. Microbiol. 2012, 62, 1171–1178. [Google Scholar] [CrossRef]
- Engene, N.; Paul, V.J.; Byrum, T.; Gerwick, W.H.; Thor, A.; Ellisman, M.H. Five chemically rich species of tropical marine cyanobacteria of the genus Okeania gen. nov. (Oscillatoriales, Cyanoprokaryota). J. Phycol. 2013, 49, 1095–1106. [Google Scholar] [CrossRef]
- Taori, K.; Paul, V.J.; Luesch, H. Structure and activity of largazole, a potent anti-proliferative agent from the Floridian marine cyanobacterium Symploca sp. J. Am. Chem. Soc. 2008, 130, 1806–1807. [Google Scholar] [CrossRef]
- Salvador-Reyes, L.A.; Luesch, H. Biological targets and mechanisms of action of natural products from marine cyanobacteria. Nat. Prod. Rep. 2015, 32, 478–503. [Google Scholar] [CrossRef] [Green Version]
- Engene, N.; Tronholm, A.; Salvador-Reyes, L.A.; Luesch, H.; Paul, V.J. Caldora penicillata gen. nov., comb. nov. (Cyanobacteria), a pantropical marine species with biomedical relevance. J. Phycol. 2015, 51, 670–681. [Google Scholar] [CrossRef] [Green Version]
- Andrianasolo, E.H.; Gross, H.; Goeger, D.; Musafija-Girt, M.; McPhail, K.; Leal, R.M.; Mooberry, S.L.; Gerwick, W.H. Isolation of swinholide A and related glycosylated derivatives from two field collections of marine cyanobacteria. Org. Lett. 2005, 7, 1375–1378. [Google Scholar] [CrossRef]
- Salvador, L.A.; Paul, V.J.; Luesch, H. Caylobolide B, a macrolactone from symplostatin 1-producing marine cyanobacteria Phormidium spp. from Florida. J. Nat. Prod. 2010, 73, 1606–1609. [Google Scholar] [CrossRef] [Green Version]
- Plaza, A.; Bewley, C.A. Largamides A-H, unusual cyclic peptides from the marine cyanobacterium Oscillatoria sp. J. Org. Chem. 2006, 71, 6898–6907. [Google Scholar] [CrossRef] [PubMed]
- Thornburg, C.C.; Thimmaiah, M.; Shaala, L.A.; Hau, A.M.; Malmo, J.M.; Ishmael, J.E.; Youssef, D.T.A.; McPhail, K.L. Cyclic depsipeptides, grassypeptolides D and e and Ibu-epidemethoxylyngbyastatin 3, from a Red Sea Leptolyngbya cyanobacterium. J. Nat. Prod. 2011, 74, 1677–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, M.; Garcia, M.; Costa-Rodrigues, J.; Costa, M.S.; Ribeiro, M.J.; Fernandes, M.H.; Barros, P.; Barreiro, A.; Vasconcelos, V.; Martins, R. Exploring bioactive properties of marine cyanobacteria isolated from the Portuguese coast: High potential as a source of anticancer compounds. Mar. Drugs 2014, 12, 98–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regueiras, A.; Pereira, S.; Costa, M.S.; Vasconcelos, V. Differential toxicity of cyanobacteria isolated from marine sponges towards echinoderms and crustaceans. Toxins 2018, 10, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gkelis, S.; Panou, M. Capturing biodiversity: Linking a cyanobacteria culture collection to the “scratchpads” virtual research environment enhances biodiversity knowledge. Biodivers. Data J. 2016, 4, e7965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rippka, R. Isolation and Purification of Cyanobacteria. Methods Enzymol. 1988, 167, 3–27. [Google Scholar] [PubMed]
- Atashpaz, S.; Khani, S.; Barzegari, A.; Barar, J.; Vahed, S.Z.; Azarbaijani, R.; Omidi, Y. A robust universal method for extraction of genomic DNA from bacterial species. Microbiology 2010, 79, 538–542. [Google Scholar] [CrossRef]
- Nübel, U.; Garcia-Pichel, F.; Muyzer, G. PCR primers to amplify 16S rRNA genes from cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 3327–3332. [Google Scholar] [CrossRef] [Green Version]
- Taton, A.; Grubisic, S.; Brambilla, E.; De Wit, R.; Wilmotte, A. Cyanobacterial diversity in natural and artificial microbial mats of Lake Fryxell (McMurdo Dry Valleys, Antarctica): A morphological and molecular approach. Appl. Environ. Microbiol. 2003, 69, 5157–5169. [Google Scholar] [CrossRef] [Green Version]
- Moffitt, M.C.; Neilan, B.A. On the presence of peptide synthetase and polyketide synthase genes in the cyanobacterial genus Nodularia. FEMS Microbiol. Lett. 2001, 196, 207–214. [Google Scholar] [CrossRef]
- Neilan, B.A.; Dittmann, E.; Rouhiainen, L.; Bass, R.A.; Schaub, V.; Sivonen, K.; Börner, T. Nonribosomal peptide synthesis and toxigenicity of cyanobacteria. J. Bacteriol. 1999, 181, 4089–4097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashelford, K.E.; Chuzhanova, N.A.; Fry, J.C.; Jones, A.J.; Weightman, A.J. Media release • communique aux medias • medienmitteilung. World 2005, 71, 7724–7736. [Google Scholar]
- Komárek, J.; Kaštovský, J.; Mareš, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Christophoridis, C.; Zervou, S.-K.; Manolidi, K.; Katsiapi, M.; Moustaka-Gouni, M.; Kaloudis, T.; Triantis, T.M.; Hiskia, A. Occurrence and diversity of cyanotoxins in Greek lakes. Sci. Rep. 2018, 8, 17877. [Google Scholar] [CrossRef] [Green Version]
- Gkelis, S.; Panou, M.; Chronis, I.; Zervou, S.K.; Christophoridis, C.; Manolidi, K.; Ntislidou, C.; Triantis, T.M.; Kaloudis, T.; Hiskia, A.; et al. Monitoring a newly re-born patient: Water quality and cyanotoxin occurrence in a reconstructed shallow mediterranean lake. Adv. Oceanogr. Limnol. 2017, 8, 33–51. [Google Scholar] [CrossRef] [Green Version]
- Mavrogonatou, E.; Angelopoulou, M.T.; Kletsas, D. The catabolic effect of TNFa on bovine nucleus pulposus intervertebral disc cells and the restraining role of glucosamine sulfate in the TNFa-mediated up-regulation of MMP-3. J. Orthop. Res. 2014, 32, 1701–1707. [Google Scholar] [CrossRef]
- Dimozi, A.; Mavrogonatou, E.; Sklirou, A.; Kletsas, D. Oxidative stress inhibits the proliferation, induces premature senescence and promotes a catabolic phenotype in human nucleus pulposus intervertebral disc cells. Eur. Cells Mater. 2015, 30, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Mavrogonatou, E.; Konstantinou, A.; Kletsas, D. Long-term exposure to TNF-α leads human skin fibroblasts to a p38 MAPK- and ROS-mediated premature senescence. Biogerontology 2018, 19, 237–249. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cyanobacteria Strain (TAU-MAC) | Bacteria | |||
|---|---|---|---|---|
| Gram+ | Gram- | |||
| Staphylococcus aureus 9518 | Pseudomonas aeruginosa 12469 | Escherichia coli 8879 | Bacillus subtilis 3610 | |
| Xenococcus sp. 0615 | − | − | − | − |
| Synechococcus sp. 0715 | − | − | − | − |
| Synechococcus sp. 0815 | + | − | − | − |
| Leptothoe sithoniana 0915 | + | − | − | − |
| Leptothoe spongobia 1015 | − | − | − | − |
| Leptothoe spongobia 1115 | + | − | − | − |
| Leptothoe kymatousa 1215 | − | − | − | − |
| Schizotrichaceae sp. 1315 | − | − | − | − |
| Pseudanabaena cf. persicina 1415 | ++ | + | + | − |
| Pseudanabaena sp. 1515 | − | − | − | − |
| Leptothoe kymatousa 1615 | + | − | − | − |
| Leptolyngbya sp. 1715 | − | − | − | − |
| Leptolyngbya sp. 1815 | − | − | − | − |
| Xenococcus sp. 1915 | − | − | − | − |
| Synechococcus sp. 2015 | − | − | − | − |
| IC50 (μg·mL−1) | p Value (ANOVA test) | Fisher’s LSD | |
|---|---|---|---|
| TAU-MAC 0915 | |||
| Skin fibroblasts | 28.76 ± 18.79 | 0.0149 | ab |
| MDA-MB-231 | 44.58 ± 2.99 | b | |
| MCF-7 | 9.68 ± 0.58 | c | |
| A-431 | 29.12 ± 1.37 | ab | |
| HT-29 | 26.45 ± 8.39 | ac | |
| TAU-MAC 1015 | |||
| Skin fibroblasts | 45.74 ± 18.40 | 0.0439 | a |
| MDA-MB-231 | 26.17 ± 7.41 | b | |
| MCF-7 | 20.53 ± 9.68 | b | |
| A-431 | 14.03 ± 1.48 | b | |
| HT-29 | 17.56 ± 0.69 | b | |
| TAU-MAC 1115 | |||
| Skin fibroblasts | 45.25 ± 3.76 | 0.0000 | a |
| MDA-MB-231 | 37.26 ± 1.44 | b | |
| MCF-7 | 14.88 ±4.55 | c | |
| A-431 | 15.86 ± 0.73 | c | |
| HT-29 | 18.39 ± 1.37 | c | |
| TAU-MAC 1215 | |||
| Skin fibroblasts | 112.58 ± 7.86 | 0.0102 | a |
| MDA-MB-231 | 65.64 ± 2.43 | b | |
| MCF-7 | 34.95 ± 8.71 | b | |
| A-431 | 42.66 ± 31.55 | b | |
| HT-29 | 55.38 ± 17.51 | b |
| Strain (TAU-MAC) | Taxonomy | Sponge-Host | Habitat-Type | Reference |
|---|---|---|---|---|
| 0615 | Xenococcus sp. | Ircinia variabilis | overhang | [15] |
| 0715 | Synechococcus sp. | Axinella cannabina | overhang | [15] |
| 0815 | Synechococcus sp. | Axinella damicornis | overhang | [15] |
| 0915 | Leptothoe sithoniana | Petrosia (Petrosia) ficiformis | rocky reef | [16] |
| 1015 | Leptothoe spongobia | Dysidea avara | rocky reef | [16] |
| 1115 | Leptothoe spongobia | Acanthella acuta | overhang | [16] |
| 1215 | Leptothoe kymatousa | Chondrilla nucula | rocky reef | [16] |
| 1315 | Schizotricaceae sp. | Aplysina aerophoba | rocky reef | [15] |
| 1415 | Pseudanabaena cf. persicina | Axinella damicornis | overhang | [15] |
| 1515 | Pseudanabaena sp. | Spirastrella cunctatrix | rocky reef | This study |
| 1615 | Leptothoe kymatousa | Chondrilla nucula | rocky reef | [15] |
| 1715 | Leptolyngbya sp. | Hexadella racovitzai | overhang | This study |
| 1815 | Leptolyngbya sp. | Agelas oroides | rocky reef | This study |
| 1915 | Xenococcus sp. | Haliclona (Halichoclona) fulva | overhang | This study |
| 2015 | Synechococcus sp. | Stryphnus ponderosus | rocky reef | This study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konstantinou, D.; Mavrogonatou, E.; Zervou, S.-K.; Giannogonas, P.; Gkelis, S. Bioprospecting Sponge-Associated Marine Cyanobacteria to Produce Bioactive Compounds. Toxins 2020, 12, 73. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020073
Konstantinou D, Mavrogonatou E, Zervou S-K, Giannogonas P, Gkelis S. Bioprospecting Sponge-Associated Marine Cyanobacteria to Produce Bioactive Compounds. Toxins. 2020; 12(2):73. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020073
Chicago/Turabian StyleKonstantinou, Despoina, Eleni Mavrogonatou, Sevasti-Kiriaki Zervou, Panagiotis Giannogonas, and Spyros Gkelis. 2020. "Bioprospecting Sponge-Associated Marine Cyanobacteria to Produce Bioactive Compounds" Toxins 12, no. 2: 73. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020073