The Effectiveness of Durian Peel as a Multi-Mycotoxin Adsorbent

 ,
,  , , ,
, , ,
Abstract
:
1. Introduction
2. Results and Discussion
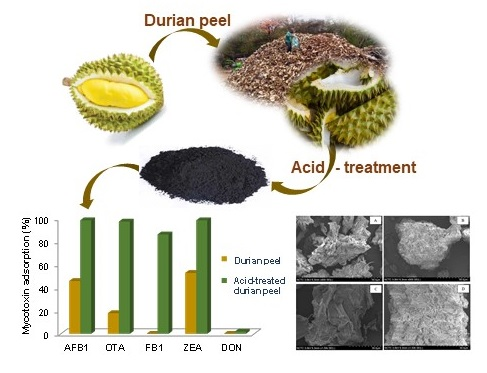
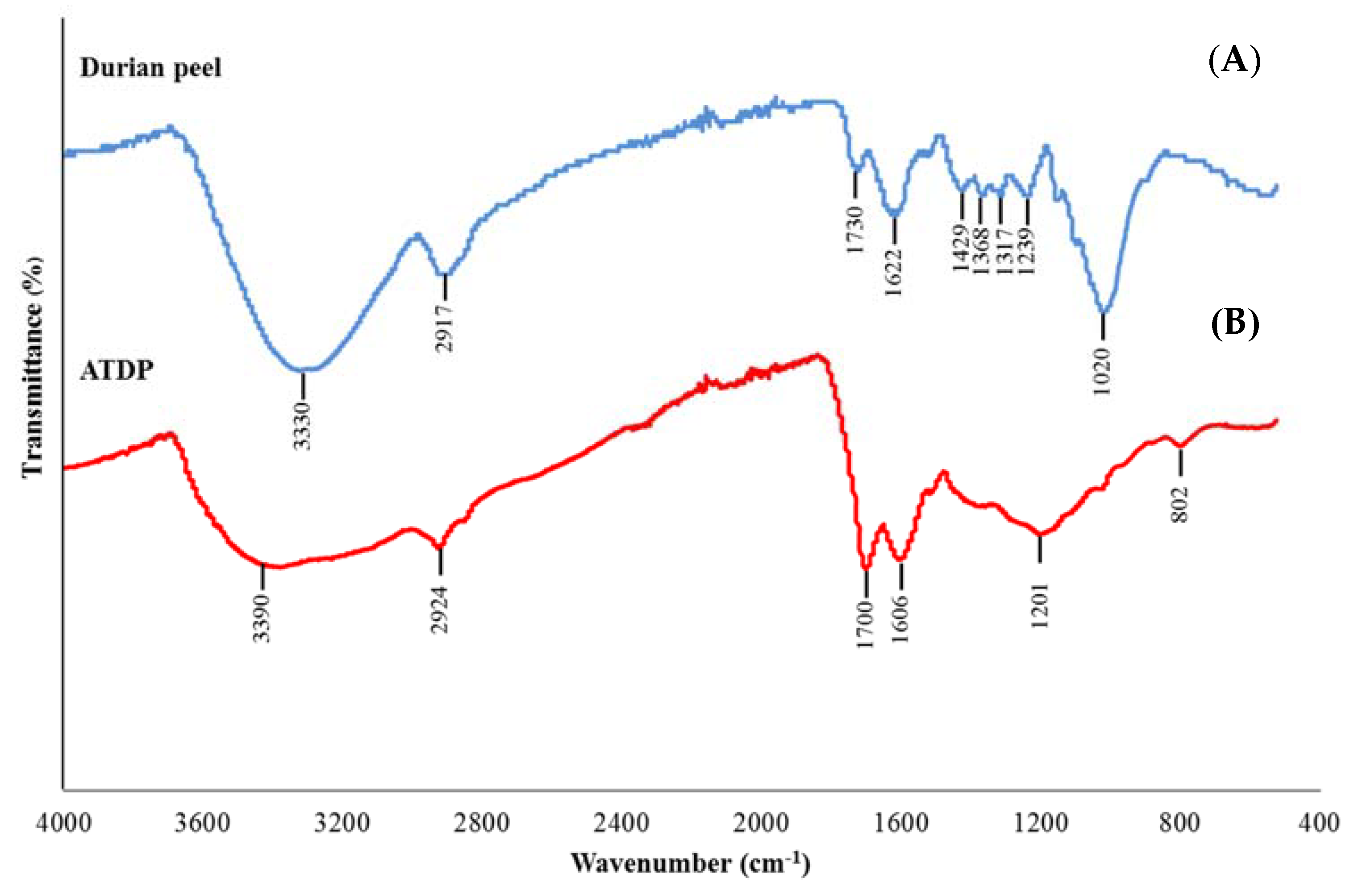
2.1. Characterization of Durian Peel
2.2. Screening of DP and ATDP as Multi-Mycotoxin Adsorbing Agents
2.3. Effect of Medium pH on Mycotoxin Adsorption and Desorption
2.4. Effect of ATDP Dosage
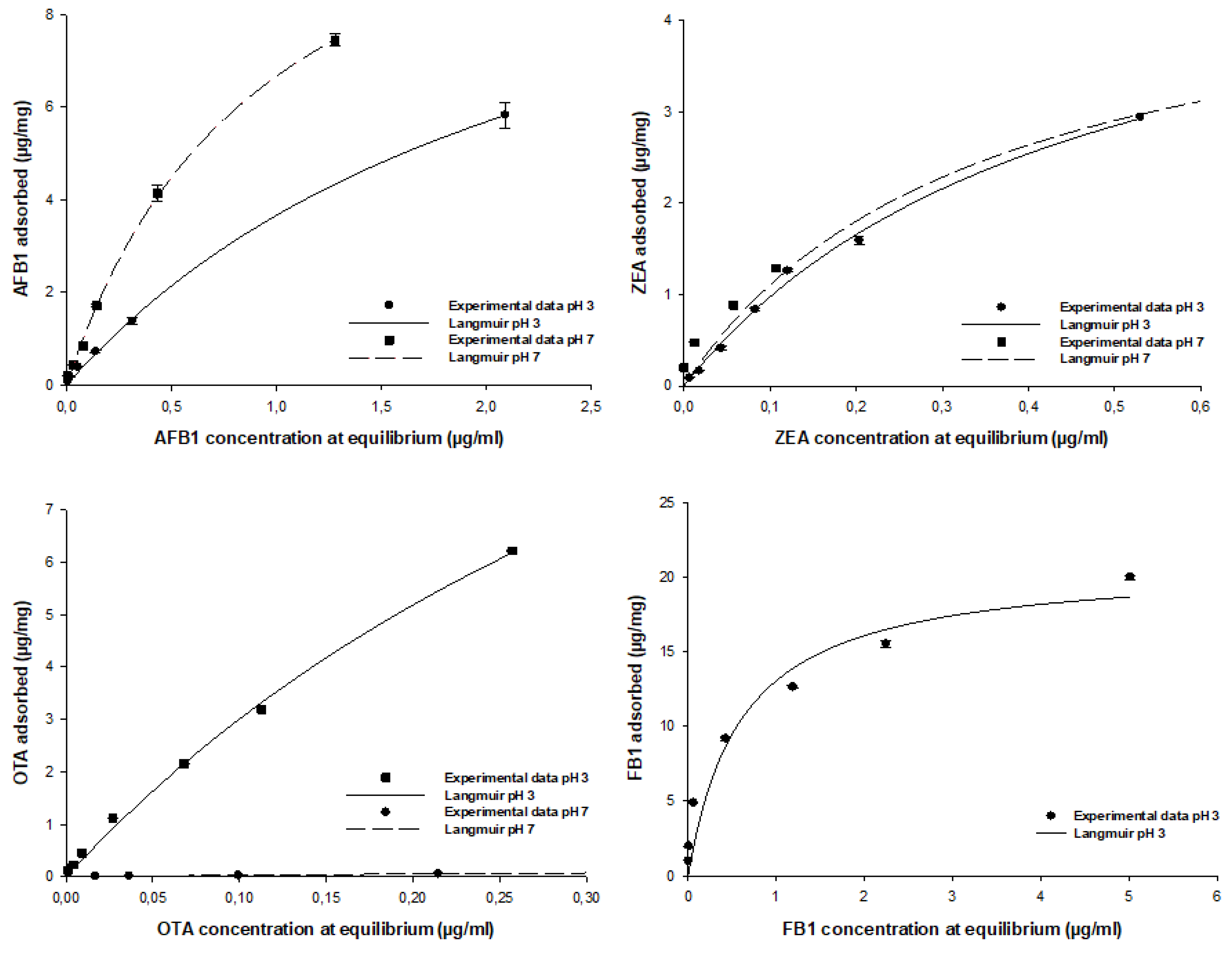
2.5. Equilibrium Adsorption Isotherm
2.6. Multi-Mycotoxin Adsorption in Simulated Gastrointestinal Fluid
3. Conclusions
4. Materials and Methods
4.1. Reagents and Samples
4.2. Physico-Chemical Characterization of DP and ATDP
4.3. Multi-Mycotoxin Adsorption Experiments
4.4. Equilibrium Adsorption Isotherms
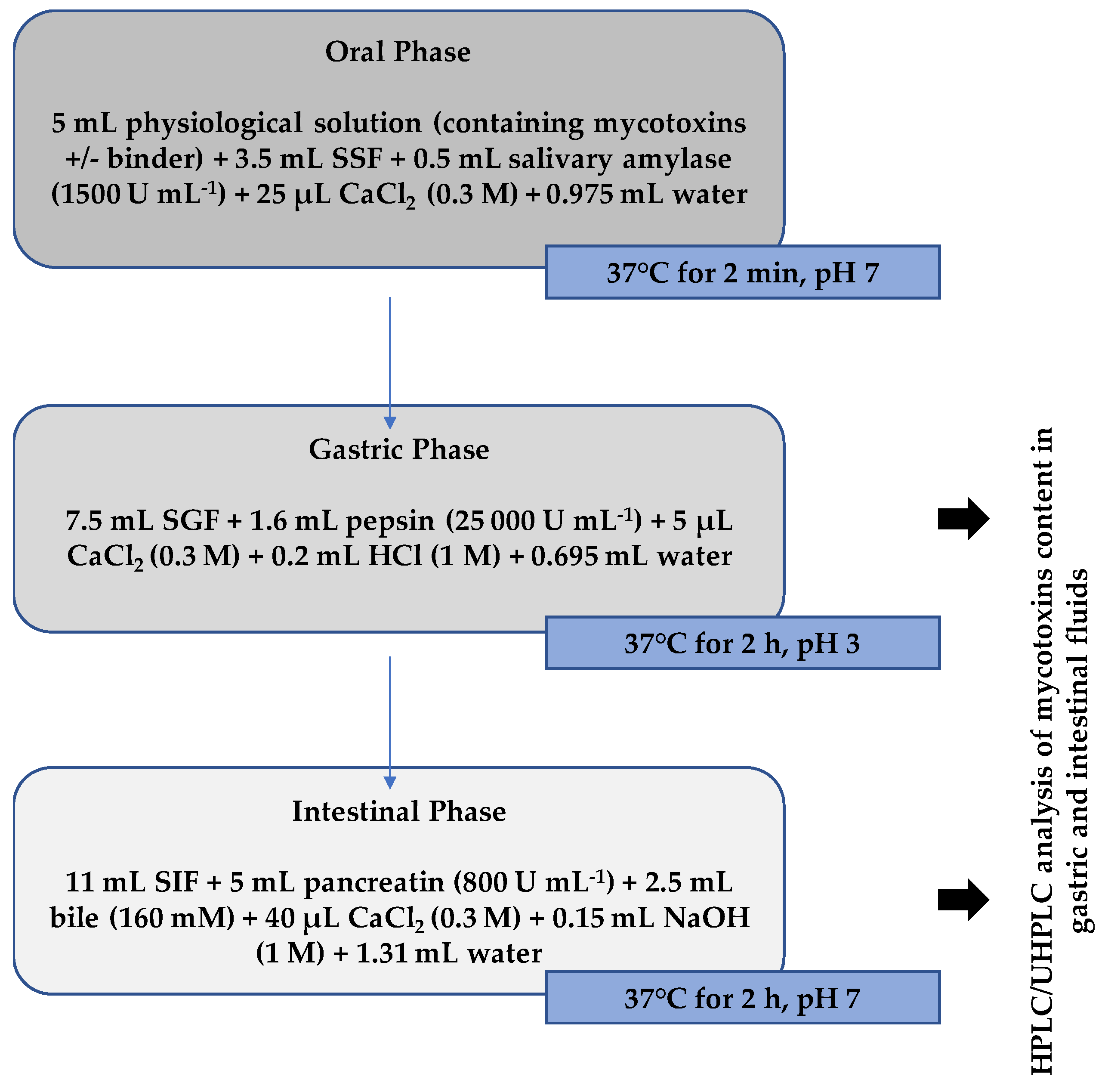
4.5. Simulated Gastrointestinal Digestion
4.6. Data Calculation and Curve Fitting
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Anfossi, L.; Giovannoli, C.; Baggiani, C. Mycotoxin detection. Curr. Opin. Biotechnol. 2016, 37, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Patriarca, A.; Pinto, V.F. Prevalence of mycotoxins in foods and decontamination. Curr. Opin. Food Sci. 2017, 14, 50–60. [Google Scholar] [CrossRef]
- Freire, L.; Sant’Ana, A.S. Modified mycotoxins: An updated review on their formation, detection, occurrence, and toxic effects. Food Chem. Toxicol. 2018, 111, 189–205. [Google Scholar] [CrossRef] [PubMed]
- Zaki, M.M.; El-Midany, S.A.; Shasheen, H.M.; Rizzi, L. Mycotoxins in animals: Occurrence, effects, prevention and management. J. Toxicol. Environ. Health Sci. 2012, 4, 13–28. [Google Scholar] [CrossRef]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [Green Version]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Wei, D.; Wang, Y.; Jiang, D.; Feng, X.; Li, J.; Wang, M. Survey of alternaria toxins and other mycotoxins in dried fruits in China. Toxins 2017, 9, 200. [Google Scholar] [CrossRef] [Green Version]
- Grenier, B.; Oswald, I. Mycotoxin co-contamination of food and feed: Meta-analysis of publications describing toxicological interactions. World Mycotoxin J. 2011, 4, 285–313. [Google Scholar] [CrossRef]
- Paterson, R.R. Toxicology of mycotoxins. Exp. Suppl. 2010, 100, 31–63. [Google Scholar]
- Speijers, G.J.; Speijers, M.H. Combined toxic effects of mycotoxins. Toxicol. Lett. 2004, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Huwig, A.; Freimund, S.; Käppeli, O.; Dutler, H. Mycotoxin detoxication of animal feed by different adsorbents. Toxicol. Lett. 2001, 122, 179–188. [Google Scholar] [CrossRef]
- Kolosova, A.; Stroka, J. Substances for reduction of the contamination of feed by mycotoxins: A review. World Mycotoxin J. 2011, 4, 225–256. [Google Scholar] [CrossRef]
- Pappas, A.C.; Tsiplakou, E.; Georgiadou, M.; Anagnostopoulos, C.; Markoglou, A.N.; Liapis, K.; Zervas, G. Bentonite binders in the presence of mycotoxins: Results of in vitro preliminary tests and an in vivo broiler trial. Appl. Clay Sci. 2014, 99, 48–53. [Google Scholar] [CrossRef]
- Avantaggiato, G.; Solfrizzo, M.; Visconti, A. Recent advances on the use of adsorbent materials for detoxification of fusarium mycotoxins. Food Addit. Contam. 2005, 22, 379–388. [Google Scholar] [CrossRef]
- Greco, D.; D’Ascanio, V.; Santovito, E.; Logrieco, A.F.; Avantaggiato, G. Comparative efficacy of agricultural by-products in sequestering mycotoxins. J. Sci. Food Agric. 2018, 99, 1623–1634. [Google Scholar] [CrossRef]
- Li, Y.; Tian, G.; Dong, G.; Bai, S.; Han, X.; Liang, J.; Meng, J.; Zhang, H. Research progress on the raw and modified montmorillonites as adsorbents for mycotoxins: A review. Appl. Clay Sci. 2018, 163, 299–311. [Google Scholar] [CrossRef]
- Dai, Y.; Sun, Q.; Wang, W.; Lu, L.; Liu, M.; Li, J.; Yang, S.; Sun, Y.; Zhang, K.; Xu, J.; et al. Utilizations of agricultural waste as adsorbent for the removal of contaminants: A review. Chemosphere 2018, 211, 235–253. [Google Scholar] [CrossRef]
- Lazim, Z.M.; Hadibarata, T.; Puteh, M.H.; Yusop, Z. Adsorption characteristics of bisphenol A onto low-cost modified phyto-waste material in aqueous solution. Water Air Soil Pollut. 2015, 226, 34. [Google Scholar] [CrossRef]
- Nuithitikul, K.; Srikhun, S.; Hirunpraditkoon, S. Influences of pyrolysis condition and acid treatment on properties of durian peel-based activated carbon. Bioresour. Technol. 2010, 101, 426–429. [Google Scholar] [CrossRef]
- Charoenvai, S. Durian peels fiber and recycled HDPE composites obtained by extrusion. Energy Procedia 2014, 56, 539–546. [Google Scholar] [CrossRef] [Green Version]
- Lazim, Z.M.; Hadibarata, T.; Puteh, M.H.; Yusop, Z.; Wirasnita, R.; Nor, N.M. Utilization of durian peel as potential adsorbent for biphenol A removal in aquoeus solution. J. Teknol. 2015, 74, 109–115. [Google Scholar]
- Mitan, N.M.M.; Ramlan, M.S.; Nawawi, M.Z.H.; Gazali, M.H.M. Performance of binders in briquetting of durian peel as a solid biofuel. Mater. Today Proc. 2018, 5, 21753–21758. [Google Scholar] [CrossRef]
- Feng, N.; Guo, X.; Liang, S.; Zhu, Y.; Liu, J. Biosorption of heavy metals from aqueous solutions by chemically modified orange peel. J. Hazard. Mater. 2011, 185, 49–54. [Google Scholar] [CrossRef]
- Ngabura, M.; Hussain, S.A.; Ghani, W.A.W.A.; Jami, M.S.; Tan, Y.P. Utilization of renewable durian peels for biosorption of zinc from wastewater. J. Environ. Chem. Eng. 2018, 6, 2528–2539. [Google Scholar] [CrossRef]
- Avantaggiato, G.; Greco, D.; Damascelli, A.; Solfrizzo, M.; Visconti, A. Assessment of multi-mycotoxin adsorption efficacy of grape pomace. J. Agric. Food Chem. 2014, 62, 497–507. [Google Scholar] [CrossRef]
- Foo, K.Y.; Hameed, B.H. Insights into the modeling of adsorption isotherm systems. Chem. Eng. J. 2010, 156, 2–10. [Google Scholar] [CrossRef]
- Lemke, S.L.; Grant, P.G.; Phillips, T.D. Adsorption of zearalenone by organophilic montmorillonite clay. J. Agric. Food Chem. 1998, 46, 3789–3796. [Google Scholar] [CrossRef]
- Niderkorn, V.; Morgavi, D.P.; Aboab, B.; Lemaire, M.; Boudra, V. Cell wall component and mycotoxin moieties involved in the binding of fumonisin B1 and B2 by lactic acid bacteria. J. Appl. Microbiol. 2009, 106, 977–985. [Google Scholar] [CrossRef]
- Sangsila, A.; Faucet-Marquis, V.; Pfohl-Leszkowicz, A.; Itsaranuwat, P. Detoxification of zearalenone by Lactobacillus pentosus strains. Food Control 2016, 62, 187–192. [Google Scholar] [CrossRef]
- Armando, M.R.; Pizzolitto, R.P.; Dogi, C.A.; Cristofolini, A.; Merkis, C.; Poloni, V.; Dalcero, A.M.; Cavaglieri, L.R. Adsorption of ochratoxin A and zearalenone by potential probiotic Saccharomyces cerevisiae strains and its relation with cell wall thickness. J. Appl. Microbiol. 2012, 113, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Bejaoui, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Biodegradation of ochratoxin A by Aspergillus section Nigri species isolated from French grapes: A potential means of ochratoxin A decontamination in grape juices and musts. FEMS Microbiol. Lett. 2006, 255, 203–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solís-Cruz, B.; Hernández-Patlán, D.; Beyssac, E.; Latorre, J.D.; Hernandez-Velasco, X.; Merino-Guzman, R.; Tellez, G.; López-Arellano, R. Evaluation of chitosan and cellulosic polymers as binding adsorbent materials to prevent aflatoxin B1, fumonisin B1, ochratoxin, trichothecene, deoxynivalenol, and zearalenone mycotoxicoses through an in vitro gastrointestinal model for poultry. Polymers 2017, 9, 529. [Google Scholar] [CrossRef] [PubMed]
- Vila-Donat, P.; Marín, S.; Sanchis, V.; Ramos, A.J. A review of the mycotoxin adsorbing agents, with an emphasis on their multi-binding capacity, for animal feed decontamination. Food Chem. Toxicol. 2018, 114, 246–259. [Google Scholar] [CrossRef] [Green Version]
- Versantvoort, C.H.; Oomen, A.G.; Van de Kamp, E.; Rompelberg, C.J.; Sips, A.J. Applicability of an in vitro digestion model in assessing the bioaccessibility of mycotoxins from food. Food Chem. Toxicol. 2005, 43, 31–40. [Google Scholar] [CrossRef]
- Avantaggiato, G.; Havenaar, R.; Visconti, A. Assessing the zearalenone-binding activity of adsorbent materials during passage through a dynamic in vitro gastrointestinal model. Food Chem. Toxicol. 2003, 41, 1283–1290. [Google Scholar] [CrossRef]
- Avantaggiato, G.; Havenaar, R.; Visconti, A. Evaluation of the intestinal absorption of deoxynivalenol and nivalenol by an in vitro gastrointestinal model, and the binding efficacy of activated carbon and other adsorbent materials. Food Chem. Toxicol. 2004, 42, 817–824. [Google Scholar] [CrossRef]
- Hur, S.J.; Beong, O.L.; Decker, E.A.; Mcclements, D.J. In vitro human digestion models for food applications. Food Chem. 2011, 125, 1–12. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Balance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Cochrane, E.L.; Lu, S.; Gibb, S.W.; Villaescusa, I. A comparison of low-cost biosorbents and commercial sorbents for the removal of copper from aqueous media. J. Hazard. Mater. 2006, 137, 198–206. [Google Scholar] [CrossRef]
- Ismail, A.; Sudrajat, H.; Jumbianti, D. Activated carbon from durian seed by H3PO4 activation: Preparation and pore structure characterization. Indones. J. Chem. 2010, 10, 36–40. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adsorbent | Nanoparticle (nm) | Pore Volume (cm3/g) | Pore Diameter (nm) | Surface Area (m2/g) |
|---|---|---|---|---|
| DP | 3032.45 | 0.004 | 7.22 | 1.98 |
| ATDP | 142.95 | 0.162 | 15.46 | 41.97 |
| Toxin | DP | ATDP | ||
|---|---|---|---|---|
| pH 3 | pH 7 | pH 3 | pH 7 | |
| AFB1 | 46 ± 4 | 37 ± 2 | 98.4 ± 0.1 | 98.4 ± 0.1 |
| ZEA | 53 ± 2 | 52 ± 4 | 98.4 ± 0.4 | 99.6 ± 0.2 |
| OTA | 18 ± 1 | 0.7 ± 0.6 | 97.3 ± 0.1 | 42.2 ± 0.2 |
| FB1 | 0 | 2.3 ± 0.7 | 86 ± 3 | 0 |
| DON | 0 | 2 ± 1 | 2.0 ± 0.8 | 13 ± 2 |
| Toxin | Adsorption (%) | Desorption (%) | |
|---|---|---|---|
| pH 3 | pH 7 | Methanol | |
| AFB1 | 100 | 0 | 34 ± 3 |
| ZEA | 98.9 ± 0.4 | 0.8 ± 0.2 | 85 ± 4 |
| OTA | 99.0 ± 0.3 | 2.0 ± 0.5 | 48 ± 3 |
| FB1 | 91 ± 3 | 1.6 ± 0.3 | 6.5 ± 0.5 |
| Toxin | Adsmax (%) | C50 (mg/mL) |
|---|---|---|
| AFB1 | 101 ± 1 | 0.11 |
| ZEA | 104 ± 1 | 0.19 |
| OTA | 103 ± 1 | 5.77 |
| Parameters | AFB1 | ZEA | OTA | FB1 | ||||
|---|---|---|---|---|---|---|---|---|
| pH 3 | pH 7 | pH 3 | pH 7 | pH 3 | pH 7 | pH 3 | ||
| Freundlich | Kf (±SE) | 3.32 ± 0.05 | 6.43 ± 0.12 | 4.66 ± 0.17 | 3.33 ± 0.04 | 18.09 ± 0.17 | 0.17 ± 0.00 | 11.86 ± 0.14 |
| 1/n (±SE) | 0.76 ± 0.02 | 0.67 ± 0.03 | 0.69 ± 0.02 | 0.45 ± 0.01 | 0.79 ± 0.01 | 0.77 ± 0.02 | 0.33 ± 0.00 | |
| R2 | 0.9977 | 0.9892 | 0.9850 | 0.9948 | 0.9997 | 0.9966 | 0.9976 | |
| SSres | 0.1705 | 1.2987 | 0.0120 | 0.1517 | 0.0272 | 0.0001 | 4.3605 | |
| Sylx | 0.1032 | 0.2849 | 0.1097 | 0.1041 | 0.0379 | 0.0022 | 0.4922 | |
| PRESS | 0.3656 | 1.6869 | 0.2661 | 0.1668 | 0.0336 | 0.0001 | 4.6588 | |
| Normality | failed | failed | failed | passed | passed | passed | failed | |
| Constant Variance Test | passed | passed | passed | passed | passed | passed | failed | |
| Langmuir | Adsmax (±SE) | 12.69 ± 0.93 | 13.02 ± 0.40 | 5.47 ± 0.28 | 4.90 ± 0.34 | 18.82 ± 1.49 | 0.26 ± 0.02 | 20.89 ± 1.20 |
| KL (±SE) | 0.41 ± 0.05 | 1.05 ± 0.06 | 2.17 ± 0.18 | 2.91 ± 0.61 | 1.90 ± 0.21 | 1.30 ± 0.12 | 1.66 ± 0.35 | |
| R2 | 0.9966 | 0.9980 | 0.9938 | 0.9566 | 0.9970 | 0.9974 | 0.9754 | |
| SSres | 0.2456 | 0.2394 | 0.0896 | 1.2691 | 0.2609 | 0.0001 | 43.4700 | |
| Sylx | 0.1239 | 0.1223 | 0.0706 | 0.3011 | 0.1170 | 0.0019 | 1.5540 | |
| PRESS | 0.4548 | 0.3329 | 0.111 | 1.9306 | 0.2996 | 0.0001 | 50.6400 | |
| Normality | passed | passed | passed | passed | passed | passed | passed | |
| Constant Variance Test | passed | passed | passed | passed | passed | passed | passed | |
| Sips | qm (±SE) | - | 11.67 ± 0.75 | 5.28 ± 0.73 | - | - | - | 91.70 ± 52.45 |
| As (±SE) | - | 1.35 ± 0.90 | 2.38 ± 0.81 | - | - | - | 0.15 ± 0.10 | |
| 1/n = nH (±SE) | - | 1.08 ± 0.97 | 1.02 ± 0.08 | - | - | - | 0.37 ± 0.03 | |
| R2 | - | 0.9983 | 0.9938 | - | - | - | 0.9979 | |
| SSres | - | 0.206 | 0.0892 | - | - | - | 3.8247 | |
| Sylx | - | 0.1172 | 0.0724 | - | - | - | 0.4743 | |
| PRESS | - | 0.3403 | 0.1242 | - | - | - | 4.2181 | |
| Normality | failed | passed | - | - | - | failed | ||
| Constant Variance Test | passed | failed | - | - | - | failed | ||
| Toxin | Bioaccessibility (%) | Bioaccessibility Reduction (%) | ||||
|---|---|---|---|---|---|---|
| Gastric Phase | Intestinal Phase | Gastric Phase | Intestinal Phase | |||
| Control | +ATDP | Control | +ATDP | |||
| AFB1 | 96.5 ± 0.9 | 6.3 ± 0.1 | 96.3 ± 0.8 | 4.3 ± 0.2 | 93.6 ± 0.1 | 95.1 ± 0.0 |
| ZEA | 33.5 ± 0.9 | 0.8 ± 0.1 | 39.7 ± 1.4 | 12.4 ± 0.4 | 97.6 ± 0.1 | 68.9 ± 1.1 |
| OTA | 49.6 ± 0.7 | 1.9 ± 0.5 | 43.6 ± 1.3 | 39.2 ± 1.1 | 96.2 ± 1.0 | 10.1 ± 2.6 |
| FB1 | 78.8 ± 1.9 | 25.8 ± 2.9 | 81.2 ± 2.9 | 78.9 ± 1.9 | 67.3 ± 3.7 | 2.6 ± 2.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adunphatcharaphon, S.; Petchkongkaew, A.; Greco, D.; D’Ascanio, V.; Visessanguan, W.; Avantaggiato, G. The Effectiveness of Durian Peel as a Multi-Mycotoxin Adsorbent. Toxins 2020, 12, 108. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020108
Adunphatcharaphon S, Petchkongkaew A, Greco D, D’Ascanio V, Visessanguan W, Avantaggiato G. The Effectiveness of Durian Peel as a Multi-Mycotoxin Adsorbent. Toxins. 2020; 12(2):108. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020108
Chicago/Turabian StyleAdunphatcharaphon, Saowalak, Awanwee Petchkongkaew, Donato Greco, Vito D’Ascanio, Wonnop Visessanguan, and Giuseppina Avantaggiato. 2020. "The Effectiveness of Durian Peel as a Multi-Mycotoxin Adsorbent" Toxins 12, no. 2: 108. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12020108