Physiological and Metabolic Responses of Marine Mussels Exposed to Toxic Cyanobacteria Microcystis aeruginosa and Chrysosporum ovalisporum

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
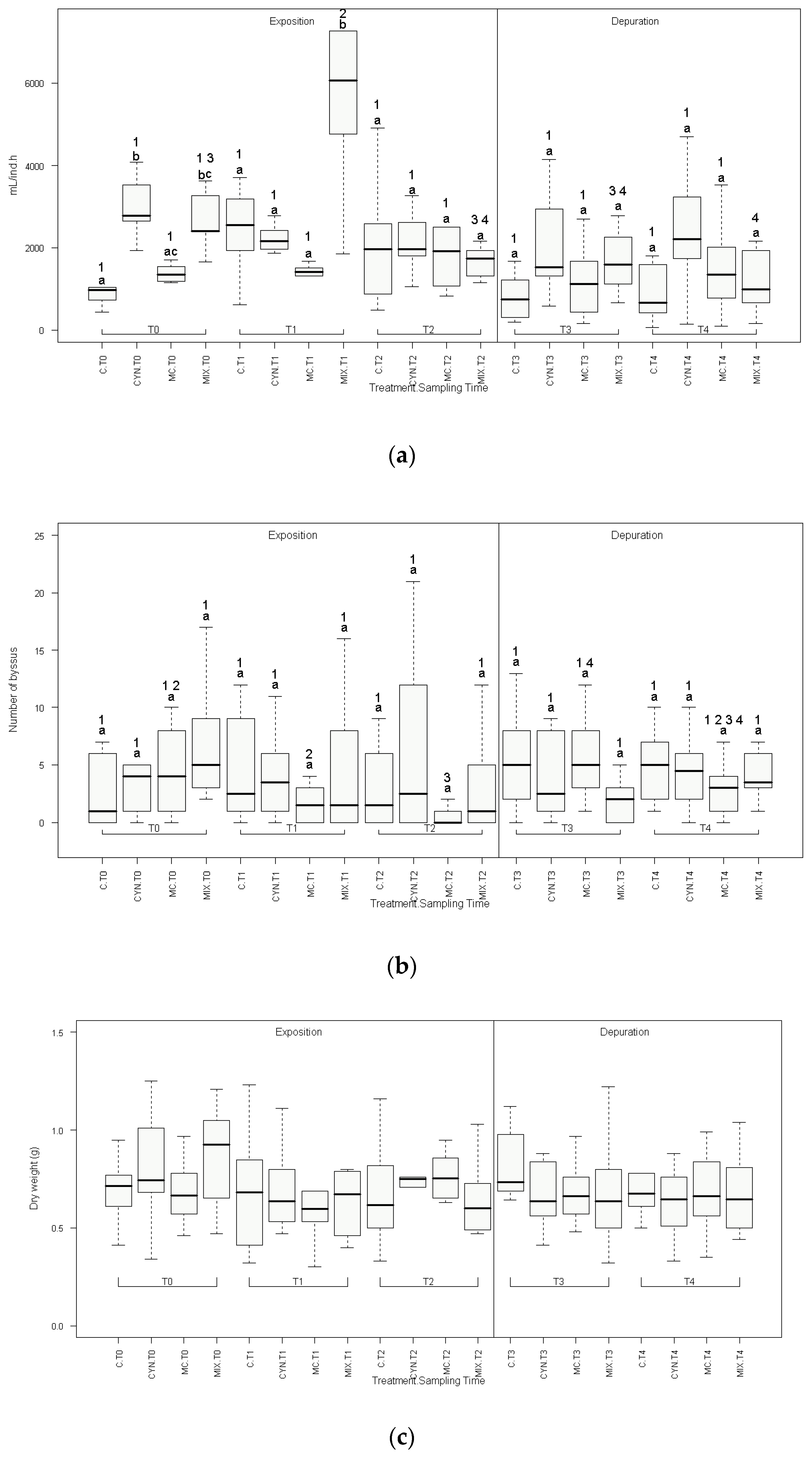
2.1. Physiological Parameters
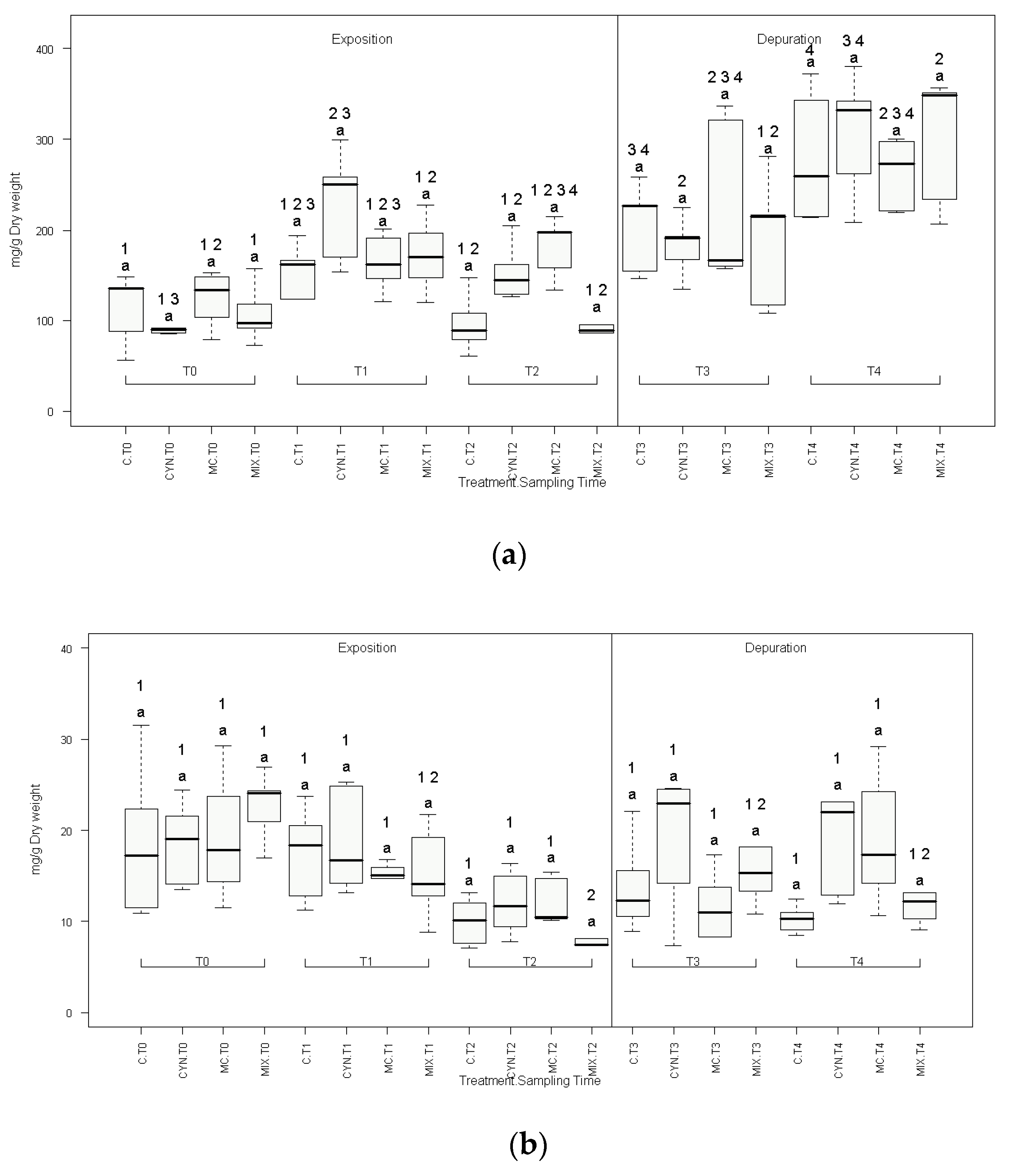
2.2. Biochemical Parameters
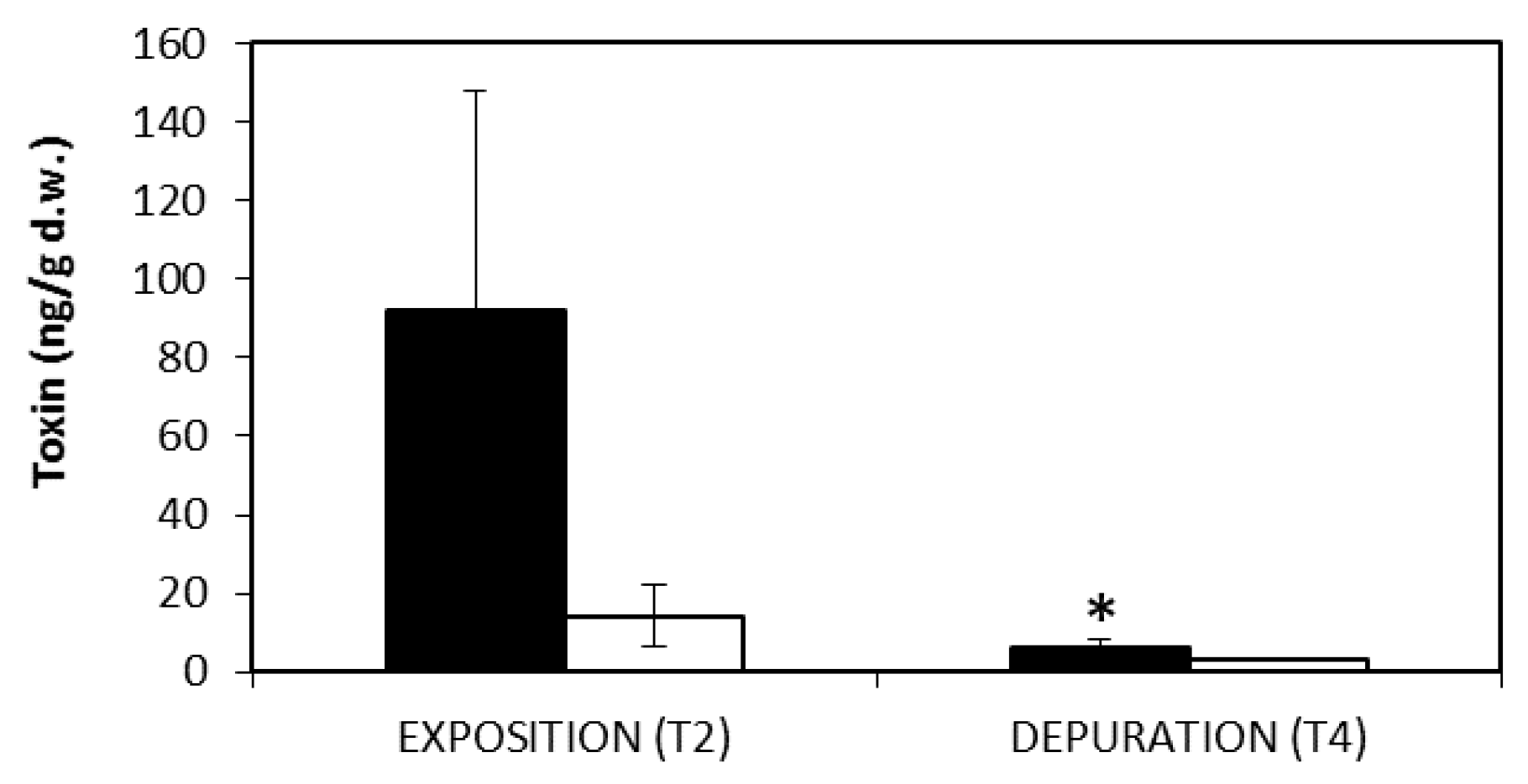
2.3. Toxin Accumulation in Mussels
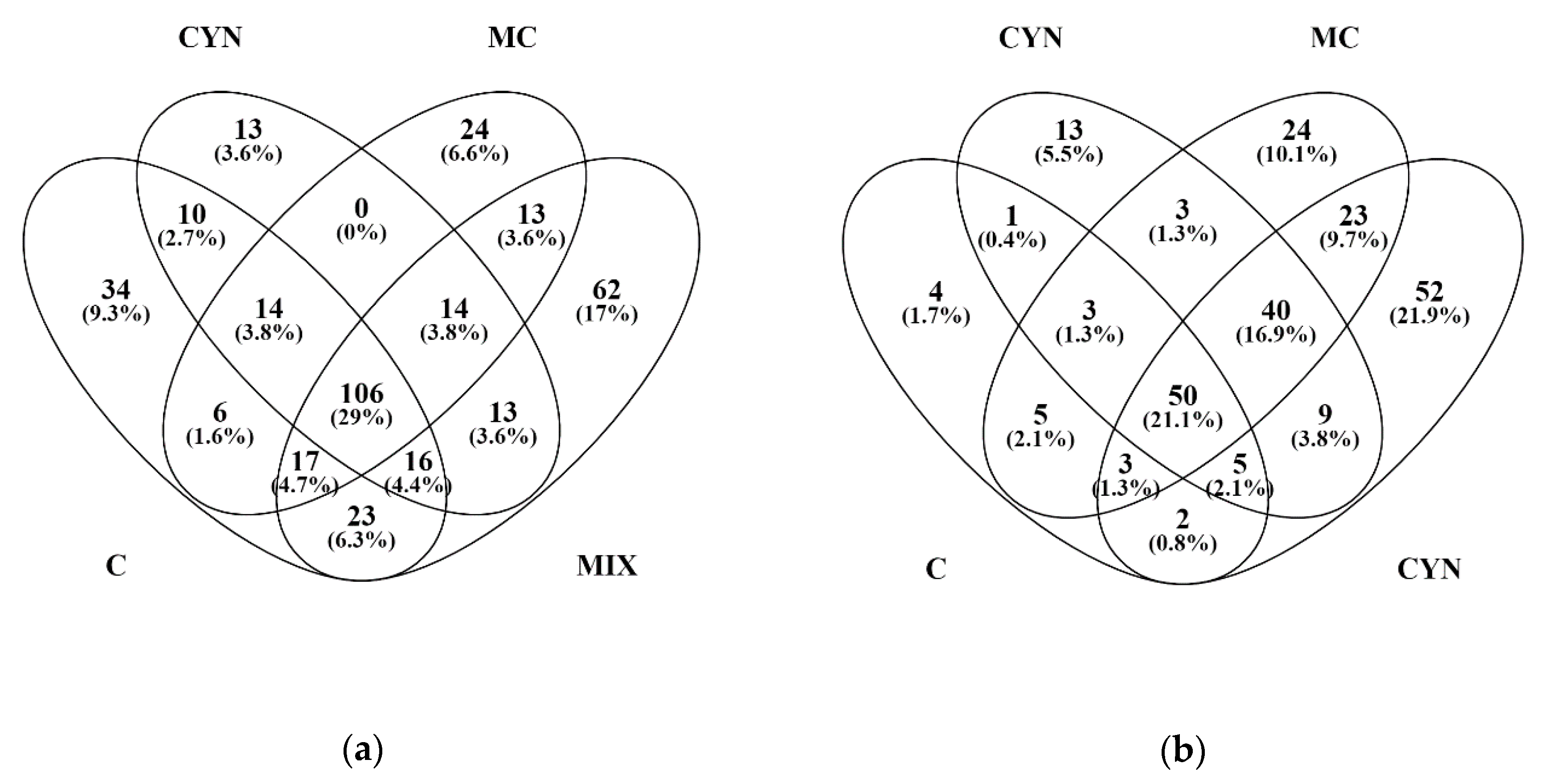
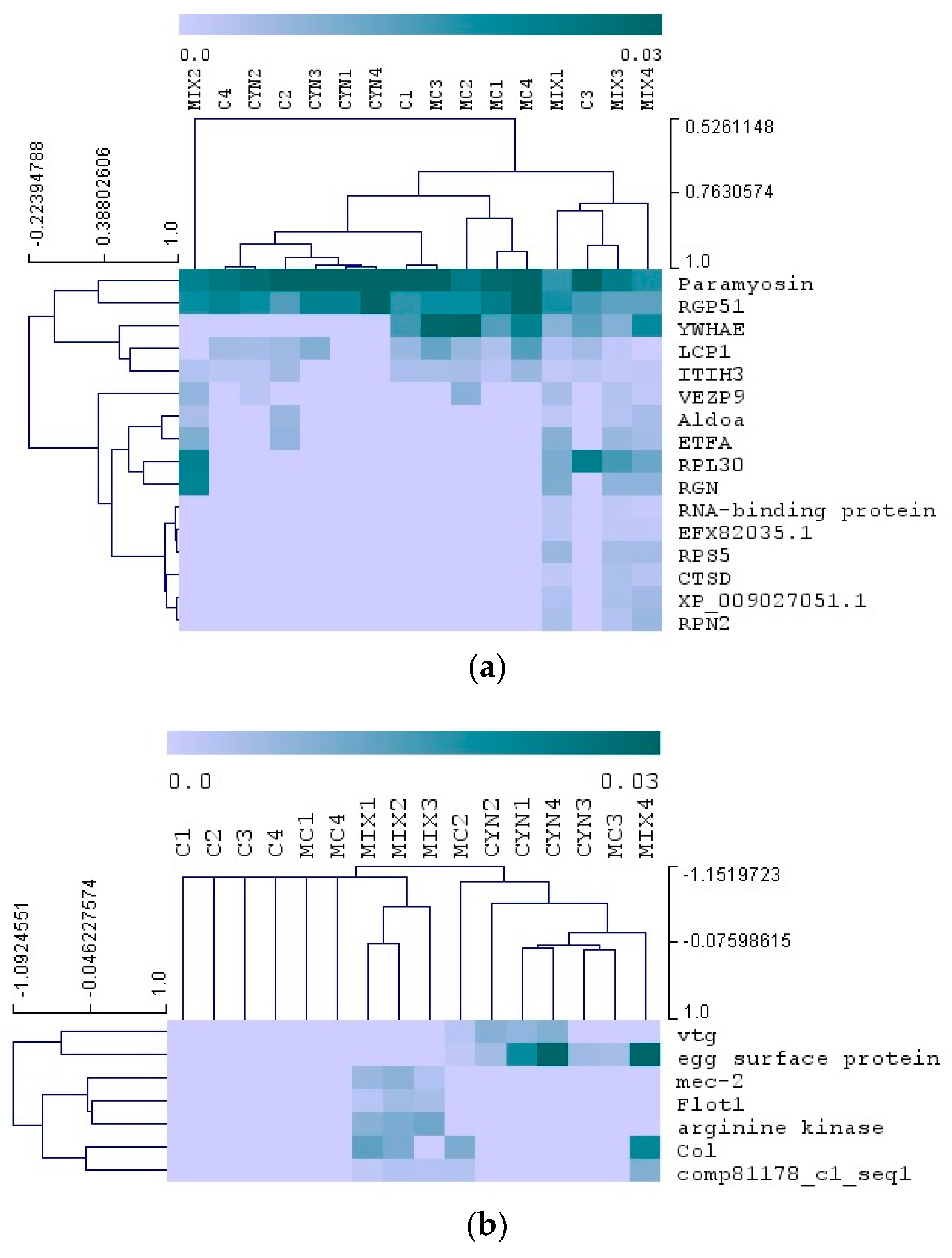
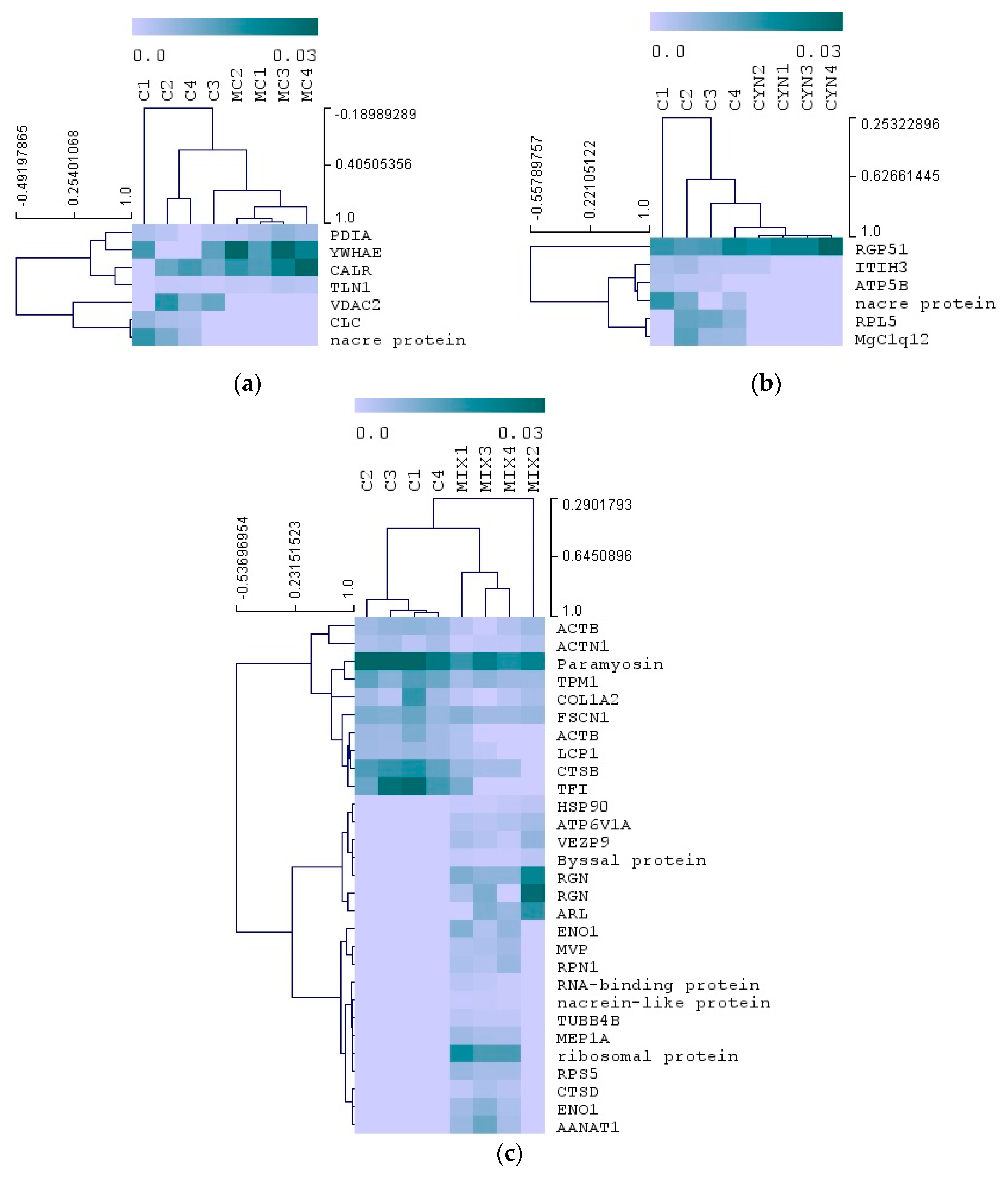
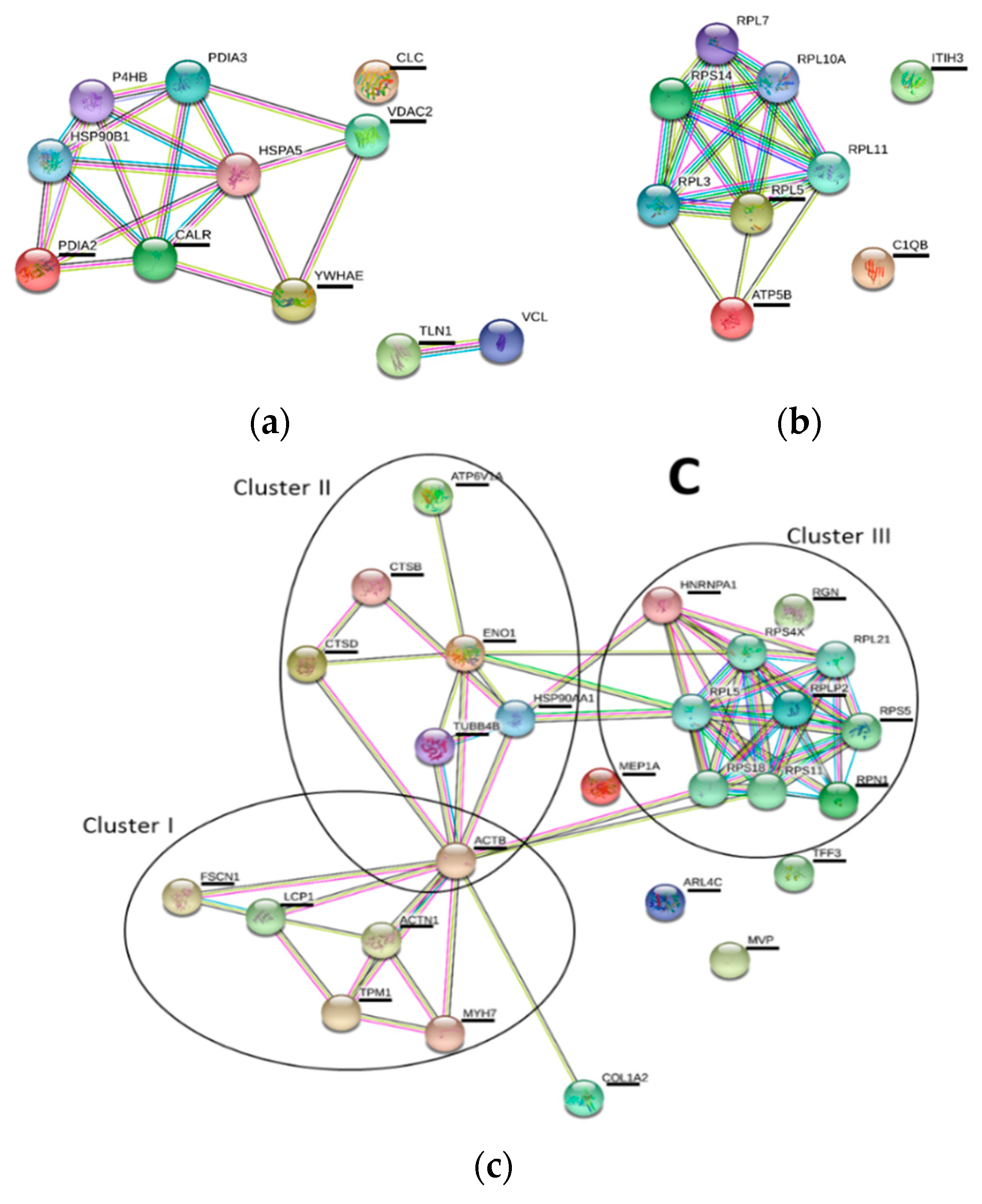
2.4. Proteomics
3. Discussion
3.1. Implication of Cyanobacteria Feeding in Mussels Health
3.2. Mussel Contamination
3.3. Metabolic Responses of Mussels to Toxic Cyanobacteria
4. Conclusions
5. Materials and Methods
5.1. Microalgae Culture
5.2. Toxin Extraction and Quantification in Microalgae
5.3. Mussel Harvest and Maintenance
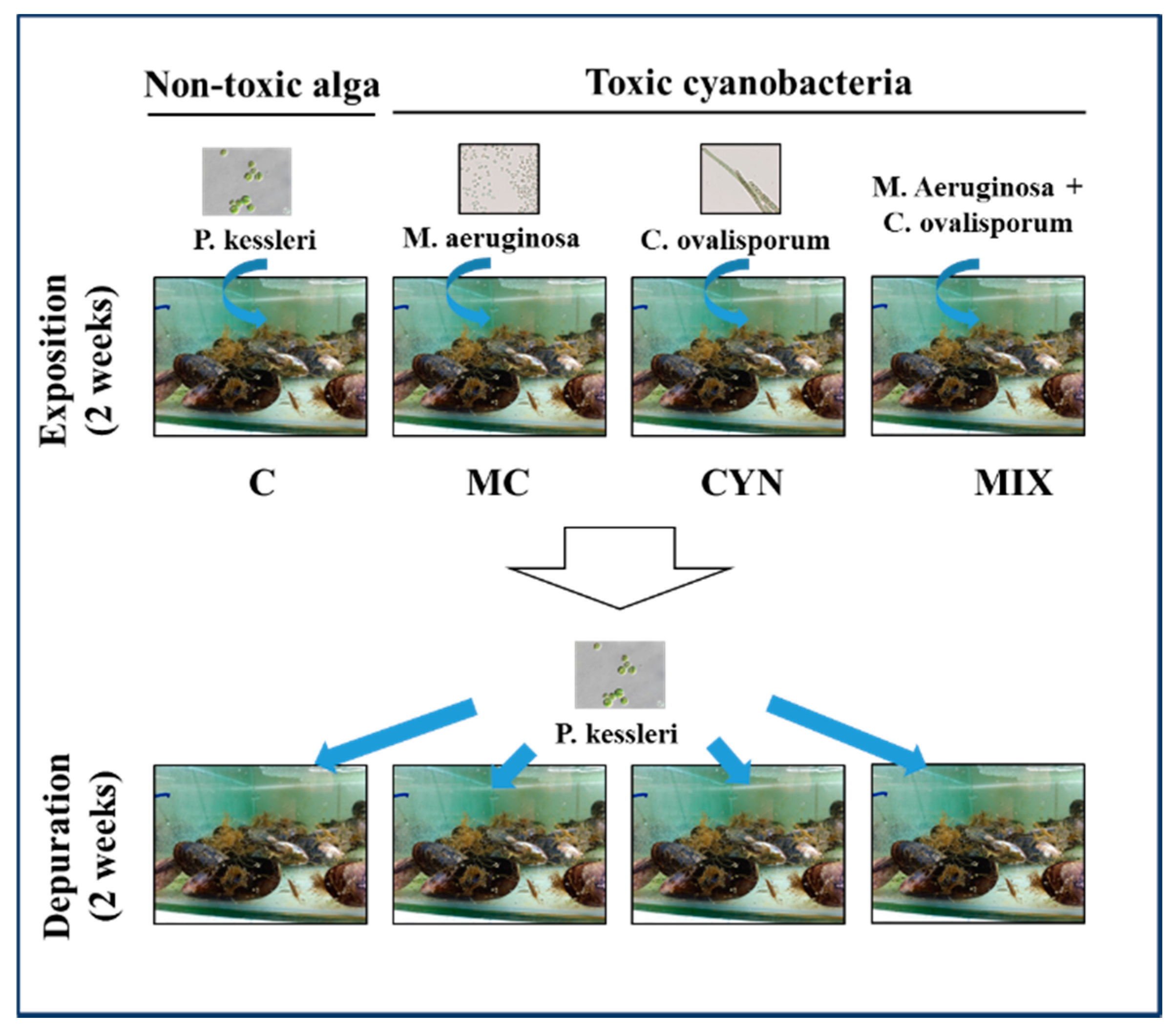
5.4. Experimental Design
5.5. Physiological Parameters
5.5.1. Filtration Rates
5.5.2. Byssus Count
5.5.3. Fresh and Dry Weight
5.6. Biochemical Parameters
5.7. Toxin Analysis in Mussels
5.8. Proteomics
5.8.1. Sample Preparation
5.8.2. LC-MS/MS
5.8.3. Protein Identification and Quantification
5.9. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Azevedo, S.M.F.O.; Brandão, C.C.S. Cianobactérias Tóxicas na água Para Consumo Humano na saúde Pública e Processos de Remoção em água Para Consumo Humano; Ministério da Saúde: Fundação Nacional de Saúde: Brasília, Brasília, 2003. [Google Scholar]
- Fernandes, S.S. Biodisponibilidade de Cianotoxinas em Bivalves; Faculdade de Ciêncas da Universidade do Porto: Porto, Portugal, 2008. [Google Scholar]
- Vasconcelos, V.M. Uptake and depuration of the heptapeptide toxin microcystin-LR in Mytilus galloprovincialis. Aquat. Toxicol. 1995, 32, 227–237. [Google Scholar] [CrossRef]
- Dawson, R.M. Review Article the Toxicology of Microcystins. Toxicon 1998, 36, 953–962. [Google Scholar] [CrossRef]
- Fischer, W.J.; Altheimer, S.; Cattori, V.; Meier, P.J.; Dietrich, D.R.; Hagenbuch, B. Organic anion transporting polypeptides expressed in liver and brain mediate uptake of microcystin. Toxicol. Appl. Pharmacol. 2005, 203, 257–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falconer, I.R.; Humpage, A.R. Cyanobacterial (blue-green algal) toxins in water supplies: Cylindrospermopsins. Environ. Toxicol. 2006, 21, 299–304. [Google Scholar] [CrossRef]
- Poniedziałek, B.; Rzymski, P.; Wiktorowicz, K. Toxicity of cylindrospermopsin in human lymphocytes: Proliferation, viability and cell cycle studies. Toxicol. Vitr. 2014, 28, 968–974. [Google Scholar] [CrossRef]
- Moreira, C.; Fathalli, A.; Vasconcelos, V.; Antunes, A. Phylogeny and biogeography of the invasive cyanobacterium Cylindrospermopsis raciborskii. Arch. Microbiol. 2014, 197, 47–52. [Google Scholar] [CrossRef]
- Moreira, C.; Mendes, R.; Azevedo, J.; Vasconcelos, V.; Antunes, A. First occurrence of cylindrospermopsin in Portugal: A contribution to its continuous global dispersal. Toxicon 2017, 130, 87–90. [Google Scholar] [CrossRef]
- Pichardo, S.; Cameán, A.M.; Jos, A. In vitro toxicological assessment of cylindrospermopsin: A review. Toxins (Basel) 2017, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Froscio, S.M.; Humpage, A.R.; Burcham, P.C.; Falconer, I.R. Cylindrospermopsin-induced protein synthesis inhibition and its dissociation from acute toxicity in mouse hepatocytes. Environ. Toxicol. 2003, 18, 243–251. [Google Scholar] [CrossRef]
- Prepas, E.E.; Kotak, B.G.; Campbell, L.M.; Evans, J.C.; Hrudey, S.E.; Holmes, C.F.B. Accumulation and elimination of cyanobacterial hepatotoxins by the freshwater clam Anodonta grandis simpsoniana. Can. J. Fish. Aquat. Sci. 1997, 54, 41–46. [Google Scholar] [CrossRef]
- Saker, M.L.; Metcalf, J.S.; Codd, G.A.; Vasconcelos, V.M. Accumulation and depuration of the cyanobacterial toxin cylindrospermopsin in the freshwater mussel Anodonta cygnea. Toxicon 2004, 43, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, A.; Park, H.D. Mechanism and prediction for contamination of freshwater bivalves (Unionidae) with the cyanobacterial toxin microcystin in hypereutrophic Lake Suwa, Japan. Environ. Toxicol. 2002, 17, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Juhel, G.; Davenport, J.; O’Halloran, J.; Culloty, S.C.; O’Riordan, R.M.; James, K.F.; Furey, A.; Allis, O. Impacts of microcystins on the feeding behaviour and energy balance of zebra mussels, Dreissena polymorpha: A bioenergetics approach. Aquat. Toxicol. 2006, 79, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Wu, F.; Yuan, M.; Liu, Q.; Wang, Y. Combined effects of toxic cyanobacteria Microcystis aeruginosa and hypoxia on the physiological responses of triangle sail mussel Hyriopsis cumingii. J. Hazard. Mater. 2016, 306, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Juhel, G.; O’Halloran, J.; Culloty, S.C.; O’Riordan, R.M.; Davenport, J.; O’Brien, N.M.; James, K.F.; Furey, A.; Allis, O. In vivo exposure to microcystins induces DNA damage in the haemocytes of the zebra mussel, Dreissena polymorpha, as measured with the comet assay. Environ. Mol. Mutagen. 2007, 48, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Gélinas, M.; Fortier, M.; Lajeunesse, A.; Fournier, M.; Gagnon, C.; Barnabé, S.; Gagné, F. Responses of freshwater mussel (Elliptio complanata) hemocytes exposed in vitro to crude extracts of Microcystis aeruginosa and Lyngbya wollei. Ecotoxicology 2014, 23, 260–266. [Google Scholar] [CrossRef]
- Martins, J.C.; Leão, P.N.; Vasconcelos, V. Differential protein expression in Corbicula fluminea upon exposure to a Microcystis aeruginosa toxic strain. Toxicon 2009, 53, 409–416. [Google Scholar] [CrossRef]
- Contardo-Jara, V.; Pflugmacher, S.; Wiegand, C. Multi-xenobiotic-resistance a possible explanation for the insensitivity of bivalves towards cyanobacterial toxins. Toxicon 2008, 52, 936–943. [Google Scholar] [CrossRef]
- Vasconcelos, V.M.; Wiegand, C.; Pflugmacher, S. Dynamics of glutathione-S-transferases in Mytilus galloprovincialis exposed to toxic Microcystis aeruginosa cells, extracts and pure toxins. Toxicon 2007, 50, 740–745. [Google Scholar] [CrossRef]
- Gérard, C.; Poullain, V.; Lance, E.; Acou, A.; Brient, L.; Carpentier, A. Influence of toxic cyanobacteria on community structure and microcystin accumulation of freshwater molluscs. Environ. Pollut. 2009, 157, 609–617. [Google Scholar] [CrossRef]
- Carneiro, M.; Antas, P.; Reis, B.; Azevedo, J.; Osório, H.; Campos, A.; Vasconcelos, V.; Martins, J.C. Modulation of hepatic glutathione transferases isoenzymes in three bivalve species exposed to purified microcystin-LR and Microcystis extracts. Toxicon 2017, 137, 150–157. [Google Scholar] [CrossRef]
- Kim, Y.D.; Kim, W.J.; Shin, Y.K.; Lee, D.H.; Kim, Y.J.; Kim, J.K.; Rhee, J.S. Microcystin-LR bioconcentration induces antioxidant responses in the digestive gland of two marine bivalves Crassostrea gigas and Mytilus edulis. Aquat. Toxicol. 2017, 188, 119–129. [Google Scholar] [CrossRef]
- Gibble, C.M.; Peacock, M.B.; Kudela, R.M. Evidence of freshwater algal toxins in marine shellfish: Implications for human and aquatic health. Harmful Algae 2016, 59, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Peacock, M.B.; Gibble, C.M.; Senn, D.B.; Cloern, J.E.; Kudela, R.M. Blurred lines: Multiple freshwater and marine algal toxins at the land-sea interface of San Francisco Bay, California. Harmful Algae 2018, 73, 138–147. [Google Scholar] [CrossRef]
- Tatters, A.O.; Howard, M.D.A.; Nagoda, C.; Busse, L.; Gellene, A.G.; Caron, D.A. Multiple stressors at the land-sea interface: Cyanotoxins at the land-sea interface in the Southern California Bight. Toxins (Basel) 2017, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Bormans, M.; Amzil, Z.; Mineaud, E.; Brient, L.; Savar, V.; Robert, E.; Lance, E. Demonstrated transfer of cyanobacteria and cyanotoxins along a freshwater-marine continuum in France. Harmful Algae 2019, 87, 101639. [Google Scholar] [CrossRef]
- Kim, D.; Hong, S.; Choi, H.; Choi, B.; Kim, J.; Khim, J.S.; Park, H.; Shin, K.H. Multimedia distributions, bioaccumulation, and trophic transfer of microcystins in the Geum River Estuary, Korea: Application of compound-specific isotope analysis of amino acids. Environ. Int. 2019, 133, 105194. [Google Scholar] [CrossRef]
- Miller, M.A.; Kudela, R.M.; Mekebri, A.; Crane, D.; Oates, S.C.; Tinker, M.T.; Staedler, M.; Miller, W.A.; Toy-Choutka, S.; Dominik, C.; et al. Evidence for a novel marine harmful algal bloom: Cyanotoxin (microcystin) transfer from land to sea otters. PLoS ONE 2010, 5, 1–11. [Google Scholar] [CrossRef]
- Des Aulnois, M.G.; Roux, P.; Caruana, A.; Réveillon, D.; Briand, E.; Hervé, F.; Savar, V.; Bormans, M.; Amzil, Z. Physiological and metabolic responses of freshwater and brackish-water strains of Microcystis aeruginosa acclimated to a salinity gradient: Insight into salt tolerance. Appl. Environ. Microbiol. 2019, 85, 1–38. [Google Scholar] [CrossRef]
- Campos, A.; Danielsson, G.; Farinha, A.P.; Kuruvilla, J.; Warholm, P.; Cristobal, S. Shotgun proteomics to unravel marine mussel (Mytilus edulis) response to long-term exposure to low salinity and propranolol in a Baltic Sea microcosm. J. Proteom. 2016, 137, 97–106. [Google Scholar] [CrossRef]
- Sylvester, F.; Dorado, J.; Boltovskoy, D.; Juárez, Á.; Cataldo, D. Filtration rates of the invasive pest bivalve Limnoperna fortunei as a function of Size and Temperature. Hydrobiologia 2005, 534, 71–80. [Google Scholar] [CrossRef]
- Pires, L.M.D.; Van Donk, E. Comparing grazing by Dreissena polymorpha on phytoplankton in the presence of toxic and non-toxic cyanobacteria. Freshw. Biol. 2002, 47, 1855–1865. [Google Scholar] [CrossRef]
- Dionisio Pires, L.M.; Ibelings, B.W.; Brehm, M.; Van Donk, E. Comparing grazing on lake seston by Dreissena and Daphnia: Lessons for biomanipulation. Microb. Ecol. 2005, 50, 242–252. [Google Scholar] [CrossRef]
- Cataldo, D.; Vinocur, A. The introduced bivalve Limnoperna fortunei boosts Microcystis growth in Salto Grande reservoir ( Argentina ): Evidence from mesocosm experiments. Hydrobiologia 2012, 680, 25–38. [Google Scholar] [CrossRef]
- Gazulha, V.; Mansur, M.; Cybis, L.; Azevedo, S. Feeding behavior of the invasive bivalve Limnoperna fortunei (Dunker, 1857) under exposure to toxic cyanobacteria Microcystis aeruginosa. Braz. J. Biol. 2012, 72, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Juhel, G.; Davenport, J.; O’Halloran, J.; Culloty, S.; Ramsay, R.; James, K.; Furey, A.; Allis, O. Pseudodiarrhoea in zebra mussels Dreissena polymorpha (Pallas) exposed to microcystins. J. Exp. Biol. 2006, 209, 810–816. [Google Scholar] [CrossRef] [Green Version]
- Gazulha, V.; Mansur, M.; Cybis, L.; Azevedo, S. Grazing impacts of the invasive bivalve Limnoperna fortunei (Dunker, 1857) on single-celled, colonial and filamentous cyanobacteria. Braz. J. Biol. 2012, 72, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Clearwater, S.J.; Wood, S.A.; Phillips, N.R.; Parkyn, S.M.; Van Ginkel, R.; Thompson, K.J. Toxicity thresholds for juvenile freshwater mussels Echyridella menziesii and crayfish Paranephrops planifrons, after acute or chronic exposure to Microcystis sp. Environ. Toxicol. 2014, 29, 487–502. [Google Scholar] [CrossRef]
- Hwang, S.J.; Kim, H.S.; Park, J.H.; Kim, B.H. Effects of cyanobacterium Microcystis aeruginosa on the filtration rate and mortality of the freshwater bivalve Corbicula leana. J. Environ. Biol. 2010, 31, 483–488. [Google Scholar]
- Juhel, G.; Ramsay, R.M.; Davenport, J.; Culloty, S.C. Effect of the microcystin-producing cyanobacterium, Microcystis aeruginosa, on immune functions of the zebra mussel Dreissena polymorpha. J. Shellfish Res. 2015, 34, 433–443. [Google Scholar] [CrossRef]
- Wu, F.; Kong, H.; Shang, Y.; Zhou, Z.; Gul, Y.; Liu, Q.; Hu, M. Histopathological alterations in triangle sail mussel ( Hyriopsis cumingii ) exposed to toxic cyanobacteria ( Microcystis aeruginosa ) under hypoxia. Aquaculture 2017, 467, 182–189. [Google Scholar] [CrossRef]
- Jin, C.H.; Zheng, Z.M.; Lu, K.H.; Zheng, C.P. Effects of Microcystis aeruginosa on Oxygen Consumption and Ammonia Excretion of Freshwater Mussel, Hyriopsis cumingii. Ningbo Daxue Xuebao Ligong Ban 2011, 24, 5–9. [Google Scholar]
- Kim, M.S.; Lee, Y.J.; Ha, S.Y.; Kim, B.H.; Hwang, S.J.; Kwon, J.T.; Choi, J.W.; Shin, K.H. Accumulation of microcystin (LR, RR and YR) in three freshwater bivalves in Microcystis aeruginosa bloom using dual isotope tracer. Mar. Drugs 2017, 15, 226. [Google Scholar] [CrossRef] [Green Version]
- Amorim, Á.; Vasconcelos, V. Dynamics of microcystins in the mussel Mytilus galloprovincialis. Toxicon 1999, 37, 1041–1052. [Google Scholar] [CrossRef]
- Marie, B.; Arivalagan, J.; Mathéron, L.; Bolbach, G.; Berland, S.; Marie, A.; Marin, F. Deep conservation of bivalve nacre proteins highlighted by shell matrix proteomics of the Unionoida Elliptio complanata and Villosa lienosa. J. R. Soc. Interface 2017, 14, 20160846. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Fan, G.; Jiao, Y.; Zhang, H.; Guo, X.; Huang, R.; Zheng, Z.; Bian, C.; Deng, Y.; Wang, Q.; et al. The pearl oyster Pinctada fucata martensii genome and multi-omic analyses provide insights into biomineralization. Gigascience 2017, 6, 1–12. [Google Scholar] [CrossRef]
- Young, F.M.; Zebian, D.; Froscio, S.; Humpage, A. Cylindrospermopsin, a blue-green algal toxin, inhibited human luteinised granulosa cell protein synthesis in vitro. Toxicol. Vitr. 2012, 26, 656–662. [Google Scholar] [CrossRef]
- Froscio, S.M.; Humpage, A.R.; Wickramasinghe, W.; Shaw, G.; Falconer, I.R. Interaction of the cyanobacterial toxin cylindrospermopsin with the eukaryotic protein synthesis system. Toxicon 2008, 51, 191–198. [Google Scholar] [CrossRef]
- Puerto, M.; Campos, A.; Prieto, A.; Cameán, A.; Almeida, A.M.; de Coelho, A.V.; Vasconcelos, V. Differential protein expression in two bivalve species; Mytilus galloprovincialis and Corbicula fluminea; exposed to Cylindrospermopsis raciborskii cells. Aquat. Toxicol. 2011, 101, 109–116. [Google Scholar] [CrossRef]
- Antas, P.; Carneiro, M.; Reis, B.; Castelo-Branco, R.; Azevedo, J.; Urbatzka, R.; Campos, A.; Vasconcelos, V.; Martins, J.C. GST transcriptional changes induced by a toxic Microcystis aeruginosa strain in two bivalve species during exposure and recovery phases. Ecotoxicology 2018, 27, 1272–1280. [Google Scholar] [CrossRef]
- Bednarska, A.; Pietrzak, B.; Pijanowska, J. Effect of poor manageability and low nutritional value of cyanobacteria on Daphnia magna life history performance. J. Plankton Res. 2014, 36, 838–847. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, K.; Jónasdóttir, S.H. Nutritional quality of two cyanobacteria: How rich is “poor” food? Mar. Ecol. Prog. Ser. 1997, 151, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Huang, I.S.; Zimba, P.V. Cyanobacterial bioactive metabolites—A review of their chemistry and biology. Harmful Algae 2019, 83, 42–94. [Google Scholar] [CrossRef]
- Namikoshi, M.; Rinehart, K.L. Bioactive compounds produced by cyanobacteria. J. Ind. Microbiol. Biotechnol. 1996, 17, 373–384. [Google Scholar] [CrossRef]
- Ramos, V.; Morais, J.; Castelo-branco, R.; Pinheiro, Â.; Martins, J.; Regueiras, A.; Pereira, A.L.; Lopes, V.R.; Frazão, B.; Gomes, D.; et al. Cyanobacterial diversity held in microbial biological resource centers as a biotechnological asset: The case study of the newly established LEGE culture collection. J. Appl. Phycol. 2018, 30, 1437–1451. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, C.; Azevedo, J.; Campos, A.; Loureiro, S.; Vasconcelos, V. Absence of negative allelopathic effects of cylindrospermopsin and microcystin-LR on selected marine and freshwater phytoplankton species. Hydrobiologia 2013, 705, 27–42. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.; Azevedo, J.; Campos, A.; Vasconcelos, V.; Pereira, A.L. Biochemical and growth performance of the aquatic macrophyte Azolla filiculoides to sub-chronic exposure to cylindrospermopsin. Ecotoxicology 2015, 24, 1848–1857. [Google Scholar] [CrossRef]
- Pereira, A.L.; Santos, C.; Azevedo, J.; Martins, T.P.; Castelo-Branco, R.; Ramos, V.; Vasconcelos, V.; Campos, A. Effects of two toxic cyanobacterial crude extracts containing microcystin-LR and cylindrospermopsin on the growth and photosynthetic capacity of the microalga Parachlorella kessleri. Algal Res. 2018, 34, 198–208. [Google Scholar] [CrossRef]
- Barker Jørgensen, C. Feeding and cleaning mechanisms in the suspension feeding bivalve Mytilus edulis. Mar. Biol. 1981, 65, 159–163. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.; Hamilton, J.K.; Rebers, P.A.; Smith, F. A colorimetric method for the determination of sugars. Nature 1951, 168, 350–356. [Google Scholar] [CrossRef]
- Freitas, M.; Azevedo, J.; Carvalho, A.P.; Campos, A.; Vasconcelos, V. Effects of storage, processing and proteolytic digestion on microcystin-LR concentration in edible clams. Food Chem. Toxicol. 2014, 66, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Freitas, M.; Azevedo, J.; Carvalho, A.P.; Mendes, V.M.; Manadas, B.; Campos, A.; Vasconcelos, V. Bioaccessibility and changes on cylindrospermopsin concentration in edible mussels with storage and processing time. Food Control 2016, 59, 567–574. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Guillén, R.; Moreno, I.; Prieto Ortega, A.I.; Eugenia Soria-Díaz, M.; Vasconcelos, V.; Cameán, A.M. CYN determination in tissues from freshwater fish by LC-MS/MS: Validation and application in tissues from subchronically exposed tilapia (Oreochromis niloticus). Talanta 2015, 131, 452–459. [Google Scholar] [CrossRef]
- Díez-Quijada, L.; Guzmán-Guillén, R.; Prieto Ortega, A.I.; Llana-Ruíz-Cabello, M.; Campos, A.; Vasconcelos, V.; Jos, Á.; Cameán, A.M. New Method for Simultaneous Determination of Microcystins and Cylindrospermopsin in Vegetable Matrices by SPE-UPLC-MS/MS. Toxins (Basel) 2018, 10, 406. [Google Scholar] [CrossRef] [Green Version]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–363. [Google Scholar] [CrossRef]
- Domínguez-Pérez, D.; Rodríguez, A.A.; Osorio, H.; Azevedo, J.; Castañeda, O.; Vasconcelos, V.; Antunes, A. Microcystin-lr detected in a low molecular weight fraction from a crude extract of Zoanthus sociatus. Toxins (Basel) 2017, 9, 89. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.r-project.org/ (accessed on 27 January 2020).









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, F.; Diez-Quijada, L.; Turkina, M.V.; Morais, J.; Felpeto, A.B.; Azevedo, J.; Jos, A.; Camean, A.M.; Vasconcelos, V.; Martins, J.C.; et al. Physiological and Metabolic Responses of Marine Mussels Exposed to Toxic Cyanobacteria Microcystis aeruginosa and Chrysosporum ovalisporum. Toxins 2020, 12, 196. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12030196
Oliveira F, Diez-Quijada L, Turkina MV, Morais J, Felpeto AB, Azevedo J, Jos A, Camean AM, Vasconcelos V, Martins JC, et al. Physiological and Metabolic Responses of Marine Mussels Exposed to Toxic Cyanobacteria Microcystis aeruginosa and Chrysosporum ovalisporum. Toxins. 2020; 12(3):196. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12030196
Chicago/Turabian StyleOliveira, Flavio, Leticia Diez-Quijada, Maria V. Turkina, João Morais, Aldo Barreiro Felpeto, Joana Azevedo, Angeles Jos, Ana M. Camean, Vitor Vasconcelos, José Carlos Martins, and et al. 2020. "Physiological and Metabolic Responses of Marine Mussels Exposed to Toxic Cyanobacteria Microcystis aeruginosa and Chrysosporum ovalisporum" Toxins 12, no. 3: 196. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12030196