Shedding Light on the Venom Proteomes of the Allergy-Relevant Hymenoptera Polistes dominula (European Paper Wasp) and Vespula spp. (Yellow Jacket)

 , , , , , and
, , , , , and
Abstract
:1. Introduction
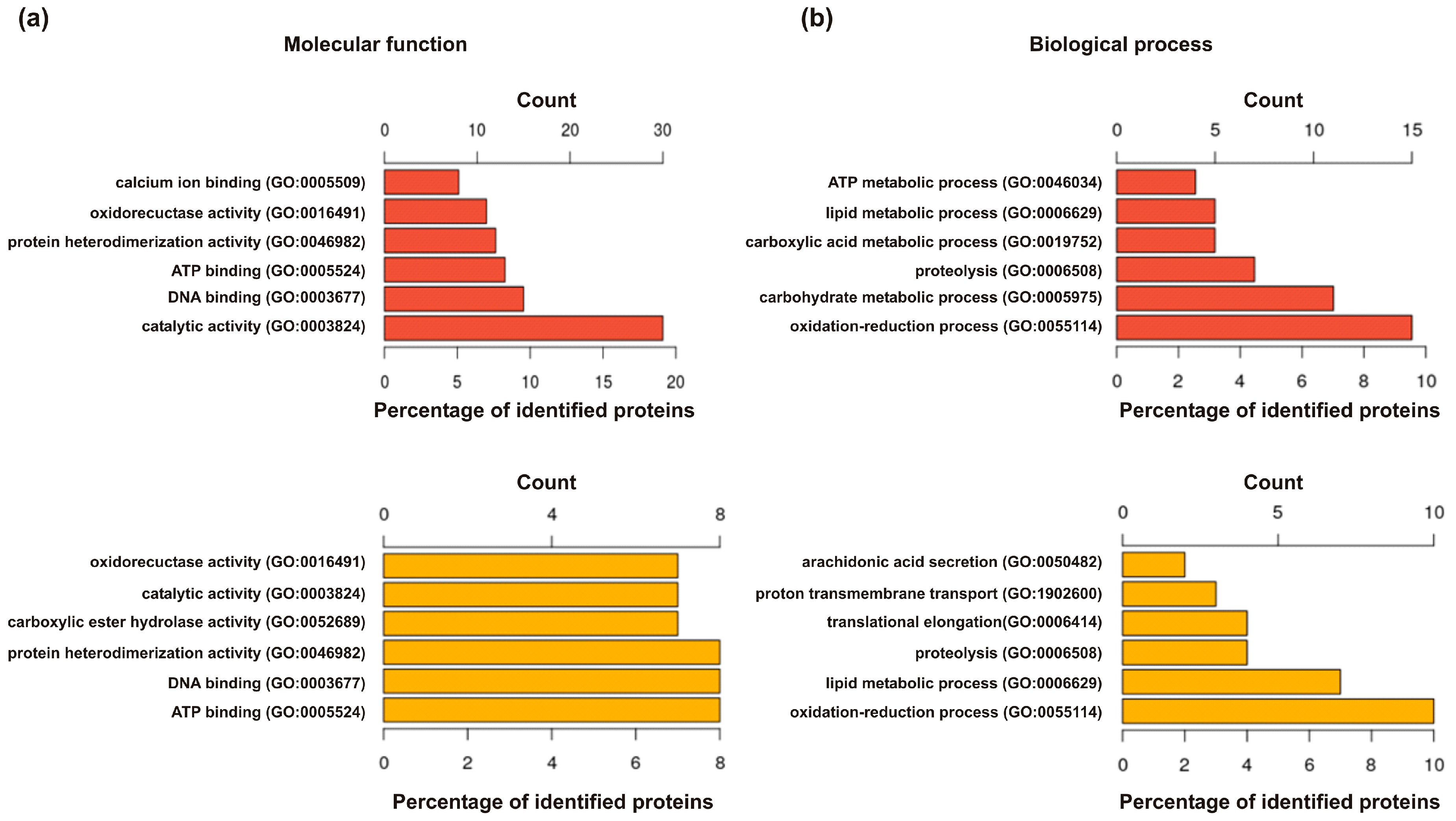
2. Results
2.1. Identification of Already Annotated Allergens
2.2. Identification of Secreted Proteins and Proteins with Known Function in the Venom
2.3. Identification of Venom Trace Molecules without Signal Peptides
3. Discussion
4. Materials and Methods
4.1. Sample Preparation
4.2. Mass Spectrometric Analysis
4.3. Data Processing
4.4. Data Availability
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Worm, M.; Moneret-Vautrin, A.; Scherer, K.; Lang, R.; Fernandez-Rivas, M.; Cardona, V.; Kowalski, M.L.; Jutel, M.; Poziomkowska-Gesicka, I.; Papadopoulos, N.G.; et al. First European data from the network of severe allergic reactions (NORA). Allergy 2014, 69, 1397–1404. [Google Scholar] [CrossRef]
- Bilo, M.B.; Ollert, M.; Blank, S. The role of component-resolved diagnosis in Hymenoptera venom allergy. Curr. Opin. Allergy Clin. Immunol. 2019, 19, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Blank, S.; Bilo, M.B.; Ollert, M. Component-resolved diagnostics to direct in venom immunotherapy: Important steps towards precision medicine. Clin. Exp. Allergy 2018, 48, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Spillner, E.; Blank, S.; Jakob, T. Hymenoptera allergens: From venom to “venome”. Front. Immunol. 2014, 5, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervo, R.; Zacchi, F.; Turillazzi, S. Polistes dominulus (Hymenoptera, Vespidae) invading North America: Some hypotheses for its rapid spread. Insectes Soc. 2000, 47, 155–157. [Google Scholar] [CrossRef]
- Smit, J. De veldwespen Polistes dominulus en P. biglumis op in Nederland (Hymenoptera: Vespidae). Nederl. Faun. Med. 2003, 18, 81–88. [Google Scholar]
- Eardley, C.; Koch, F.; Wood, A.R. Polistes dominulus (Christ, 1791) (Hymenoptera: Polistinae: Vespidae) newly recorded from South Africa: Short communication. Afr. Entomol. 2009, 17, 226–227. [Google Scholar] [CrossRef]
- Buck, M.; Marshall, S.A.; Cheung, D.K.B. Identification atlas of the Vespidae (Hymenoptera, Aculeata) of the northeastern Nearctic region. Can. J. Arthropod Identif. 2008, 5, 1–492. [Google Scholar] [CrossRef]
- Sackmann, P.; Villacide, J.M.; Corley, J. Presencia de una nueva avispa social exótica, Polistes dominulus (Hymenoptera: Vespidae) en la Patagonia Argentina. Rev. Soc. Entomol. Arge. 2003, 62, 72–74. [Google Scholar]
- Demain, J.G.; Gessner, B.D.; McLaughlin, J.B.; Sikes, D.S.; Foote, J.T. Increasing insect reactions in Alaska: Is this related to changing climate? Allergy Asthma Proc. 2009, 30, 238–243. [Google Scholar] [CrossRef] [Green Version]
- Oswalt, M.L.; Foote, J.T.; Kemp, S.F. Anaphylaxis: Report of two fatal yellow jacket (YJ) stings in Alaska (AK). J. Allergy Clin. Immunol. 2007, 119, S34. [Google Scholar] [CrossRef]
- Fitzgerald, K.T.; Flood, A.A. Hymenoptera stings. Clin. Tech. Small Anim. Pr. 2006, 21, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, D.R.; Jacobson, R.S. Allergens in hymenoptera venom XII: How much protein is in a sting? Ann. Allergy 1984, 52, 276–278. [Google Scholar] [PubMed]
- Schumacher, M.J.; Tveten, M.S.; Egen, N.B. Rate and quantity of delivery of venom from honeybee stings. J. Allergy Clin. Immunol. 1994, 93, 831–835. [Google Scholar] [CrossRef]
- Irazazabal, L.N.; Porto, W.F.; Ribeiro, S.M.; Casale, S.; Humblot, V.; Ladram, A.; Franco, O.L. Selective amino acid substitution reduces cytotoxicity of the antimicrobial peptide mastoparan. Biochim. Biophys. Acta 2016, 1858, 2699–2708. [Google Scholar] [CrossRef] [PubMed]
- Leite, N.B.; Aufderhorst-Roberts, A.; Palma, M.S.; Connell, S.D.; Neto, R.J.; Beales, P.A. PE and PSps lipids synergistically enhance membrane poration by a peptide with anticancer properties. Biophys. J. 2015, 109, 936–947. [Google Scholar] [CrossRef] [Green Version]
- Van Vaerenbergh, M.; Debyser, G.; Devreese, B.; de Graaf, D.C. Exploring the hidden honeybee (Apis mellifera) venom proteome by integrating a combinatorial peptide ligand library approach with FTMS. J. Proteom. 2014, 99, 169–178. [Google Scholar] [CrossRef]
- Dos Santos-Pinto, J.R.; Fox, E.G.; Saidemberg, D.M.; Santos, L.D.; da Silva-Menegasso, A.R.; Costa-Manso, E.; Machado, E.A.; Bueno, O.C.; Palma, M.S. Proteomic view of the venom from the fire ant Solenopsis invicta Buren. J. Proteome Res. 2012, 11, 4643–4653. [Google Scholar] [CrossRef]
- De Souza, C.L.; Dos Santos-Pinto, J.R.A.; Esteves, F.G.; Perez-Riverol, A.; Fernandes, L.G.R.; de Lima Zollner, R.; Palma, M.S. Revisiting Polybia paulista wasp venom using shotgun proteomics-insights into the N-linked glycosylated venom proteins. J. Proteom. 2019, 200, 60–73. [Google Scholar] [CrossRef]
- Harris, J.B.; Scott-Davey, T. Secreted phospholipases A2 of snake venoms: Effects on the peripheral neuromuscular system with comments on the role of phospholipases A2 in disorders of the CNS and their uses in industry. Toxins 2013, 5, 2533–2571. [Google Scholar] [CrossRef] [Green Version]
- Habermann, E. Bee and wasp venoms. Science 1972, 177, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Markovic-Housley, Z.; Miglierini, G.; Soldatova, L.; Rizkallah, P.J.; Muller, U.; Schirmer, T. Crystal structure of hyaluronidase, a major allergen of bee venom. Structure 2000, 8, 1025–1035. [Google Scholar] [CrossRef]
- Schiener, M.; Eberlein, B.; Moreno-Aguilar, C.; Pietsch, G.; Serrano, P.; McIntyre, M.; Schwarze, L.; Russkamp, D.; Biedermann, T.; Spillner, E.; et al. Application of recombinant antigen 5 allergens from seven allergy-relevant Hymenoptera species in diagnostics. Allergy 2017, 72, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Radauer, C.; Nandy, A.; Ferreira, F.; Goodman, R.E.; Larsen, J.N.; Lidholm, J.; Pomes, A.; Raulf-Heimsoth, M.; Rozynek, P.; Thomas, W.R.; et al. Update of the WHO/IUIS allergen nomenclature database based on analysis of allergen sequences. Allergy 2014, 69, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, G.M.; Roelants, K.; O’Bryan, M.K. The CAP superfamily: Cysteine-rich secretory proteins, antigen 5, and pathogenesis-related 1 proteins-roles in reproduction, cancer, and immune defense. Endocr. Rev. 2008, 29, 865–897. [Google Scholar] [CrossRef] [PubMed]
- Assumpcao, T.C.; Ma, D.; Schwarz, A.; Reiter, K.; Santana, J.M.; Andersen, J.F.; Ribeiro, J.M.; Nardone, G.; Yu, L.L.; Francischetti, I.M. Salivary antigen-5/CAP family members are Cu2+-dependent antioxidant enzymes that scavenge O2⨪ and inhibit collagen-induced platelet aggregation and neutrophil oxidative burst. J. Biol. Chem. 2013, 288, 14341–14361. [Google Scholar] [CrossRef] [Green Version]
- Morrissette, J.; Kratzschmar, J.; Haendler, B.; el-Hayek, R.; Mochca-Morales, J.; Martin, B.M.; Patel, J.R.; Moss, R.L.; Schleuning, W.D.; Coronado, R.; et al. Primary structure and properties of helothermine, a peptide toxin that blocks ryanodine receptors. Biophys. J. 1995, 68, 2280–2288. [Google Scholar] [CrossRef] [Green Version]
- Nobile, M.; Magnelli, V.; Lagostena, L.; Mochca-Morales, J.; Possani, L.D.; Prestipino, G. The toxin helothermine affects potassium currents in newborn rat cerebellar granule cells. J. Membr. Biol. 1994, 139, 49–55. [Google Scholar] [CrossRef]
- Nobile, M.; Noceti, F.; Prestipino, G.; Possani, L.D. Helothermine, a lizard venom toxin, inhibits calcium current in cerebellar granules. Exp. Brain Res. 1996, 110, 15–20. [Google Scholar] [CrossRef]
- Monsalve, R.I.; Vega, A.; Marques, L.; Miranda, A.; Fernandez, J.; Soriano, V.; Cruz, S.; Dominguez-Noche, C.; Sanchez-Morillas, L.; Armisen-Gil, M.; et al. Component-resolved diagnosis of vespid venom-allergic individuals: Phospholipases and antigen 5s are necessary to identify Vespula or Polistes sensitization. Allergy 2012, 67, 528–536. [Google Scholar] [CrossRef]
- Aertgeerts, K.; Ye, S.; Tennant, M.G.; Kraus, M.L.; Rogers, J.; Sang, B.C.; Skene, R.J.; Webb, D.R.; Prasad, G.S. Crystal structure of human dipeptidyl peptidase IV in complex with a decapeptide reveals details on substrate specificity and tetrahedral intermediate formation. Protein Sci. 2004, 13, 412–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blank, S.; Seismann, H.; Bockisch, B.; Braren, I.; Cifuentes, L.; McIntyre, M.; Ruhl, D.; Ring, J.; Bredehorst, R.; Ollert, M.W.; et al. Identification, recombinant expression, and characterization of the 100 kDa high molecular weight Hymenoptera venom allergens Api m 5 and Ves v 3. J. Immunol. 2010, 184, 5403–5413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreil, G.; Haiml, L.; Suchanek, G. Stepwise cleavage of the pro part of promelittin by dipeptidylpeptidase IV. Evidence for a new type of precursor-product conversion. Eur. J. Biochem. 1980, 111, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Lee, V.S.; Tu, W.C.; Jinn, T.R.; Peng, C.C.; Lin, L.J.; Tzen, J.T. Molecular cloning of the precursor polypeptide of mastoparan B and its putative processing enzyme, dipeptidyl peptidase IV, from the black-bellied hornet, Vespa basalis. Insect Mol. Biol. 2007, 16, 231–237. [Google Scholar] [CrossRef]
- Schiener, M.; Hilger, C.; Eberlein, B.; Pascal, M.; Kuehn, A.; Revets, D.; Planchon, S.; Pietsch, G.; Serrano, P.; Moreno-Aguilar, C.; et al. The high molecular weight dipeptidyl peptidase IV Pol d 3 is a major allergen of Polistes dominula venom. Sci. Rep. 2018, 8, 1318. [Google Scholar] [CrossRef]
- Pantera, B.; Hoffman, D.R.; Carresi, L.; Cappugi, G.; Turillazzi, S.; Manao, G.; Severino, M.; Spadolini, I.; Orsomando, G.; Moneti, G.; et al. Characterization of the major allergens purified from the venom of the paper wasp Polistes gallicus. Biochim. Biophys. Acta 2003, 1623, 72–81. [Google Scholar] [CrossRef]
- Winningham, K.M.; Fitch, C.D.; Schmidt, M.; Hoffman, D.R. Hymenoptera venom protease allergens. J. Allergy Clin. Immunol. 2004, 114, 928–933. [Google Scholar] [CrossRef]
- Vaiyapuri, S.; Wagstaff, S.C.; Harrison, R.A.; Gibbins, J.M.; Hutchinson, E.G. Evolutionary analysis of novel serine proteases in the venom gland transcriptome of Bitis gabonica rhinoceros. PLoS ONE 2011, 6, e21532. [Google Scholar] [CrossRef] [Green Version]
- Almagro-Armenteros, J.J.; Tsirigos, K.D.; Sonderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Blank, S.; Seismann, H.; Michel, Y.; McIntyre, M.; Cifuentes, L.; Braren, I.; Grunwald, T.; Darsow, U.; Ring, J.; Bredehorst, R.; et al. Api m 10, a genuine A. mellifera venom allergen, is clinically relevant but underrepresented in therapeutic extracts. Allergy 2011, 66, 1322–1329. [Google Scholar] [CrossRef]
- Peiren, N.; de Graaf, D.C.; Brunain, M.; Bridts, C.H.; Ebo, D.G.; Stevens, W.J.; Jacobs, F.J. Molecular cloning and expression of icarapin, a novel IgE-binding bee venom protein. FEBS Lett. 2006, 580, 4895–4899. [Google Scholar] [CrossRef] [Green Version]
- Frick, M.; Muller, S.; Bantleon, F.; Huss-Marp, J.; Lidholm, J.; Spillner, E.; Jakob, T. rApi m 3 and rApi m 10 improve detection of honey bee sensitization in Hymenoptera venom-allergic patients with double sensitization to honey bee and yellow jacket venom. Allergy 2015, 70, 1665–1668. [Google Scholar] [CrossRef] [PubMed]
- Atakuziev, B.U.; Nuritova, F.; Usmanov, P.B. Phospholipase A2 from the venom of the spider Eresus niger. Chem. Nat. Compd. 1991, 27, 487–489. [Google Scholar] [CrossRef]
- Kohler, J.; Blank, S.; Muller, S.; Bantleon, F.; Frick, M.; Huss-Marp, J.; Lidholm, J.; Spillner, E.; Jakob, T. Component resolution reveals additional major allergens in patients with honeybee venom allergy. J. Allergy Clin. Immunol. 2014, 133, 1383–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamazaki, Y.; Takani, K.; Atoda, H.; Morita, T. Snake venom vascular endothelial growth factors (VEGF) exhibit potent activity through their specific recognition of KDR (VEGF receptor 2). J. Biol. Chem. 2003, 278, 51985–51988. [Google Scholar] [CrossRef] [Green Version]
- Peiren, N.; Vanrobaeys, F.; de Graaf, D.C.; Devreese, B.; Van Beeumen, J.; Jacobs, F.J. The protein composition of honeybee venom reconsidered by a proteomic approach. Biochim. Biophys. Acta 2005, 1752, 1–5. [Google Scholar] [CrossRef]
- Russkamp, D.; Van Vaerenbergh, M.; Etzold, S.; Eberlein, B.; Darsow, U.; Schiener, M.; De Smet, L.; Absmaier, M.; Biedermann, T.; Spillner, E.; et al. Characterization of the honeybee venom proteins C1q-like protein and PVF1 and their allergenic potential. Toxicon 2018, 150, 198–206. [Google Scholar] [CrossRef] [Green Version]
- Turillazzi, S.; Mastrobuoni, G.; Dani, F.R.; Moneti, G.; Pieraccini, G.; la Marca, G.; Bartolucci, G.; Perito, B.; Lambardi, D.; Cavallini, V.; et al. Dominulin A and B: Two new antibacterial peptides identified on the cuticle and in the venom of the social paper wasp Polistes dominulus using MALDI-TOF, MALDI-TOF/TOF, and ESI-ion trap. J. Am. Soc. Mass Spectrom. 2006, 17, 376–383. [Google Scholar] [CrossRef] [Green Version]
- Park, N.G.; Yamato, Y.; Lee, S.; Sugihara, G. Interaction of mastoparan-B from venom of a hornet in Taiwan with phospholipid bilayers and its antimicrobial activity. Biopolymers 1995, 36, 793–801. [Google Scholar] [CrossRef]
- Yoshida, H.; Geller, R.G.; Pisano, J.J. Vespulakinins: New carbohydrate-containing bradykinin derivatives. Biochemistry 1976, 15, 61–64. [Google Scholar] [CrossRef]
- Franz, M.; Mense, S. Muscle receptors with group IV afferent fibres responding to application of bradykinin. Brain Res. 1975, 92, 369–383. [Google Scholar] [CrossRef]
- Ollert, M.; Blank, S. Anaphylaxis to insect venom allergens: Role of molecular diagnostics. Curr. Allergy Asthma Rep. 2015, 15, 527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanca, M.; Garcia, F.; Miranda, A.; Carmona, M.J.; Garcia, J.; Fernandez, J.; Terrados, S.; Vega, J.M.; Juarez, C. Determination of IgE antibodies to Polistes dominulus, Vespula germanica and Vespa crabro in sera of patients allergic to vespids. Allergy 1991, 46, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.A.; Rahr, R.; Thueson, D.O.; Lett-Brown, M.A.; Hokanson, J.A.; Yunginger, J.W. Diagnosis of Polistes wasp hypersensitivity. J. Allergy Clin. Immunol. 1983, 72, 399–406. [Google Scholar] [CrossRef]
- Severino, M.G.; Campi, P.; Macchia, D.; Manfredi, M.; Turillazzi, S.; Spadolini, I.; Bilo, M.B.; Bonifazi, F. European Polistes venom allergy. Allergy 2006, 61, 860–863. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Bonilla, P.A.; Galan-Nieto, A.; Alfaya-Arias, T.; Garcia-Rodriguez, C.; de la Roca-Pinzon, F.; Feo-Brito, F. Component-resolved diagnosis in vespid venom-allergic individuals. Allergol. Immunopathol. 2015, 43, 398–402. [Google Scholar] [CrossRef]
- Caruso, B.; Bonadonna, P.; Bovo, C.; Melloni, N.; Lombardo, C.; Senna, G.; Lippi, G. Wasp venom allergy screening with recombinant allergen testing. Diagnostic performance of rPol d 5 and rVes v 5 for differentiating sensitization to Vespula and Polistes subspecies. Clin. Chim. Acta 2016, 453, 170–173. [Google Scholar] [CrossRef]
- Savi, E.; Peveri, S.; Makri, E.; Pravettoni, V.; Incorvaia, C. Comparing the ability of molecular diagnosis and CAP-inhibition in identifying the really causative venom in patients with positive tests to Vespula and Polistes species. Clin. Mol. Allergy 2016, 14, 3. [Google Scholar] [CrossRef] [Green Version]
- Quercia, O.; Cova, V.; Martini, M.; Cortellini, G.; Murzilli, F.; Bignardi, D.; Cilia, M.; Scarpa, A.; Bilo, M.B. CAP-inhibition, molecular diagnostics, and total IgE in the evaluation of Polistes and Vespula double sensitization. Int. Arch. Allergy Immunol. 2018, 177, 365–369. [Google Scholar] [CrossRef]
- Heneberg, P.; Riegerova, K.; Kucera, P. Pimecrolimus is a potent inhibitor of allergic reactions to hymenopteran venom extracts and birch pollen allergen in vitro. PLoS ONE 2015, 10, e0142953. [Google Scholar] [CrossRef]
- Eberlein, B.; Krischan, L.; Darsow, U.; Ollert, M.; Ring, J. Double positivity to bee and wasp venom: Improved diagnostic procedure by recombinant allergen-based ige testing and basophil activation test including data about cross-reactive carbohydrate determinants. J. Allergy Clin. Immunol. 2012, 130, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.S.; Foehr, S.; Garfield, D.A.; Furlong, E.E.; Steinmetz, L.M.; Krijgsveld, J. Ultrasensitive proteome analysis using paramagnetic bead technology. Mol. Syst. Biol. 2014, 10, 757. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.Z.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Identifier | Description | Allergen |
|---|---|---|
| AAT95010.1 | Allergen Pol d 5 precursor | Pol d 5 |
| NP_001310265.1 | Antigen 5 precursor | Pol d 5 |
| XP_015174448.1 | Dipeptidyl peptidase IV | Pol d 3/Ves v 3 |
| ACA00159.1 | Dipeptidylpeptidase IV preproprotein | Pol d 3/Ves v 3 |
| XP_015179722.1 | Hyaluronidase | Pol d 2 |
| P49370 | Hyaluronidase A | Ves v 2.0101 |
| Q5D7H4.1 | Hyaluronidase B | Ves v 2.0201 |
| AAS67042.1 | Phospholipase A1 2 precursor, partial | Pol d 1 |
| AAS67043.1 | Phospholipase A1 3 precursor, partial | Pol d 1 |
| Q6Q249.1 | Phospholipase A1 4 | Pol d 1 |
| AAS67044.1 | Phospholipase A1 4 precursor, partial | Pol d 1 |
| Q3ZU95 | Phospholipase A1 | Ves g 1 |
| P49369 | Phospholipase A1 | Ves v 1 |
| P35784 | Venom allergen 5 | Ves g 5 |
| Q05110 | Venom allergen 5 | Ves v 5 |
| NP_001310266.1 | Venom serine protease precursor | Pol d 4 |
| Identifier | Description | PDV | YJV |
|---|---|---|---|
| XP_015182550.1 | 60S acidic ribosomal protein P2 | X | |
| XP_015183814.1 | Acidic phospholipase A2 PA4-like isoform X1 | X | |
| XP_015183815.1 | Acidic phospholipase A2 PA4-like isoform X2 [P. dominula] | X | X |
| XP_015181576.1 | Alpha-N-acetylgalactosaminidase isoform X1 [P. dominula] | X | |
| XP_015181584.1 | Alpha-N-acetylgalactosaminidase isoform X2 [P. dominula] | X | |
| XP_015184655.1 | Apolipophorins [P. dominula] | X | X |
| XP_015183810.1 | Apolipoprotein D-like [P. dominula] | X | |
| XP_015179728.1 | Cyclin-dependent kinase 8-like [P. dominula] | X | |
| XP_015182852.1 | Digestive cysteine proteinase 1 [P. dominula] | X | |
| P0C1M6.1 | Dominulin-A | X | |
| P0C1M7.1 | Dominulin-B | X | |
| XP_015185362.1 | FK506-binding protein 2 isoform X1 [P. dominula] | X | |
| XP_015185363.1 | FK506-binding protein 2 isoform X2 [P. dominula] | X | |
| XP_015183963.1 | Furin-like protease 1 isoform 1-CRR isoform X1 [P. dominula] | X | |
| XP_015183982.1 | Furin-like protease 1 isoform 1-CRR isoform X3 [P. dominula] | X | |
| XP_015179268.1 | Heat shock 70 kDa protein cognate 3 isoform X1 [P. dominula] | X | X |
| XP_015185877.1 | Icarapin-like | X | |
| XP_015186100.1 | Interferon-related developmental regulator 1-like [P. dominula] | X | |
| XP_015187740.1 | Juvenile hormone epoxide hydrolase 1-like | X | |
| XP_015183311.1 | Maltase 1-like [P. dominula] | X | |
| P01514 | Mastoparan-L [Vespula lewisii] * | X | |
| P0C1Q8 | Mastoparan-V1 [V. vulgaris] * | X | |
| A0A310SDE3 | Neuroblastoma suppressor of tumorigenicity 1 [Eufriesea mexicana] | X | |
| XP_015173581.1 | Nucleobindin-2-like isoform X1 [P. dominula] | X | |
| XP_015173583.1 | Nucleobindin-2-like isoform X2 [P. dominula] | X | |
| XP_015173585.1 | Nucleobindin-2-like isoform X3 [P. dominula] | X | |
| XP_015173600.1 | Peptidylglycine alpha-hydroxylating monooxygenase [P. dominula] * | X | |
| XP_015187119.1 | Phospholipase A1-like [P. dominula] | X | X |
| XP_015190124.1 | Phospholipid hydroperoxide glutathione peroxidase isoform X2 [P. dominula] | X | |
| XP_015186079.1 | Prosaposin isoform X1 [P. dominula] | X | |
| XP_015186089.1 | Prosaposin isoform X2 [P. dominula] | X | |
| XP_015191732.1 | Protein 5NUC-like [P. dominula] | X | |
| XP_015181251.1 | Protein D2-like [P. dominula] | X | |
| XP_015189991.1 | Protein disulfide-isomerase A3 [P. dominula] | X | |
| XP_015190044.1 | Protein lethal(2)essential for life-like | X | |
| XP_015171801.1 | Renin receptor isoform X1 [P. dominula] | X | |
| XP_015171802.1 | Renin receptor isoform X2 [P. dominula] | X | |
| XP_015171803.1 | Renin receptor isoform X3 [P. dominula] | X | |
| XP_015171804.1 | Renin receptor isoform X4 [P. dominula] | X | |
| AAP37412.1 | Serine protease precursor | X | |
| A0A1B1JID1 | Superoxide dismutase [Cu-Zn] [A. mellifera] * | X | |
| A0A0M9A935 | Transferrin [Melipona quadrifasciata] * | X | |
| XP_015185020.1 | Tyramine beta-hydroxylase [P. dominula] | X | |
| XP_015179456.1 | Uncharacterized protein LOC107067990 | X | |
| XP_015181315.1 | Uncharacterized protein LOC107068952 | X | |
| XP_015183315.1 | Uncharacterized protein LOC107070028 [P. dominula] | X | |
| XP_015185303.1 | Vascular endothelial growth factor C | X | |
| D0EY66 | Venom CUB-protease [V. vulgaris] | X | |
| A0A3S6I504 | Vespakinin [Parapolybia varia] * | X | |
| P57672 | Vespulakinin-1 [Vespula maculifrons] * | X |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grosch, J.; Hilger, C.; Bilò, M.B.; Kler, S.; Schiener, M.; Dittmar, G.; Bernardin, F.; Lesur, A.; Ollert, M.; Schmidt-Weber, C.B.; et al. Shedding Light on the Venom Proteomes of the Allergy-Relevant Hymenoptera Polistes dominula (European Paper Wasp) and Vespula spp. (Yellow Jacket). Toxins 2020, 12, 323. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12050323
Grosch J, Hilger C, Bilò MB, Kler S, Schiener M, Dittmar G, Bernardin F, Lesur A, Ollert M, Schmidt-Weber CB, et al. Shedding Light on the Venom Proteomes of the Allergy-Relevant Hymenoptera Polistes dominula (European Paper Wasp) and Vespula spp. (Yellow Jacket). Toxins. 2020; 12(5):323. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12050323
Chicago/Turabian StyleGrosch, Johannes, Christiane Hilger, Maria Beatrice Bilò, Stephanie Kler, Maximilian Schiener, Gunnar Dittmar, François Bernardin, Antoine Lesur, Markus Ollert, Carsten B. Schmidt-Weber, and et al. 2020. "Shedding Light on the Venom Proteomes of the Allergy-Relevant Hymenoptera Polistes dominula (European Paper Wasp) and Vespula spp. (Yellow Jacket)" Toxins 12, no. 5: 323. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12050323