Behavioral, Physiological, Demographic and Ecological Impacts of Hematophagous and Endoparasitic Insects on an Arctic Ungulate


{kind=link}
{kind=link}
Abstract
:“The buck, who had been standing motionless at the opposite end of the gravel bar all this time, suddenly exploded into action, showing all the typical reactions to an attack by a nose or warble fly. He threw up his head, stamped his feet, twitched his hide, and then began to gallop along the beach, bucking and tossing as he ran. He suddenly stopped and stood stock still. Then he wheeled, galloped back in his tracks, stopped again, shook his head, wheeled once more and galloped full speed toward the band at the water’s edge. He literally burrowed his way into their midst where l lost him and presumably the nose fly did likewise.”—W. O. Pruitt [1]
1. Introduction
2. Impacts of Mosquitoes and Warble Flies on Caribou
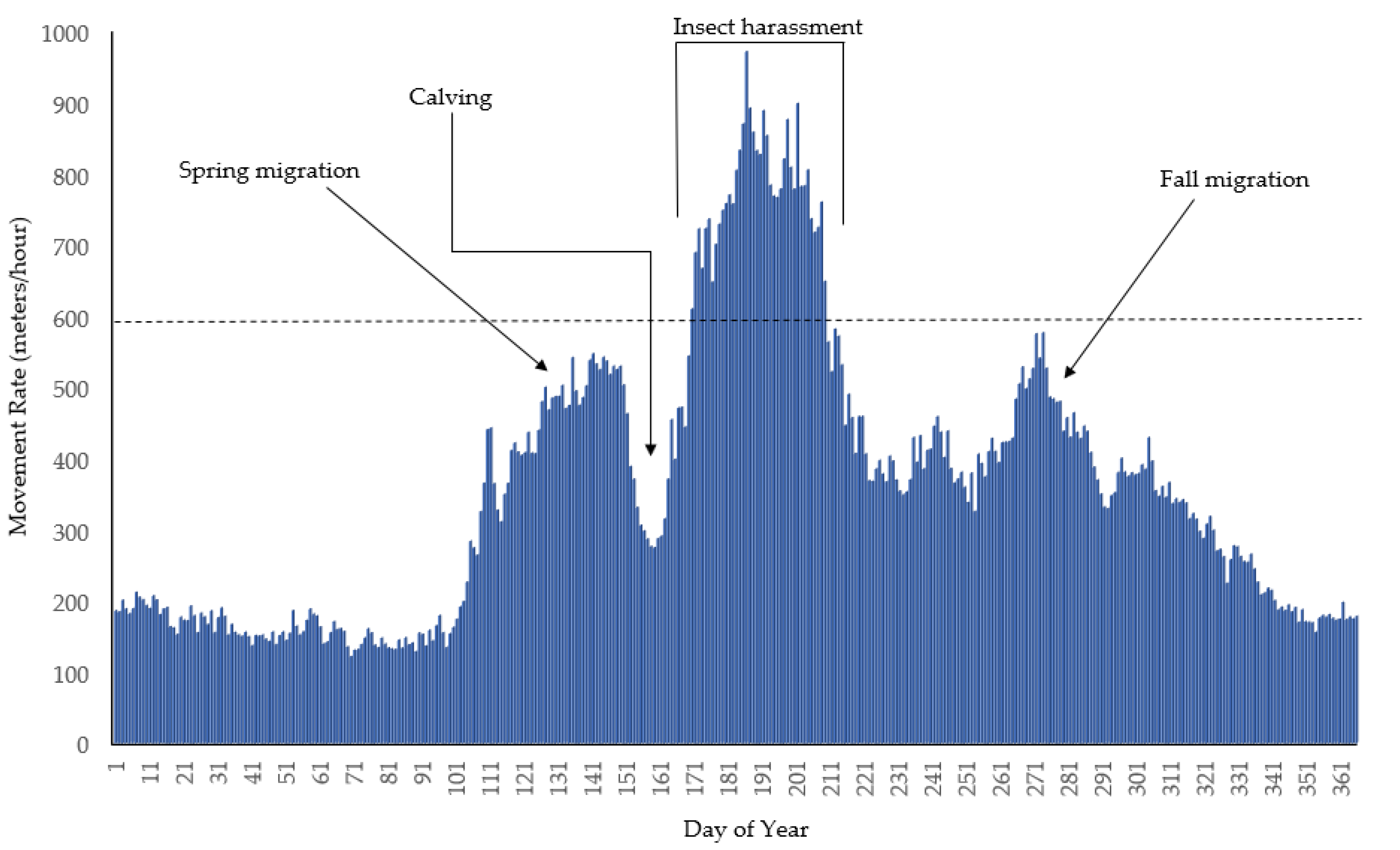
2.1. Behavioral Impacts
2.2. Physiological Impacts
2.3. Demographic Impacts
2.4. Ecological Impacts
3. Conclusions
4. Materials and Methods
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pruitt, W.O. Behavior of the barren-ground caribou. Biol. Pap. Univ. Alsk. 1960, 3, 1–44. [Google Scholar]
- Casewell, N.R.; Wuester, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evolut. 2013, 28, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Nelsen, D.R.; Nisani, Z.; Cooper, A.M.; Fox, G.A.; Gren, E.C.K.; Corbit, A.G.; Hayes, W.K. Poisons, toxungens, and venoms: Redefining and classifying toxic biological secretions and the organisms that employ them. Biol. Rev. 2014, 89, 450–465. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, C. Venomous: How Earth’s Deadliest Creatures Mastered Biochemistry; Scientific American: New York, NY, USA, 2016. [Google Scholar]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.A.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, J.M.C. Role of saliva in blood-feeding by arthropods. Ann. Rev. Entomol. 1987, 32, 463–478. [Google Scholar] [CrossRef] [PubMed]
- Champagne, D.E. Antihemostatic molecules from saliva of blood-feeding arthropods. Pathophysiol. Haemost. Thromb. 2005, 34, 221–227. [Google Scholar] [CrossRef]
- Colwell, D.D. Life cycle strategies. In The Oestrid Flies: Biology, Host-Parasite Relationships, Impact and Management; Colwell, D.D., Hall, M.J.R., Scholl, P.J., Eds.; CABI Publishing: Oxforshire, UK, 2006; p. 376. [Google Scholar]
- Åsbakk, K.; Kumpula, J.; Oksanen, A.; Laaksonen, S. Infestation by Hypoderma tarandi in reindeer calves from northern Finland—Prevalence and risk factors. Vet. Parasitol. 2014, 200, 172–178. [Google Scholar] [CrossRef]
- Culler, L.E.; Ayres, M.P.; Virginia, R.A. In a warmer Arctic, mosquitoes avoid increased mortality from predators by growing faster. Proc. R. Soc. B 2015, 282, 20151549. [Google Scholar] [CrossRef]
- Witter, L.A.; Johnson, C.J.; Croft, B.; Gunn, A.; Gillingham, M.P. Behavioural trade-offs in response to external stimuli: Time allocation of an Arctic ungulate during varying intensities of harassment by parasitic flies. J. Anim. Ecol. 2012, 81, 284–295. [Google Scholar] [CrossRef]
- Cameron, M.D.; Joly, K.; Weaver, L. Caribou: Did You Know? National Park Service: Fairbanks, AK, USA, 2017. Available online: https://www.nps.gov/gaar/learn/nature/upload/DidYouKnow_CaribouFinalMarch2017.pdf (accessed on 7 May 2020).
- Walsh, N.E.; Fancy, S.G.; McCabe, T.R.; Pank, F. Habitat use by the Porcupine Caribou Herd during predicted insect harassment. J. Wildl. Manag. 1992, 56, 465–473. [Google Scholar] [CrossRef]
- Witter, L.A.; Johnson, C.J.; Croft, B.; Gunn, A.; Poirier, L.M. Gauging climate change effects at local scales: Weather-based indices to monitor insect harassment in caribou. Ecol. Appl. 2012, 22, 1838–1851. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.C.; Kiliaan, H.P. Warble infestations in some Canadian caribou and their significance. Rangifer 1990, 3, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Cuyler, C.; White, R.R.; Lewis, K.; Soulliere, C.; Gunn, A.; Russell, D.E.; Daniel, C. Are warbles and bots related to reproductive status in West Greenland? Rangifer 2012, 12, 243–257. [Google Scholar] [CrossRef] [Green Version]
- Simard, A.; Kutz, S.; Ducrocq, J.; Beckmen, K.; Brodeur, V.; Croft, B.; Côté, S.D. Variation in the intensity and prevalence of macroparasites in migratory caribou: A quasi-circumpolar study. Can. J. Zool. 2016, 94, 607–617. [Google Scholar] [CrossRef]
- Nilssen, A.C.; Anderson, J.R. Flight capacity of the reindeer warble fly, Hypoderma tarandi (L.), and the reindeer nose bot fly, Cephenemyia trompe (Modeer) (Diptera:Oestridae). Can. J. Zool. 1995, 73, 1228–1238. [Google Scholar] [CrossRef]
- Helle, T. Abundance of warble fly (Oedemagena tarandi) larvae in semi-domestic reindeer (Rangifer tarandus) in Finland. Rep. Kevo Subarct. Res. Stn. 1980, 16, 1–6. [Google Scholar]
- Anderson, J.R.; Nilssen, A.C. Trapping oestrid parasites of reindeer: The response of Cephenemyia trompe and Hypoderma tarandi to baited traps. Med. Vet. Entomol. 1996, 10, 337–346. [Google Scholar] [CrossRef]
- Nilssen, A.C.; Haugerud, R.E. The timing and departure rate of larvae of the warble fly Hypoderma (= Oedemagena) tarandi (L.) and the nose bot fly Cephenemyia trompe (Modeer) (Diptera: Oestridae) from reindeer. Rangifer 1994, 14, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Mallory, C.D.; Boyce, M.S. Observed and predicted effects of climate change on Arctic caribou and reindeer. Environ. Rev. 2018, 26, 13–25. [Google Scholar] [CrossRef]
- Joly, K.; Klein, D.R.; Verbyla, D.L.; Rupp, T.S.; Chapin, F.S., III. Linkages between large-scale climate patterns and the dynamics of Arctic caribou populations. Ecography 2011, 34, 345–352. [Google Scholar] [CrossRef]
- Wolfe, R.J.; Walker, R.J. Subsistence economies in Alaska: Productivity, geography, and development impacts. Arctic Anthropo. 1987, 24, 56–81. [Google Scholar]
- Schwatka, F. A Summer in Alaska; J. W. Henry: St. Louis, MO, USA, 1894; p. 418. [Google Scholar]
- White, R.G.; Thompson, B.R.; Skoogland, T.; Person, S.J.; Holleman, D.F.; Luick, J.R. Ecology of caribou at Prudhoe Bay, Alaska. In Ecological Investigations of the Tundra Biome in the Prudhoe Bay Region, Alaska; Biological papers of the University of Alaska; Special Report No. 2; University of Alaska: Fairbanks, AK, USA, 1975; pp. 150–201. [Google Scholar]
- Russell, D.E.; Martell, A.M.; Nixon, W.A.C. Range ecology of the Porcupine Caribou Herd in Canada. Rangifer 1993, 8, 1–168. [Google Scholar] [CrossRef]
- Hagemoen, R.I.M.; Reimers, E. Reindeer summer activity pattern in relation to weather and insect harassment. J. Anim. Ecol. 2002, 71, 883–892. [Google Scholar] [CrossRef]
- Colman, J.E.; Pedersen, C.; Hjermann, D.Ø.; Holand, Ø.; Stein, R.; Moe, S.R.; Reimers, E. Do wild reindeer exhibit grazing compensation during insect harassment? J. Wildl. Manag. 2003, 67, 11–19. [Google Scholar] [CrossRef]
- Fancy, S.G.; Pank, L.F.; Whitten, K.R.; Regelin, W.L. Seasonal movements of caribou in Arctic Alaska as determined by satellite. Can. J. Zool. 1989, 67, 644–694. [Google Scholar] [CrossRef] [Green Version]
- Person, B.T.; Prichard, A.K.; Carroll, G.M.; Yokel, D.A.; Suydam, R.S.; George, J.C. Distribution and movements of the Teshekpuk Caribou Herd, 1990–2005: Prior to oil and gas development. Arctic 2007, 60, 238–250. [Google Scholar] [CrossRef] [Green Version]
- Prichard, A.K.; Yokel, D.A.; Rea, C.L.; Person, B.T.; Parrett, L.S. The effect of telemetry locations on movement-rate calculations in arctic caribou. Wildl. Soc. Bull. 2014, 38, 78–88. [Google Scholar] [CrossRef]
- Murphy, S.M.; Curatolo, J.A. Activity budgets and movement rates of caribou encountering pipelines, roads, and traffic in northern Alaska. Can. J. Zool. 1987, 65, 2483–2490. [Google Scholar] [CrossRef]
- Skoog, R.O. Ecology of the Caribou (Rangifer tarandus granti) in Alaska; University of California Berkeley: Berkeley, CA, USA, 1968. [Google Scholar]
- Skogland, T. Comparative social organization of wild reindeer in relation to food, mates and predator avoidance. In Life History Characteristics of Wild Reindeer (Rangifer tarandus L.) in Relation to their Food Resources; Ecological Effects and Behavioral Adaptations; Skogland, T., Ed.; Meddelelser fra norsk viltforskning: Berlin, Germany, 1985. [Google Scholar]
- Downes, C.M.; Theberge, J.B.; Smith, S.M. The influence of insects on the distribution, microhabitat choice, and behavior of the Burwash caribou herd. Can. J. Zool. 1986, 64, 622–629. [Google Scholar] [CrossRef]
- Wilson, R.R.; Prichard, A.K.; Parrett, L.S.; Person, B.T.; Carroll, G.M.; Smith, M.A.; Rea, C.L.; Yokel, D.A. Summer resource selection and identification of important habitat prior to industrial development for the Teshekpuk caribou herd in Northern Alaska. PLoS ONE 2012, 7, e48697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelsall, J.P. The Migratory Barren-Ground Caribou of North America; Queen’s Printer: Ottawa, ON, Canada, 1968. [Google Scholar]
- Fauchald, P.; Rødven, R.; Bårdsen, B.-J.; Langeland, K.; Tveraa, T.; Yoccoz, N.G.; Ims, R.A. Escaping parasitism in the selfish herd: Age, size and density dependent warble fly infestation in reindeer. Oikos 2007, 116, 491–499. [Google Scholar] [CrossRef]
- Hughes, J.; Albon, S.D.; Irvine, R.J.; Woodin, S. Is there a cost of parasites to caribou? Parasitology 2009, 136, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Folstad, I.; Nilssen, A.C.; Halvorsen, O.; Andersen, J. Parasite avoidance—The cause of post-calving migrations in Rangifer. Can. J. Zool. 1991, 69, 2423–2429. [Google Scholar] [CrossRef]
- Fancy, S.G.; White, R.G. Energy expenditures for locomotion by barren-ground caribou. Can. J. Zool. 1987, 65, 122–128. [Google Scholar] [CrossRef]
- Helle, T.; Tarvainen, L. Effects of insect harassment on weight gain and survival in reindeer calves. Rangifer 1984, 4, 24–27. [Google Scholar] [CrossRef] [Green Version]
- Gurarie, E.; Hebblewhite, M.; Joly, K.; Kelly, A.P.; Adamczewski, J.; Davidson, S.C.; Davison, T.; Gunn, A.; Suitor, M.J.; Fagan, W.F.; et al. Tactical departures and strategic arrivals: Divergent effects of climate and weather on caribou spring migrations. Ecosphere 2019, 10, e02971. [Google Scholar] [CrossRef] [Green Version]
- Folstad, I.; Nilssen, A.C.; Halvorsen, O.; Andersen, J. Why do male reindeer (Rangifer t. tarandus) have higher abundance of second and third instar larvae of Hypoderma tarandi than females? Oikos 1989, 55, 87–92. [Google Scholar] [CrossRef]
- Dale, B.W.; Adams, L.G.; Collins, W.B.; Joly, K.; Valkenburg, P.; Tobey, R. Stochastic and compensatory effects limit persistence of variation in body mass of young caribou. J. Mammal. 2008, 89, 1130–1135. [Google Scholar] [CrossRef]
- Weladji, R.B.; Holand, Ø.; Almøy, T. Use of climatic data to assess the effect of insect harassment on the autumn weight of reindeer (Rangifer tarandus) calves. J. Zool. 2003, 260, 79–85. [Google Scholar] [CrossRef]
- Ballesteros, M.; Bårdsen, B.-J.; Langeland, K.; Fauchald, P.; Stien, A.T.; Tveraa, T. The effect of warble flies on reindeer fitness: A parasite removal experiment. J. Zool. 2012, 287, 34–40. [Google Scholar] [CrossRef]
- Zhigunov, P. Reindeer Husbandry, 2nd ed.; Israel Program for Scientific Translations: Jerusalem, Israel, 2008; p. 343. [Google Scholar]
- Anderson, R.M.; May, R.M. Coevolution of hosts and parasites. Parasitology 1982, 85, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Cameron, R.D.; Smith, W.T.; Fancy, S.G.; Gerhart, K.L.; White, R.G. Calving success of female caribou in relation to body weight. Can. J. Zool. 1993, 71, 480–486. [Google Scholar] [CrossRef]
- Cameron, R.D.; Ver Hoef, J.M. Predicting parturition rate of caribou from autumn body mass. J. Wildl. Manag. 1994, 58, 674–679. [Google Scholar] [CrossRef]
- White, R.G. Foraging patterns and their multiplier effects on productivity of northern ungulates. Oikos 1983, 40, 377–384. [Google Scholar] [CrossRef]
- Albon, S.D.; Stien, A.; Irvine, R.J.; Langvatn, R.; Ropstad, E.; Halvorsen, O. The role of parasites in the dynamics of a reindeer population. Proc. R. Soc. Lond. B 2002, 269, 1625–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pachkowski, M.; Cote, S.D.; Festa-Bianchet, M. Spring-loaded reproduction: Effects of body condition and population size on fertility in migratory caribou (Rangifer tarandus). Can. J. Zool. 2013, 91, 473–479. [Google Scholar] [CrossRef] [Green Version]
- Henry, G.H.R.; Gunn, A. Recovery of tundra vegetation after overgrazing by caribou in arctic Canada. Arctic 1991, 44, 38–42. [Google Scholar] [CrossRef] [Green Version]
- Joly, K.; Jandt, R.R.; Meyers, C.R.; Cole, M.J. Changes in vegetative cover on Western Arctic Herd winter range from 1981 to 2005: Potential effects of grazing and climate change. Rangifer 2007, 27, 199–207. [Google Scholar] [CrossRef] [Green Version]
- te Beest, M.; Sitters, J.; Ménard, C.B.; Olofsson, J. Reindeer grazing increases summer albedo by reducing shrub abundance in Arctic tundra. Environ. Res. Lett. 2016, 11, 125013. [Google Scholar] [CrossRef] [Green Version]
- McKendrigk, J.D.; Batzli, G.O.; Everett, K.R.; Swanson, J.C. Some effects of mammalian herbivores and fertilization on tundra soils and vegetation. Arct. Alp. Res. 1980, 12, 565–578. [Google Scholar] [CrossRef]
- Joly, K.; Gurarie, E.; Sorum, M.S.; Kaczensky, P.; Cameron, M.D.; Jakes, A.F.; Borg, B.L.; Nandintsetseg, D.; Hopcraft, J.G.C.; Buuveibaatar, B.; et al. Longest terrestrial migrations and movements around the world. Sci. Rep. 2019, 9, 15333. [Google Scholar] [CrossRef] [PubMed]
- Adams, L.G.; Singer, F.J.; Dale, B.W. Caribou calf mortality in Denali National Park, Alaska. J. Wildl. Manag. 1995, 59, 584–594. [Google Scholar] [CrossRef]
- Magoun, A.J.; Laird, C.R.; Keech, M.A.; Valkenburg, P.; Parrett, L.S.; Robards, M.D. Predation on caribou (Rangifer tarandus) by wolverines (Gulo gulo) after long pursuits. Can. Field-Nat. 2018, 132, 382–385. [Google Scholar] [CrossRef]
- Fang, J. A world without mosquitoes. Nature 2010, 466, 432–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brazda, A.R. Elk migration patterns, and some of the factors affecting movements in the Gallatin River drainage, Montana. J. Wildl. Manag. 1953, 17, 9–23. [Google Scholar] [CrossRef] [Green Version]
- Tempelis, C.H. Host-feeding patterns of mosquitoes, with a review of advances in analysis of blood meals by serology. J. Med. Entomol. 1975, 11, 635–653. [Google Scholar] [CrossRef]
- Kulasekera, V.L.; Kramer, L.; Nasci, R.S.; Mostashari, F.; Cherry, B.; Trock, S.C.; Glaser, C.; Miller, J.R. West Nile virus infection in mosquitoes, birds, horses, and humans, Staten Island, New York, 2000. Emerg. Infect. Dis. 2001, 7, 722–725. [Google Scholar] [CrossRef]
- Rätti, O.; Ojanen, U.; Helle, P. Increasing group size dilutes black fly attack rate in Black Grouse. Ornis Fennica 2006, 83, 86–90. [Google Scholar]
- Cohen, J.; Screen, J.A.; Furtado, J.C.; Barlow, M.; Whittleston, D.; Coumou, D.; Francis, J.; Dethloff, K.; Entekhabi, D.; Overland, J.; et al. Recent Arctic amplification and extreme mid-latitude weather. Nature Geosci 2014, 7, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Samuel, W.M. Grooming by moose (Alces alces) infested with the winter tick, Dermacentor albipictus (Acari): A mechanism for premature loss of winter hair. Can. J. Zool. 1991, 69, 1255–1260. [Google Scholar] [CrossRef]
- Cabezas-Cruz, A.; Valdes, J.J. Are ticks venomous animals? Front. Zool. 2014, 11, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witter, L.; Johnson, C.; Croft, B. Weather-Based Indices of Parasitic Fly Activity and Abundance for the Bathurst Caribou Post-Calving and Summer Range: Users Guide; Manuscript Report 246; University of North British Columbia, Prince George, British Columbia: Prince George, BC, Canada, 2014; p. 37. [Google Scholar]
- Joly, K.; Cameron, M.D. Caribou Vital Sign Annual Report for the Arctic Network Inventory and Monitoring Program: September 2018-August 2019; Natural Resource Report NPS/ARCN/NRR—2019/2041; National Park Service: Fort Collins, CO, USA, 2019; p. 29.
- Joly, K. The effects of sampling regime on the analysis of movements of overwintering female caribou in east-central Alaska. Rangifer 2005, 25, 67–74. [Google Scholar] [CrossRef] [Green Version]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joly, K.; Couriot, O.; Cameron, M.D.; Gurarie, E. Behavioral, Physiological, Demographic and Ecological Impacts of Hematophagous and Endoparasitic Insects on an Arctic Ungulate. Toxins 2020, 12, 334. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12050334
Joly K, Couriot O, Cameron MD, Gurarie E. Behavioral, Physiological, Demographic and Ecological Impacts of Hematophagous and Endoparasitic Insects on an Arctic Ungulate. Toxins. 2020; 12(5):334. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12050334
Chicago/Turabian StyleJoly, Kyle, Ophélie Couriot, Matthew D. Cameron, and Eliezer Gurarie. 2020. "Behavioral, Physiological, Demographic and Ecological Impacts of Hematophagous and Endoparasitic Insects on an Arctic Ungulate" Toxins 12, no. 5: 334. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12050334