The Virulence Potential of Livestock-Associated Methicillin-Resistant Staphylococcus aureus Cultured from the Airways of Cystic Fibrosis Patients


 and
and
Abstract
:1. Introduction
2. Results
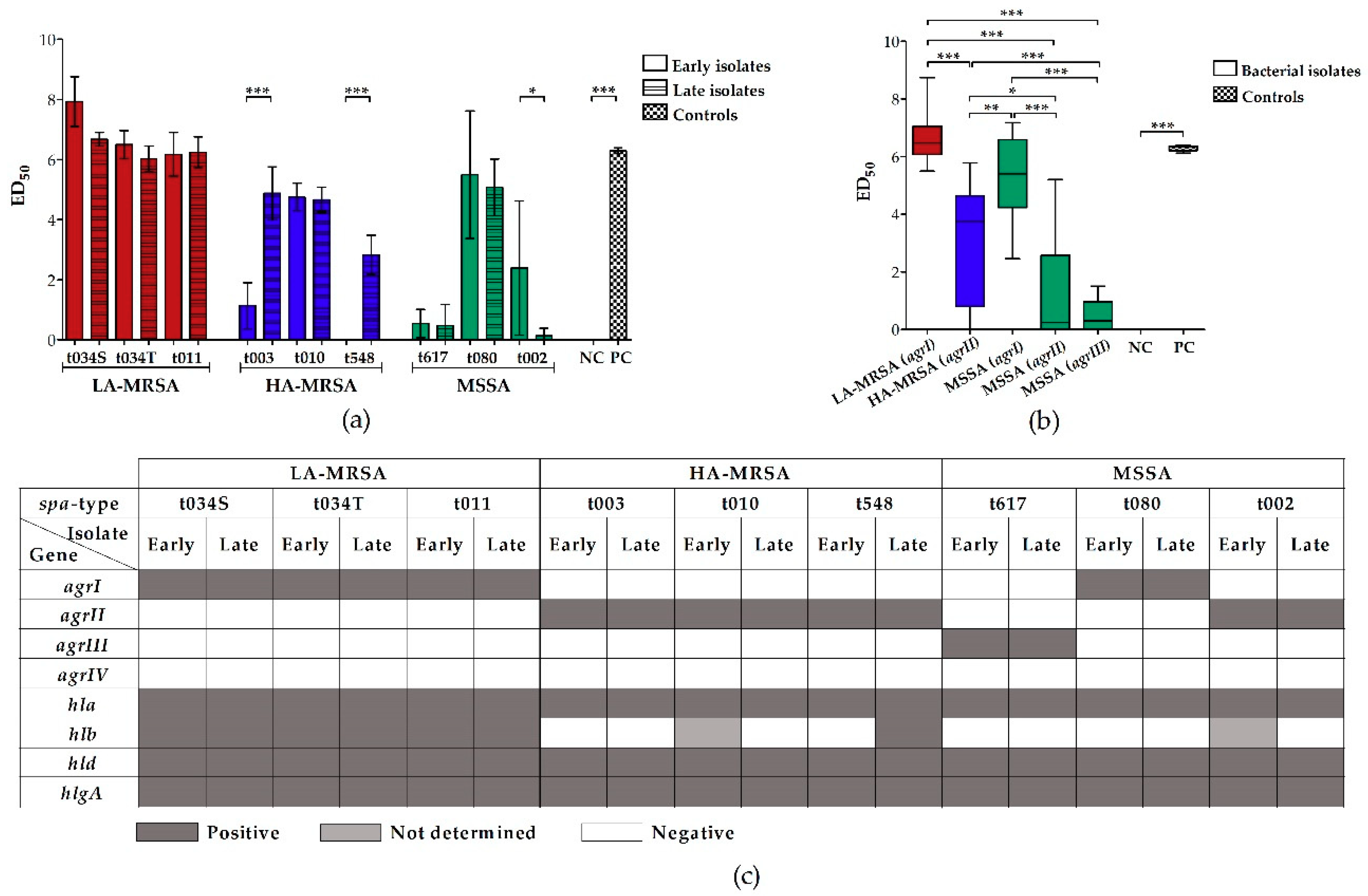
2.1. LA-MRSA Are Strongly Hemolytic
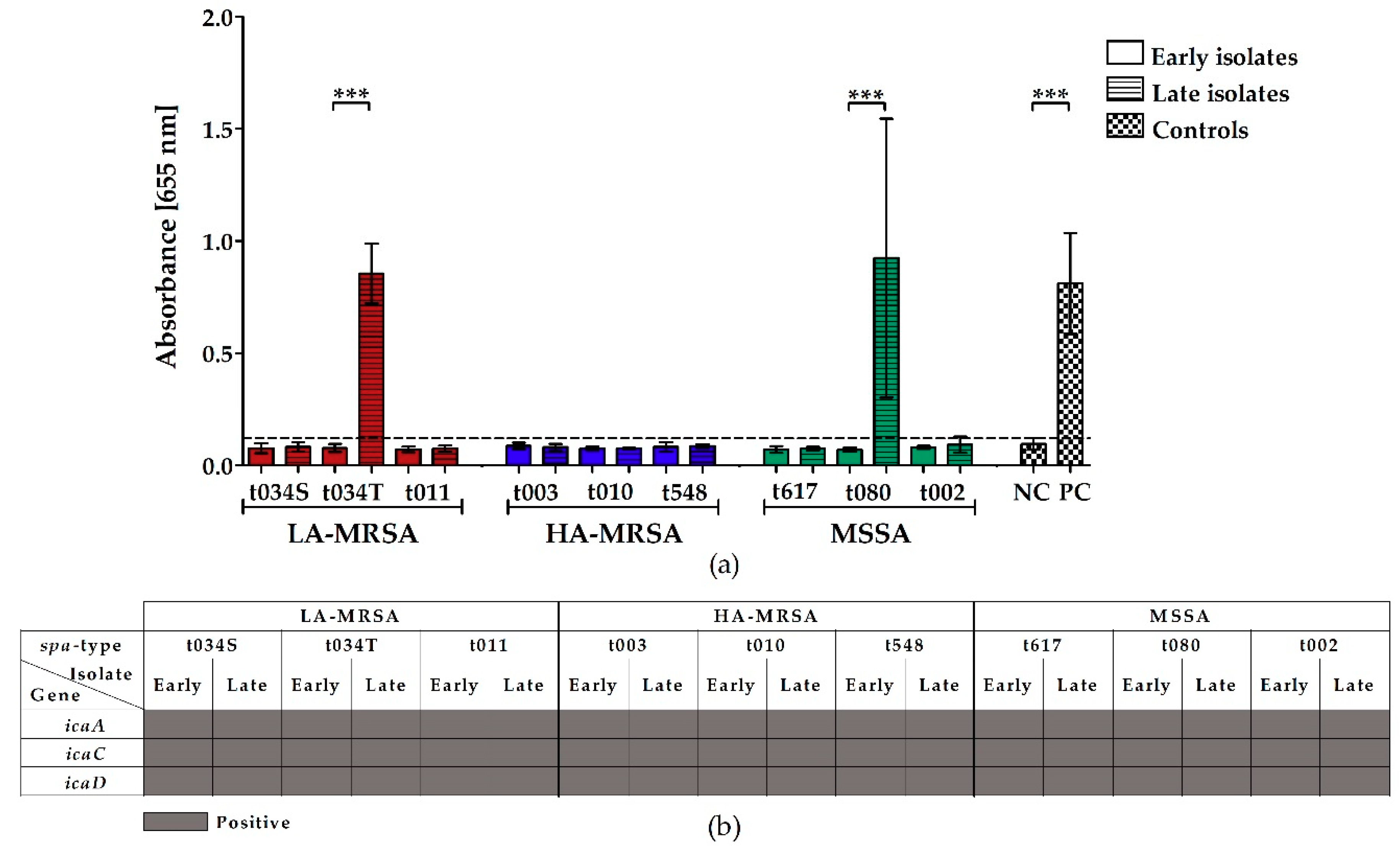
2.2. LA-MRSA Are Weak Biofilm Formers
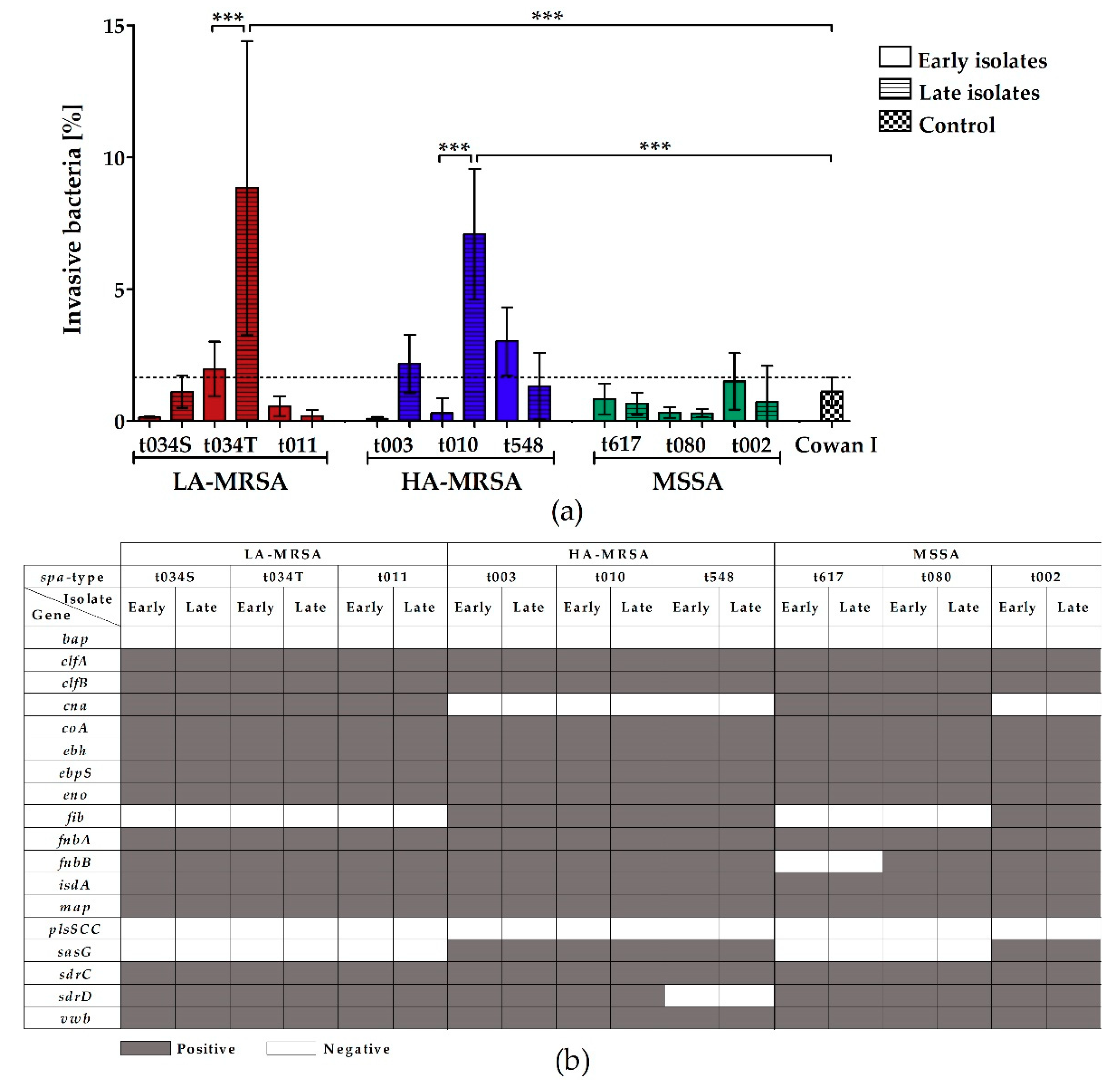
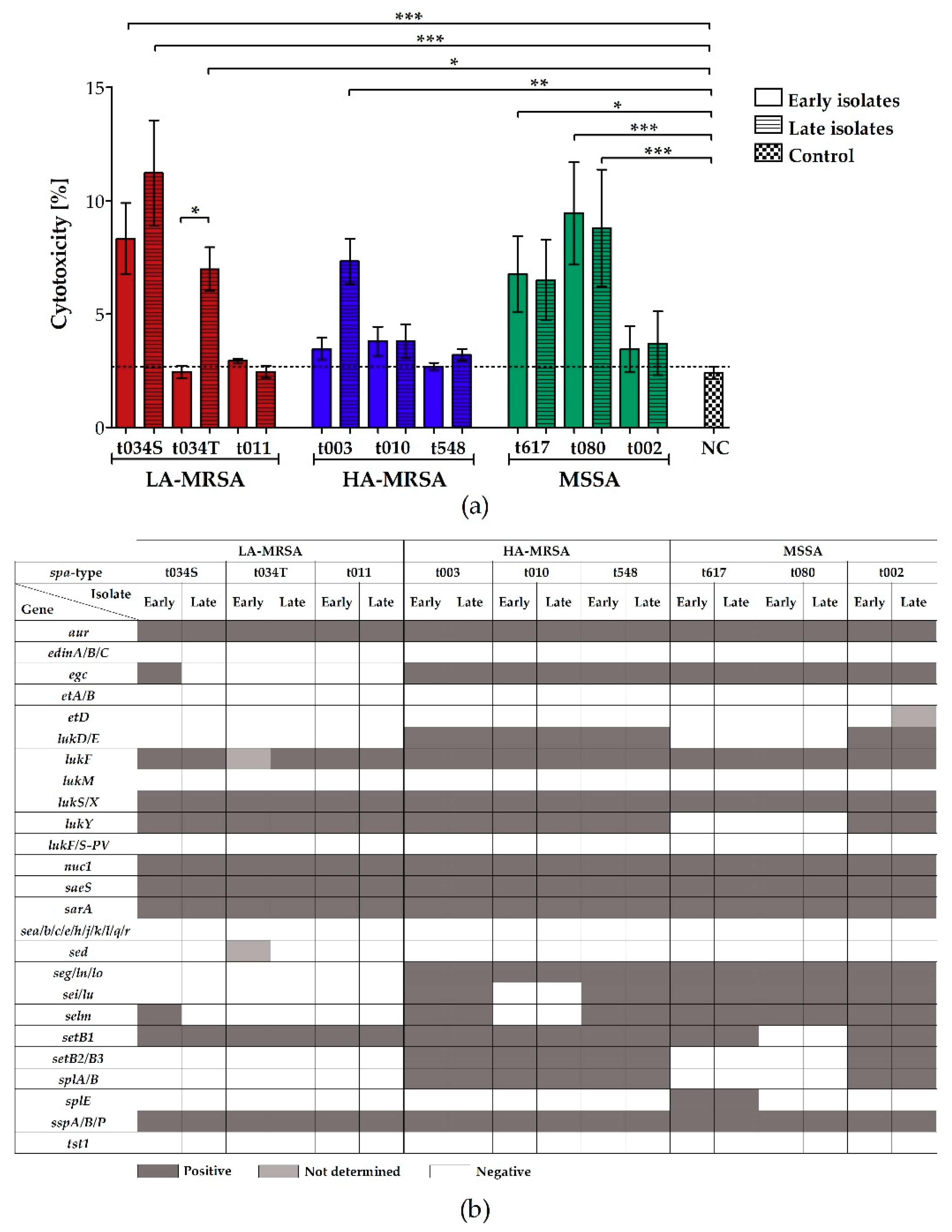
2.3. LA-MRSA Invade Airway Epithelial Cells and Are Cytotoxic
2.4. LA-MRSA Are Tetracycline-Resistant
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Bacterial Strains and Growth Conditions
4.3. spa-Typing
4.4. Microarray Analysis
4.5. Hemolysis Assay
4.5.1. Macroscopic Evaluation
4.5.2. Photometrical Quantification
4.6. Biofilm Assay
4.7. Cell Culture Infection Model
4.8. Cytotoxicity Assay
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cystic Fibrosis Foundation Patient Registry. 2017 Annual Data Report; American Cystic Fibrosis Foundation: Bethesda, MD, USA, 2018. [Google Scholar]
- Schwerdt, M.; Neumann, C.; Schwartbeck, B.; Kampmeier, S.; Herzog, S.; Görlich, D.; Dübbers, A.; Große-Onnebrink, J.; Kessler, C.; Küster, P.; et al. Staphylococcus aureus in the Airways of Cystic Fibrosis Patients—A Retrospective Long-Term Study. Int. J. Med. Microbiol. 2018, 308, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Cutting, G.R. Cystic Fibrosis Genetics: From Molecular Understanding to Clinical Application. Nat. Rev. Genet. 2015, 16, 45–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Döring, G.; Gulbins, E. Cystic Fibrosis and Innate Immunity: How Chloride Channel Mutations Provoke Lung Disease. Cell. Microbiol. 2009, 11, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Junge, S.; Görlich, D.; den Reijer, M.; Wiedemann, B.; Tümmler, B.; Ellemunter, H.; Dübbers, A.; Küster, P.; Ballmann, M.; Koerner-Rettberg, C.; et al. Factors Associated with Worse Lung Function in Cystic Fibrosis Patients with Persistent Staphylococcus aureus. PLoS ONE 2016, 11, e0166220. [Google Scholar] [CrossRef] [PubMed]
- Dasenbrook, E.C.; Checkley, W.; Merlo, C.A.; Konstan, M.W.; Lechtzin, N.; Boyle, M.P. Association between Respiratory Tract Methicillin-Resistant Staphylococcus Aureus and Survival in Cystic Fibrosis. JAMA 2010, 303, 2386–2392. [Google Scholar] [CrossRef] [Green Version]
- Döring, G.; Hoiby, N.; Assael, B.; Ballmann, M.; Bush, A.; Button, B.; Clement, A.; Conway, S.; Dankert-Roelse, J.; de Baets, F.; et al. Early Intervention and Prevention of Lung Disease in Cystic Fibrosis: A European Consensus. J. Cyst. Fibros. 2004, 3, 67–91. [Google Scholar] [CrossRef] [Green Version]
- Chambers, H.F.; DeLeo, F.R. Waves of Resistance: Staphylococcus Aureus in the Antibiotic Era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef]
- Schaumburg, F.; Köck, R.; Mellmann, A.; Richter, L.; Hasenberg, F.; Kriegeskorte, A.; Friedrich, A.W.; Gatermann, S.; Peters, G.; Von Eiff, C.; et al. Population Dynamics among Methicillin-Resistant Staphylococcus aureus Isolates in Germany during a 6-Year Period. J. Clin. Microbiol. 2012, 50, 3186–3192. [Google Scholar] [CrossRef] [Green Version]
- Price, L.B.; Stegger, M.; Hasman, H.; Aziz, M.; Larsen, J.; Andersen, P.S.; Pearson, T.; Waters, A.E.; Foster, J.T.; Schupp, J.; et al. Staphylococcus aureus CC398: Host Adaptation and Emergence of Methicillin Resistance in Livestock. MBio 2012, 3, e00305–e00311. [Google Scholar] [CrossRef] [Green Version]
- Köck, R.; Schaumburg, F.; Mellmann, A.; Köksal, M.; Jurke, A.; Becker, K.; Friedrich, A.W. Livestock-Associated Methicillin-Resistant Staphylococcus aureus (MRSA) as Causes of Human Infection and Colonization in Germany. PLoS ONE 2013, 8, e55040. [Google Scholar] [CrossRef] [Green Version]
- Becker, K.; Ballhausen, B.; Kahl, B.C.; Köck, R. The Clinical Impact of Livestock-Associated Methicillin-Resistant Staphylococcus Aureus of the Clonal Complex 398 for Humans. Vet. Microbiol. 2017, 200, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Clarke, S.R.; Foster, S.J. Surface Adhesins of Staphylococcus aureus. Adv. Microb. Physiol. 2006, 51, 187–224. [Google Scholar] [PubMed]
- Bien, J.; Sokolova, O.; Bozko, P. Characterization of Virulence Factors of Staphylococcus aureus: Novel Function of Known Virulence Factors That Are Implicated in Activation of Airway Epithelial Proinflammatory Response. J. Pathog. 2011, 2011, 601905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Recsei, P.; Kreiswirth, B.; O’Reilly, M.; Schlievert, P.; Gruss, A.; Novick, R.P. Regulation of Exoprotein Gene Expression in Staphylococcus aureus by Agr. MGG Mol. Gen. Genet. 1986, 202, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Vandenesch, F.; Lina, G.; Henry, T. Staphylococcus aureus Hemolysins, Bi-Component Leukocidins, and Cytolytic Peptides: A Redundant Arsenal of Membrane-Damaging Virulence Factors? Front. Cell. Infect. Microbiol. 2012, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- Van Wamel, W.J.B.; Rooijakkers, S.H.M.; Ruyken, M.; Van Kessel, K.P.M.; Van Strijp, J.A.G. The Innate Immune Modulators Staphylococcal Complement Inhibitor and Chemotaxis Inhibitory Protein of Staphylococcus aureus Are Located on β-Hemolysin-Converting Bacteriophages. J. Bacteriol. 2006, 188, 1310–1315. [Google Scholar] [CrossRef] [Green Version]
- Otto, M. Staphylococcal Infections: Mechanisms of Biofilm Maturation and Detachment as Critical Determinants of Pathogenicity. Annu. Rev. Med. 2013, 64, 175–188. [Google Scholar] [CrossRef]
- Schwartbeck, B.; Birtel, J.; Treffon, J.; Langhanki, L.; Mellmann, A.; Kale, D.; Kahl, J.; Hirschhausen, N.; Neumann, C.; Lee, J.C.; et al. Dynamic in Vivo Mutations within the Ica Operon during Persistence of Staphylococcus aureus in the Airways of Cystic Fibrosis Patients. PLoS Pathog. 2016, 12, e1006024. [Google Scholar] [CrossRef]
- Kahl, B.C.; Goulian, M.; Van Wamel, W.; Herrmann, M.; Simon, S.M.; Kaplan, G.; Peters, G.; Cheung, A.L. Staphylococcus aureus RN6390 Replicates and Induces Apoptosis in a Pulmonary Epithelial Cell Line. Infect. Immun. 2000, 68, 5385–5392. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Coureuil, M.; Ramond, E.; Euphrasie, D.; Dupuis, M.; Tros, F.; Meyer, J.; Nemazanyy, I.; Chhuon, C.; Guerrera, I.C.; et al. Chronic Staphylococcus aureus Lung Infection Correlates with Proteogenomic and Metabolic Adaptations Leading to an Increased Intracellular Persistence. Clin. Infect. Dis. 2019, 69, 1937–1945. [Google Scholar] [CrossRef]
- Lima, D.F.; Cohen, R.W.F.; Rocha, G.A.; Albano, R.M.; Marques, E.A.; Leão, R.S. Genomic Information on Multidrug-Resistant Livestock-Associated Methicillin-Resistant Staphylococcus aureus ST398 Isolated from a Brazilian Patient with Cystic Fibrosis. Mem. Inst. Oswaldo Cruz 2017, 112, 79–80. [Google Scholar] [CrossRef] [PubMed]
- Garbacz, K.; Piechowicz, L.; Podkowik, M.; Mroczkowska, A.; Empel, J.; Bania, J. Emergence and Spread of Worldwide Staphylococcus aureus Clones among Cystic Fibrosis Patients. Infect. Drug Resist. 2018, 11, 247–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodémont, M.; Argudín, M.A.; Willekens, J.; Vanderhelst, E.; Pierard, D.; Miendje Deyi, V.Y.; Hanssens, L.; Franckx, H.; Schelstraete, P.; Leroux-Roels, I.; et al. Emergence of Livestock-Associated MRSA Isolated from Cystic Fibrosis Patients: Result of a Belgian National Survey. J. Cyst. Fibros. 2019, 18, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Ballhausen, B.; Kriegeskorte, A.; Schleimer, N.; Peters, G.; Becker, K. The MecA Homolog MecC Confers Resistance against β-Lactams in Staphylococcus aureus Irrespective of the Genetic Strain Background. Antimicrob. Agents Chemother. 2014, 58, 3791–3798. [Google Scholar] [CrossRef] [Green Version]
- Bernheimmer, A.W.; Avigad, L.S.; Kim, K.S. Staphylococcal Sphingomyelinase (Beta-Hemolysin). Ann. N. Y. Acad. Sci. 1974, 236, 292–306. [Google Scholar] [CrossRef]
- Chan, F.K.M.; Moriwaki, K.; De Rosa, M.J. Detection of Necrosis by Release of Lactate Dehydrogenase Activity. Methods Mol. Biol. 2013, 979, 65–70. [Google Scholar]
- Van Alen, S.; Ballhausen, B.; Peters, G.; Friedrich, A.W.; Mellmann, A.; Köck, R.; Becker, K. In the Centre of an Epidemic: Fifteen Years of LA-MRSA CC398 at the University Hospital Münster. Vet. Microbiol. 2017, 200, 19–24. [Google Scholar] [CrossRef]
- Köck, R.; Harlizius, J.; Bressan, N.; Laerberg, R.; Wieler, L.H.; Witte, W.; Deurenberg, R.H.; Voss, A.; Becker, K.; Friedrich, A.W. Prevalence and Molecular Characteristics of Methicillin-Resistant Staphylococcus aureus (MRSA) among Pigs on German Farms and Import of Livestock-Related MRSA into Hospitals. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 1375–1382. [Google Scholar] [CrossRef] [Green Version]
- Mairpady Shambat, S.; Haggar, A.; Vandenesch, F.; Lina, G.; Van Wamel, W.J.B.; Arakere, G.; Svensson, M.; Norrby-Teglund, A. Levels of Alpha-Toxin Correlate with Distinct Phenotypic Response Profiles of Blood Mononuclear Cells and with Agr Background of Community-Associated Staphylococcus aureus Isolates. PLoS ONE 2014, 9, e106107. [Google Scholar] [CrossRef] [Green Version]
- Salgado-Pabón, W.; Herrera, A.; Vu, B.G.; Stach, C.S.; Merriman, J.A.; Spaulding, A.R.; Schlievert, P.M. Staphylococcus aureus β-Toxin Production Is Common in Strains with the β-Toxin Gene Inactivated by Bacteriophage. J. Infect. Dis. 2014, 210, 784–792. [Google Scholar] [CrossRef] [Green Version]
- Van Alen, S.; Ballhausen, B.; Kaspar, U.; Köck, R.; Becker, K. Prevalence and Genomic Structure of Bacteriophage Phi3 in Human-Derived Livestock-Associated Methicillin-Resistant Staphylococcus aureus Isolates from 2000 to 2015. J. Clin. Microbiol. 2018, 56, e00140-18. [Google Scholar] [CrossRef] [Green Version]
- Kaspar, U.; von Lützau, A.; Schlattmann, A.; Roesler, U.; Köck, R.; Becker, K. Zoonotic Multidrug-Resistant Microorganisms among Small Companion Animals in Germany. PLoS ONE 2018, 13, e0208364. [Google Scholar] [CrossRef] [Green Version]
- Köck, R.; Siam, K.; Al-Malat, S.; Christmann, J.; Schaumburg, F.; Becker, K.; Friedrich, A.W. Characteristics of Hospital Patients Colonized with Livestock-Associated Meticillin-Resistant Staphylococcus aureus (MRSA) CC398 versus Other MRSA Clones. J. Hosp. Infect. 2011, 79, 292–296. [Google Scholar] [CrossRef]
- Kinross, P.; Petersen, A.; Skov, R.; Van Hauwermeiren, E.; Pantosti, A.; Laurent, F.; Voss, A.; Kluytmans, J.; Struelens, M.; Heuer, O.; et al. Livestock-Associated Meticillin-Resistant Staphylococcus aureus (MRSA) among Human MRSA Isolates, European Union/European Economic Area Countries, 2013. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speziale, P.; Pietrocola, G.; Foster, T.J.; Geoghegan, J.A. Protein-Based Biofilm Matrices in Staphylococci. Front. Cell. Infect. Microbiol. 2014, 4, 171. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Barat, L.; Ben-Aicha, S.; Motos, A.; Vila, J.; Marco, F.; Rigol, M.; Muñoz, L.; Li Bassi, G.; Ferrer, M.; Torres, A. Assessment of in vivo versus in vitro Biofilm Formation of Clinical Methicillin-Resistant Staphylococcus aureus Isolates from Endotracheal Tubes. Sci. Rep. 2018, 8, 11906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballhausen, B.; Jung, P.; Kriegeskorte, A.; Makgotlho, P.E.; Ruffing, U.; von Müller, L.; Köck, R.; Peters, G.; Herrmann, M.; Ziebuhr, W.; et al. LA-MRSA CC398 Differ from Classical Community Acquired-MRSA and Hospital Acquired-MRSA Lineages: Functional Analysis of Infection and Colonization Processes. Int. J. Med. Microbiol. 2014, 304, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Tuchscherr, L.; Bischoff, M.; Lattar, S.M.; Noto Llana, M.; Pförtner, H.; Niemann, S.; Geraci, J.; Van de Vyver, H.; Fraunholz, M.J.; Cheung, A.L.; et al. Sigma Factor SigB Is Crucial to Mediate Staphylococcus aureus Adaptation during Chronic Infections. PLoS Pathog. 2015, 11, e1004870. [Google Scholar] [CrossRef]
- Treffon, J.; Chaves-Moreno, D.; Niemann, S.; Pieper, D.H.; Vogl, T.; Roth, J.; Kahl, B.C. Importance of Superoxide Dismutase A and M for Protection of Staphylococcus aureus in the Oxidative Stressful Environment of Cystic Fibrosis Airways. Cell. Microbiol. 2020, e13158. [Google Scholar] [CrossRef] [Green Version]
- Battistoni, A.; Pacello, F.; Folcarelli, S.; Ajello, M.; Donnarumma, G.; Greco, R.; Ammendolia, M.G.; Touati, D.; Rotilio, G.; Valenti, P. Increased Expression of Periplasmic Cu, Zn Superoxide Dismutase Enhances Survival of Escherichia coli Invasive Strains within Nonphagocytic Cells. Infect. Immun. 2000, 68, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Gunaratnam, G.; Tuchscherr, L.; Elhawy, M.I.; Bertram, R.; Eisenbeis, J.; Spengler, C.; Tschernig, T.; Löffler, B.; Somerville, G.A.; Jacobs, K.; et al. ClpC Affects the Intracellular Survival Capacity of Staphylococcus aureus in Non-Professional Phagocytic Cells. Sci. Rep. 2019, 9, 16267. [Google Scholar] [CrossRef] [PubMed]
- Lekagul, A.; Tangcharoensathien, V.; Yeung, S. Patterns of Antibiotic Use in Global Pig Production: A Systematic Review. Vet. Anim. Sci. 2019, 7, 100058. [Google Scholar] [CrossRef]
- Chmiel, J.F.; Aksamit, T.R.; Chotirmall, S.H.; Dasenbrook, E.C.; Elborn, J.S.; LiPuma, J.J.; Ranganathan, S.C.; Waters, V.J.; Ratjen, F.A. Antibiotic Management of Lung Infections in Cystic Fibrosis: I. The Microbiome, Methicillin-Resistant Staphylococcus aureus, Gram-Negative Bacteria, and Multiple Infections. Ann. Am. Thorac. Soc. 2014, 11, 1120–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschhausen, N.; Block, D.; Bianconi, I.; Bragonzi, A.; Birtel, J.; Lee, J.C.; Dübbers, A.; Küster, P.; Kahl, J.; Peters, G.; et al. Extended Staphylococcus aureus Persistence in Cystic Fibrosis Is Associated with Bacterial Adaptation. Int. J. Med. Microbiol. 2013, 303, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Löfblom, J.; Rosenstein, R.; Nguyen, M.T.; Ståhl, S.; Götz, F. Staphylococcus carnosus: From Starter Culture to Protein Engineering Platform. Appl. Microbiol. Biotechnol. 2017, 101, 8293–8307. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Investigated Bacteria | 37 °C Incubation | 4 °C Incubation | |||
|---|---|---|---|---|---|
| Group | spa-Type | Isolate | Clear Hemolytic Zone | Dark, Diffuse Hemolytic Zone | Clear Hemolytic Zone |
| LA-MRSA | t034S | Early | + | ++++ | ++++ |
| Late | + | ++++ | ++++ | ||
| t034T | Early | + | ++++ | ++++ | |
| Late | + | ++++ | ++++ | ||
| t011 | Early | + | ++++ | ++++ | |
| Late | + | ++++ | ++++ | ||
| HA-MRSA | t003 | Early | + | - | ++ |
| Late | ++ | - | ++ | ||
| t010 | Early | ++ | - | +++ | |
| Late | ++ | - | +++ | ||
| t548 | Early | - | - | - | |
| Late | - | ++++ | ++++ | ||
| MSSA | t617 | Early | + | - | + |
| Late | - | - | + | ||
| t080 | Early | ++ | - | +++ | |
| Late | + | - | ++ | ||
| t002 | Early | ++ | - | +++ | |
| Late | ++ | - | +++ | ||
| LA-MRSA | HA-MRSA | MSSA | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| spa-type | t034S | t034T | t011 | t003 | t010 | t548 | t617 | t080 | t002 | |||||||||
| Isolate | Early | Late | Early | Late | Early | Late | Early | Late | Early | Late | Early | Late | Early | Late | Early | Late | Early | Late |
| Gene | ||||||||||||||||||
| aacA-aphD | ||||||||||||||||||
| aadD | ||||||||||||||||||
| aphA3 | ||||||||||||||||||
| blaI/R/Z | ||||||||||||||||||
| cat | ||||||||||||||||||
| cfr | ||||||||||||||||||
| dfrS1 | ||||||||||||||||||
| ermA | ||||||||||||||||||
| ermB | ||||||||||||||||||
| ermC | ||||||||||||||||||
| fexA | ||||||||||||||||||
| fosB | ||||||||||||||||||
| inuA | ||||||||||||||||||
| mecA | ||||||||||||||||||
| mefA | ||||||||||||||||||
| mphC | ||||||||||||||||||
| msrA | ||||||||||||||||||
| mupA | ||||||||||||||||||
| tetK | ||||||||||||||||||
| tetM | ||||||||||||||||||
| vanA/B/Z | ||||||||||||||||||
| vatA/B | ||||||||||||||||||
| vgaA | ||||||||||||||||||
| Strain | Patient | Isolate | MLST | spa-type | spa-type Repeats | Origin | Year of Isolation |
|---|---|---|---|---|---|---|---|
| LA-MRSA | 1 | Early | CC398 | t034 | 08-16-02-25-02-25-34-24-25 | Sputum | 2014 |
| Late | 2017 | ||||||
| 2 | Early | CC398 | t034 | 08-16-02-25-02-25-34-24-25 | Throat | 2002 | |
| Late | 2010 | ||||||
| 3 | Early | CC398 | t011 | 08-16-02-25-34-24-25 | Sputum | 2012 | |
| Late | 2016 | ||||||
| HA-MRSA | 4 | Early | CC5 | t003 | 26-17-20-17-12-17-17-16 | Sputum | 2010 |
| Late | 2016 | ||||||
| 5 | Early | CC5 | t010 | 26-17-34-17-20-17-12-17-16 | Sputum | 2013 | |
| Late | 2017 | ||||||
| 6 | Early | CC5 | t548 | 26-23-17-34-17-20-17-12-16 | Throat | 2008 | |
| Late | 2014 | ||||||
| MSSA | 7 | Early | CC30 | t617 | 15-21-16-02-24-24 | Throat | 2001 |
| Late | Sputum | 2010 | |||||
| 8 | Early | CC45 | t080 | 09-02-16-34-42-17-16-34 | Throat | 1995 | |
| Late | 2008 | ||||||
| 9 | Early | CC5 | t002 | 26-23-17-34-17-20-17-12-17-16 | Sputum | 1994 | |
| Late | 1997 |
| Bacterial species | Strain | Application | Reference |
|---|---|---|---|
| Staphylococcus aureus | Cowan I | Invasion assay (PC) | ATCC 12598 |
| Staphylococcus carnosus | TM300 | Biofilm assay (NC) | [46] |
| Staphylococcus epidermidis | RP62A | Biofilm assay (PC) | ATCC 35984 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Treffon, J.; Fotiadis, S.A.; van Alen, S.; Becker, K.; Kahl, B.C. The Virulence Potential of Livestock-Associated Methicillin-Resistant Staphylococcus aureus Cultured from the Airways of Cystic Fibrosis Patients. Toxins 2020, 12, 360. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12060360
Treffon J, Fotiadis SA, van Alen S, Becker K, Kahl BC. The Virulence Potential of Livestock-Associated Methicillin-Resistant Staphylococcus aureus Cultured from the Airways of Cystic Fibrosis Patients. Toxins. 2020; 12(6):360. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12060360
Chicago/Turabian StyleTreffon, Janina, Sarah Ann Fotiadis, Sarah van Alen, Karsten Becker, and Barbara C. Kahl. 2020. "The Virulence Potential of Livestock-Associated Methicillin-Resistant Staphylococcus aureus Cultured from the Airways of Cystic Fibrosis Patients" Toxins 12, no. 6: 360. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12060360