Exoenzyme Y Contributes to End-Organ Dysfunction Caused by Pseudomonas aeruginosa Pneumonia in Critically Ill Patients: An Exploratory Study

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
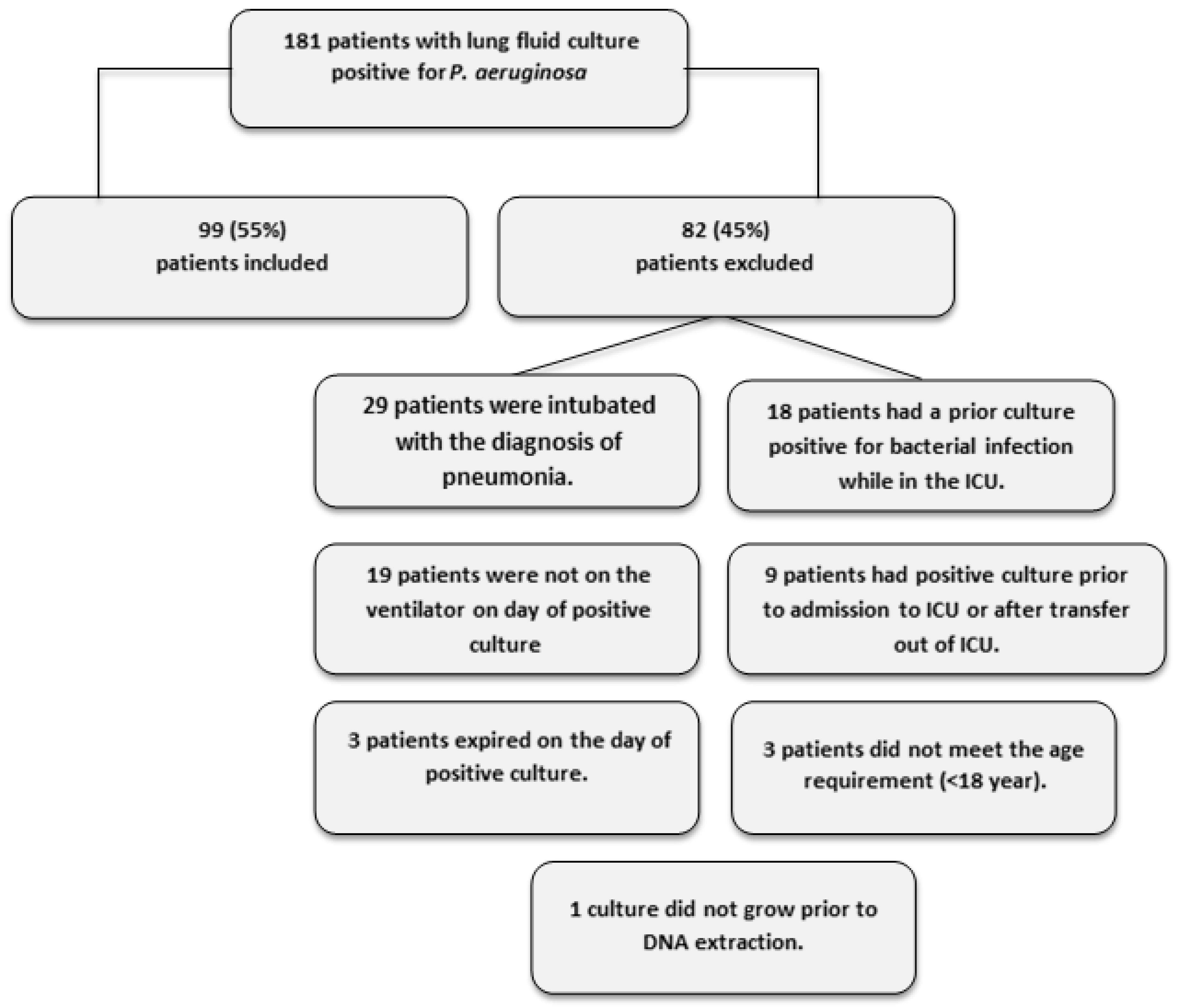
2.1. Patient Recruitment and Demographics
2.2. ExoY is More Prevalent than ExoU in Patients with P. aeruginosa Pneumonia
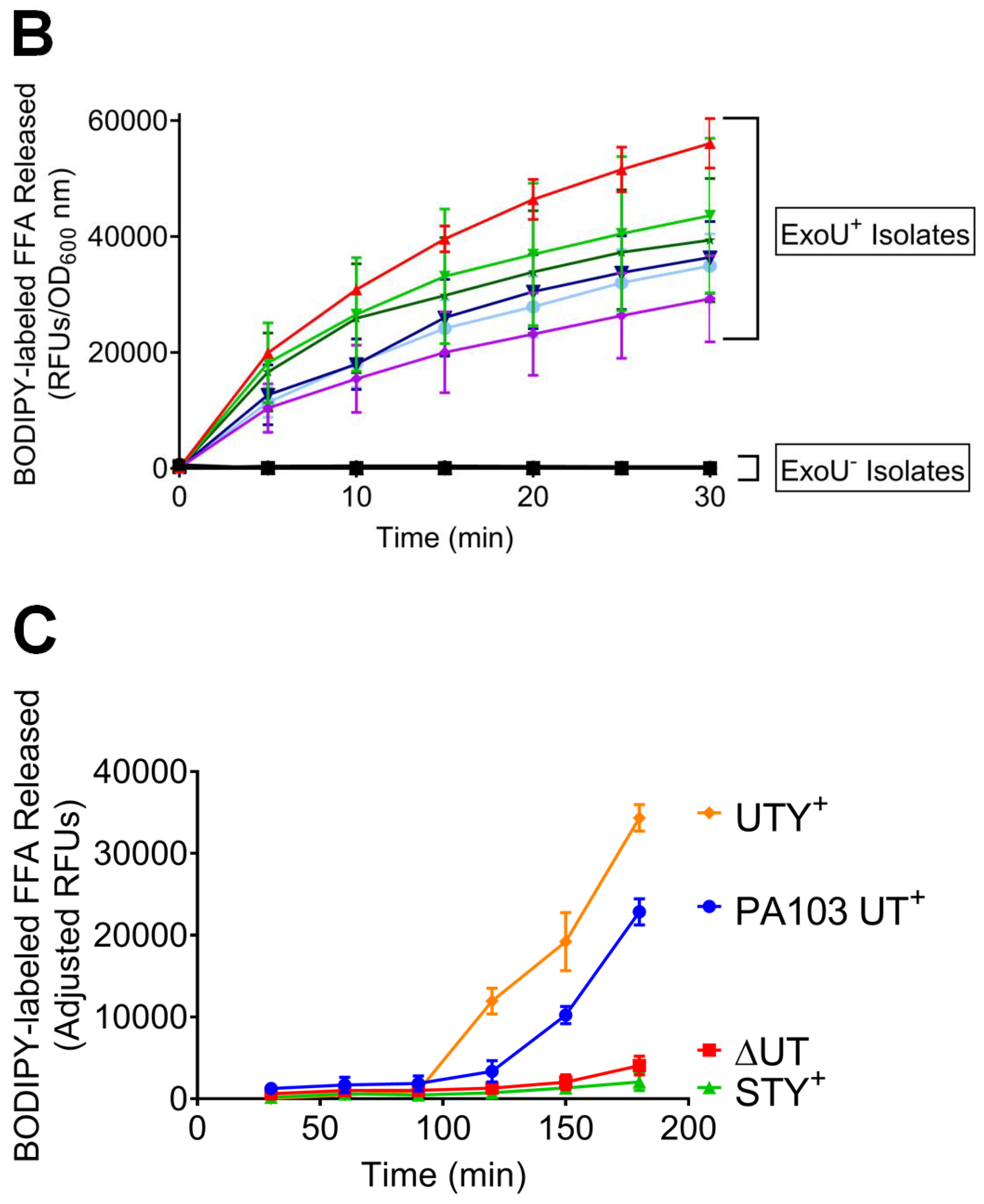
2.3. Clinical Isolates Exhibit ExoU PLA2 Enzyme Activity In Vitro and During the Infection of Cultured PMVECs
2.4. Clinical Isolates Have ExoY Activity in PMVECs
2.5. End-Organ Injury Attributable to ExoY and ExoU
2.6. Mortality and Drug Resistance in ExoY+ Clinical Isolates
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Study Design
5.2. Isolation and Identification of Clinical P. aeruginosa
5.3. PCR Method and Gel Electrophoresis
5.4. Measurement of Secreted ExoU Activity In Vitro
5.5. Rat Pulmonary Microvascular Endothelial Cell Isolation and Culture
5.6. Measurement of ExoU Activity in PMVECs
5.7. Measurement of ExoY Activity in PMVECs
5.8. Determination of End-Organ Dysfunction
5.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Sawa, T.; Shimizu, M.; Moriyama, K.; Wiener-Kronish, J.P. Association between Pseudomonas aeruginosa type III secretion, antibiotic resistance, and clinical outcome: A review. Crit. Care 2014, 18, 668. [Google Scholar] [CrossRef] [Green Version]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad bugs, no drugs: No ESKAPE! An update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the ESKAPE pathogens. Expert Rev. Anti Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef]
- El Solh, A.A.; Akinnusi, M.E.; Wiener-Kronish, J.P.; Lynch, S.V.; Pineda, L.A.; Szarpa, K. Persistent infection with Pseudomonas aeruginosa in ventilator-associated pneumonia. Am. J. Respir. Crit. Care Med. 2008, 178, 513–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertugrul, B.M.; Yildirim, A.; Ay, P.; Oncu, S.; Cagatay, A.; Cakar, N.; Ertekin, C.; Ozsut, H.; Eraksoy, H.; Calangu, S. Ventilator-associated pneumonia in surgical emergency intensive care unit. Saudi Med. J. 2006, 27, 52–57. [Google Scholar] [PubMed]
- Pena, C.; Cabot, G.; Gomez-Zorrilla, S.; Zamorano, L.; Ocampo-Sosa, A.; Murillas, J.; Almirante, B.; Pomar, V.; Aguilar, M.; Granados, A.; et al. Influence of virulence genotype and resistance profile in the mortality of Pseudomonas aeruginosa bloodstream infections. Clin. Infect. Dis. 2015, 60, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Roy-Burman, A.; Savel, R.H.; Racine, S.; Swanson, B.L.; Revadigar, N.S.; Fujimoto, J.; Sawa, T.; Frank, D.W.; Wiener-Kronish, J.P. Type III protein secretion is associated with death in lower respiratory and systemic Pseudomonas aeruginosa infections. J. Infect. Dis. 2001, 183, 1767–1774. [Google Scholar] [CrossRef] [Green Version]
- Angus, A.A.; Evans, D.J.; Barbieri, J.T.; Fleiszig, S.M. The ADP-ribosylation domain of Pseudomonas aeruginosa ExoS is required for membrane bleb niche formation and bacterial survival within epithelial cells. Infect. Immun. 2010, 78, 4500–4510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juan, C.; Pena, C.; Oliver, A. Host and Pathogen Biomarkers for Severe Pseudomonas aeruginosa Infections. J. Infect. Dis. 2017, 215, S44–S51. [Google Scholar] [CrossRef]
- Wong-Beringer, A.; Wiener-Kronish, J.; Lynch, S.; Flanagan, J. Comparison of type III secretion system virulence among fluoroquinolone-susceptible and -resistant clinical isolates of Pseudomonas aeruginosa. Clin. Microbiol. Infect. 2008, 14, 330–336. [Google Scholar] [CrossRef] [Green Version]
- Engel, J.; Balachandran, P. Role of Pseudomonas aeruginosa type III effectors in disease. Curr. Opin. Microbiol. 2009, 12, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Morrow, K.A.; Frank, D.W.; Balczon, R.; Stevens, T. The Pseudomonas aeruginosa Exoenzyme Y: A Promiscuous Nucleotidyl Cyclase Edema Factor and Virulence Determinant. Handb. Exp. Pharmacol. 2017, 238, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Kloth, C.; Schirmer, B.; Munder, A.; Stelzer, T.; Rothschuh, J.; Seifert, R. The Role of Pseudomonas aeruginosa ExoY in an Acute Mouse Lung Infection Model. Toxins 2018, 10, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrow, K.A.; Seifert, R.; Kaever, V.; Britain, A.L.; Sayner, S.L.; Ochoa, C.D.; Cioffi, E.A.; Frank, D.W.; Rich, T.C.; Stevens, T. Heterogeneity of pulmonary endothelial cyclic nucleotide response to Pseudomonas aeruginosa ExoY infection. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 309, L1199–L1207. [Google Scholar] [CrossRef] [Green Version]
- Munder, A.; Rothschuh, J.; Schirmer, B.; Klockgether, J.; Kaever, V.; Tummler, B.; Seifert, R.; Kloth, C. The Pseudomonas aeruginosa ExoY phenotype of high-copy-number recombinants is not detectable in natural isolates. Open Biol. 2018, 8, 170250. [Google Scholar] [CrossRef] [Green Version]
- Seifert, R.; Schneider, E.H.; Bahre, H. From canonical to non-canonical cyclic nucleotides as second messengers: Pharmacological implications. Pharmacol. Ther. 2015, 148, 154–184. [Google Scholar] [CrossRef]
- Belyy, A.; Raoux-Barbot, D.; Saveanu, C.; Namane, A.; Ogryzko, V.; Worpenberg, L.; David, V.; Henriot, V.; Fellous, S.; Merrifield, C.; et al. Actin activates Pseudomonas aeruginosa ExoY nucleotidyl cyclase toxin and ExoY-like effector domains from MARTX toxins. Nat. Commun. 2016, 7, 13582. [Google Scholar] [CrossRef] [Green Version]
- Cowell, B.A.; Evans, D.J.; Fleiszig, S.M. Actin cytoskeleton disruption by ExoY and its effects on Pseudomonas aeruginosa invasion. FEMS Microbiol. Lett. 2005, 250, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Hritonenko, V.; Mun, J.J.; Tam, C.; Simon, N.C.; Barbieri, J.T.; Evans, D.J.; Fleiszig, S.M. Adenylate cyclase activity of Pseudomonas aeruginosa ExoY can mediate bleb-niche formation in epithelial cells and contributes to virulence. Microb. Pathog. 2011, 51, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Yahr, T.L.; Vallis, A.J.; Hancock, M.K.; Barbieri, J.T.; Frank, D.W. ExoY, an adenylate cyclase secreted by the Pseudomonas aeruginosa type III system. Proc. Natl. Acad. Sci. USA 1998, 95, 13899–13904. [Google Scholar] [CrossRef] [Green Version]
- Morrow, K.A.; Ochoa, C.D.; Balczon, R.; Zhou, C.; Cauthen, L.; Alexeyev, M.; Schmalzer, K.M.; Frank, D.W.; Stevens, T. Pseudomonas aeruginosa exoenzymes U and Y induce a transmissible endothelial proteinopathy. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 310, L337–L353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balczon, R.; Pittet, J.F.; Wagener, B.M.; Moser, S.A.; Voth, S.; Vorhees, C.V.; Williams, M.T.; Bridges, J.P.; Alvarez, D.F.; Koloteva, A.; et al. Infection-induced endothelial amyloids impair memory. FASEB J. 2019, 33, 10300–10314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.T.; Balczon, R.; Pittet, J.F.; Wagener, B.M.; Moser, S.A.; Morrow, K.A.; Voth, S.; Francis, C.M.; Leavesley, S.; Bell, J.; et al. Nosocomial Pneumonia Elicits an Endothelial Proteinopathy: Evidence for a Source of Neurotoxic Amyloids in Critically Ill Patients. Am. J. Respir. Crit. Care Med. 2018, 198, 1575–1578. [Google Scholar] [CrossRef] [PubMed]
- Shaver, C.M.; Hauser, A.R. Relative contributions of Pseudomonas aeruginosa ExoU, ExoS, and ExoT to virulence in the lung. Infect. Immun. 2004, 72, 6969–6977. [Google Scholar] [CrossRef] [Green Version]
- Schulert, G.S.; Feltman, H.; Rabin, S.D.; Martin, C.G.; Battle, S.E.; Rello, J.; Hauser, A.R. Secretion of the toxin ExoU is a marker for highly virulent Pseudomonas aeruginosa isolates obtained from patients with hospital-acquired pneumonia. J. Infect. Dis. 2003, 188, 1695–1706. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.V. The roles of various fractions of Pseudomonas aeruginosa in its pathogenesis. 3. Identity of the lethal toxins produced in vitro and in vivo. J. Infect. Dis. 1966, 116, 481–489. [Google Scholar] [CrossRef]
- Liu, P.V. The roles of various fractions of Pseudomonas aeruginosa in its pathogenesis. II. Effects of lecithinase and protease. J. Infect. Dis. 1966, 116, 112–116. [Google Scholar] [CrossRef]
- Park, M.H.; Kim, S.Y.; Roh, E.Y.; Lee, H.S. Difference of Type 3 secretion system (T3SS) effector gene genotypes (exoU and exoS) and its implication to antibiotics resistances in isolates of Pseudomonas aeruginosa from chronic otitis media. Auris Nasus Larynx 2017, 44, 258–265. [Google Scholar] [CrossRef]
- Pobiega, M.; Maciag, J.; Chmielarczyk, A.; Romaniszyn, D.; Pomorska-Wesolowska, M.; Ziolkowski, G.; Heczko, P.B.; Bulanda, M.; Wojkowska-Mach, J. Molecular characterization of carbapenem-resistant Pseudomonas aeruginosa strains isolated from patients with urinary tract infections in Southern Poland. Diagn. Microbiol. Infect. Dis. 2015, 83, 295–297. [Google Scholar] [CrossRef]
- Pobiega, M.; Maciag, J.; Pomorska-Wesolowska, M.; Chmielarczyk, A.; Romaniszyn, D.; Ziolkowski, G.; Heczko, P.B.; Wojkowska-Mach, J.; Bulanda, M. Urinary tract infections caused by Pseudomonas aeruginosa among children in Southern Poland: Virulence factors and antibiotic resistance. J. Pediatr. Urol. 2016, 12, 36.e1–36.e6. [Google Scholar] [CrossRef]
- Yousefi-Avarvand, A.; Khashei, R.; Sedigh Ebrahim-Saraie, H.; Emami, A.; Zomorodian, K.; Motamedifar, M. The Frequency of Exotoxin A and Exoenzymes S and U Genes Among Clinical Isolates of Pseudomonas aeruginosa in Shiraz, Iran. Int. J. Mol. Cell. Med. 2015, 4, 167–173. [Google Scholar]
- Ochoa, C.D.; Alexeyev, M.; Pastukh, V.; Balczon, R.; Stevens, T. Pseudomonas aeruginosa exotoxin Y is a promiscuous cyclase that increases endothelial tau phosphorylation and permeability. J. Biol. Chem. 2012, 287, 25407–25418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brummel, N.E.; Girard, T.D. Preventing delirium in the intensive care unit. Crit. Care Clin. 2013, 29, 51–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, A.C.; Hay, A.W.; Swann, D.G.; Everingham, K.; McCulloch, C.; McNulty, J.; Brooks, O.; Laurenson, I.F.; Cook, B.; Walsh, T.S. Reducing ventilator-associated pneumonia in intensive care: Impact of implementing a care bundle. Crit. Care Med. 2011, 39, 2218–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pileggi, C.; Mascaro, V.; Bianco, A.; Nobile, C.G.A.; Pavia, M. Ventilator Bundle and Its Effects on Mortality Among ICU Patients: A Meta-Analysis. Crit. Care Med. 2018, 46, 1167–1174. [Google Scholar] [CrossRef]
- Rivosecchi, R.M.; Kane-Gill, S.L.; Svec, S.; Campbell, S.; Smithburger, P.L. The implementation of a nonpharmacologic protocol to prevent intensive care delirium. J. Crit. Care 2016, 31, 206–211. [Google Scholar] [CrossRef]
- Sen, S.; Johnston, C.; Greenhalgh, D.; Palmieri, T. Ventilator-Associated Pneumonia Prevention Bundle Significantly Reduces the Risk of Ventilator-Associated Pneumonia in Critically Ill Burn Patients. J. Burn Care Res. 2016, 37, 166–171. [Google Scholar] [CrossRef]
- Ibanez, J.; Riera, M.; Amezaga, R.; Herrero, J.; Colomar, A.; Campillo-Artero, C.; De Ibarra, J.I.; Bonnin, O. Long-Term Mortality After Pneumonia in Cardiac Surgery Patients: A Propensity-Matched Analysis. J. Intensive Care Med. 2016, 31, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Karhu, J.; Ala-Kokko, T.I.; Ylipalosaari, P.; Ohtonen, P.; Laurila, J.J.; Syrjala, H. Hospital and long-term outcomes of ICU-treated severe community- and hospital-acquired, and ventilator-associated pneumonia patients. Acta Anaesthesiol. Scand. 2011, 55, 1254–1260. [Google Scholar] [CrossRef]
- Wang, C.Y.; Calfee, C.S.; Paul, D.W.; Janz, D.R.; May, A.K.; Zhuo, H.; Bernard, G.R.; Matthay, M.A.; Ware, L.B.; Kangelaris, K.N. One-year mortality and predictors of death among hospital survivors of acute respiratory distress syndrome. Intensive Care Med. 2014, 40, 388–396. [Google Scholar] [CrossRef]
- Balczon, R.; Morrow, K.A.; Zhou, C.; Edmonds, B.; Alexeyev, M.; Pittet, J.F.; Wagener, B.M.; Moser, S.A.; Leavesley, S.; Zha, X.; et al. Pseudomonas aeruginosa infection liberates transmissible, cytotoxic prion amyloids. FASEB J. 2017, 31, 2785–2796. [Google Scholar] [CrossRef] [Green Version]
- Leber, A.L. (Ed.) Clinical Microbiology Procedures Handbook, 4th ed.; American Society for Microbiology: Washington, DC, USA, 2016. [Google Scholar]
- Iglewski, B.H.; Sadoff, J.; Bjorn, M.J.; Maxwell, E.S. Pseudomonas aeruginosa exoenzyme S: An adenosine diphosphate ribosyltransferase distinct from toxin A. Proc. Natl. Acad. Sci. USA 1978, 75, 3211–3215. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.M.; Sato, H.; Dirck, A.T.; Feix, J.B.; Frank, D.W. Ubiquitin activates patatin-like phospholipases from multiple bacterial species. J. Bacteriol. 2015, 197, 529–541. [Google Scholar] [CrossRef] [Green Version]
- Benson, M.A.; Schmalzer, K.M.; Frank, D.W. A sensitive fluorescence-based assay for the detection of ExoU-mediated PLA(2) activity. Clin. Chim. Acta 2010, 411, 190–197. [Google Scholar] [CrossRef] [Green Version]
- Housley, N.A.; Winkler, H.H.; Audia, J.P. The Rickettsia prowazekii ExoU homologue possesses phospholipase A1 (PLA1), PLA2, and lyso-PLA2 activities and can function in the absence of any eukaryotic cofactors in vitro. J. Bacteriol. 2011, 193, 4634–4642. [Google Scholar] [CrossRef] [Green Version]
- Jian, M.Y.; Alexeyev, M.F.; Wolkowicz, P.E.; Zmijewski, J.W.; Creighton, J.R. Metformin-stimulated AMPK-alpha1 promotes microvascular repair in acute lung injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 305, L844–L855. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, D.F.; Housley, N.; Koloteva, A.; Zhou, C.; O’Donnell, K.; Audia, J.P. Caspase-1 Activation Protects Lung Endothelial Barrier Function during Infection-Induced Stress. Am. J. Respir. Cell. Mol. Biol. 2016, 55, 500–510. [Google Scholar] [CrossRef] [Green Version]
- Khwaja, A. KDIGO clinical practice guidelines for acute kidney injury. Nephron Clin. Pract. 2012, 120, c179–c184. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | n (%) |
|---|---|
| Total | 99 |
| Age in years | 57 |
| Gender (Male) | 68 (68) |
| Admission Type | |
| Trauma | 49 (49) |
| Other | 50 (50) |
| Race | |
| White | 63 (63) |
| Black | 30 (30) |
| Hispanic | 6 (6) |
| Alcohol Consumer | |
| Current | 33 (33) |
| Former | 3 (3) |
| Unknown | 9 (9) |
| Smoking | |
| Current | 29 (29) |
| Former | 22 (22) |
| Unknown | 9 (9) |
| Illicit Drug Use | |
| Current | 11 (11) |
| Former | 4 (4) |
| Unknown | 12 (12) |
| Injury Scores | Median (Range) |
|---|---|
| APACHE II | 16 (3–37) |
| SOFA | 7 (0–16) |
| SAPS II | 35 (13–67) |
| ODIN | 1 (0–6) |
| 4 Point Lung Injury | 1.67 (0.33–3.33) |
| Trauma ISS | 18 (4–45) |
| Characteristics | Median (Range) |
| Duration of Mechanical Ventilation, median | 21 (3–118) |
| Length of ICU Stay | 24 (3–118) |
| Time from Ventilation until Onset of pneumonia, median | 8 (0–52) |
| Isolates (n) | Single-Drug Resistant (n, %) | Multi-Drug Resistant (n, %) | Died (n, %) | Survived (n, %) | |
|---|---|---|---|---|---|
| ExoY+/ExoU− | 74 | 6 (8.1) | 13 (17.6) | 17 (23) | 57 (77) |
| ExoY−/ExoU+ | 2 | 0 (0) | 0 (0) | 0 (0) | 2 (100) |
| ExoY+/ExoU+ | 18 | 0 (0) | 3 (16.7) | 3 (16.7) | 15 (83.3) |
| ExoY−/ExoU− | 5 | 2 (40) | 1 (20) | 2 (40) | 3 (60) |
| All Isolates | 99 | 8 (8.1) | 17 (17) | 22 (22) | 77 (78) |
| All Patients | ExoY+/ExoU− Patients | All ExoU+ Patients | |
|---|---|---|---|
| Acute Kidney Injury | 42/94 | 35/71 | 6/18 |
| CV dysfunction | 42/99 | 28/74 | 6/20 |
| Coagulopathy | 72/92 | 54/70 | 14/17 |
| Primer Name | Primer Sequence | PCR Fragment Size |
|---|---|---|
| exoY forward | TGAGCGAGGACGGATTCTA | 309 bp |
| exoY reverse | GATAGCCGTTGCCCTTGAT | |
| exoU forward | CTCAATGTACTCCCACGCATAG | 406 bp |
| exoU reverse | CATCCTGGAATTCTGTCCACTC | |
| exoT forward | GCCGAGATCAAGCAGATGAT | 405 bp |
| exoT reverse | GACAGGCTCGCCCTTTAC | |
| exoS forward | CATCAGGTAATGAGCGAGGTC | 410 bp |
| exoS reverse | TTCAGGGAGGTGGAGAGATAG | |
| PolA forward | TTTCCTGCAGCCAGTTATCC | 707 bp |
| PolA reverse | CAAGCTCAAGAGCACCTACA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagener, B.M.; Anjum, N.; Christiaans, S.C.; Banks, M.E.; Parker, J.C.; Threet, A.T.; Walker, R.R.; Isbell, K.D.; Moser, S.A.; Stevens, T.; et al. Exoenzyme Y Contributes to End-Organ Dysfunction Caused by Pseudomonas aeruginosa Pneumonia in Critically Ill Patients: An Exploratory Study. Toxins 2020, 12, 369. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12060369
Wagener BM, Anjum N, Christiaans SC, Banks ME, Parker JC, Threet AT, Walker RR, Isbell KD, Moser SA, Stevens T, et al. Exoenzyme Y Contributes to End-Organ Dysfunction Caused by Pseudomonas aeruginosa Pneumonia in Critically Ill Patients: An Exploratory Study. Toxins. 2020; 12(6):369. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12060369
Chicago/Turabian StyleWagener, Brant M., Naseem Anjum, Sarah C. Christiaans, Morgan E. Banks, Jordan C. Parker, Adam T. Threet, Rashidra R. Walker, Kayla D. Isbell, Stephen A. Moser, Troy Stevens, and et al. 2020. "Exoenzyme Y Contributes to End-Organ Dysfunction Caused by Pseudomonas aeruginosa Pneumonia in Critically Ill Patients: An Exploratory Study" Toxins 12, no. 6: 369. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12060369