Anti-Biofilm Activity of the Fungal Phytotoxin Sphaeropsidin A against Clinical Isolates of Antibiotic-Resistant Bacteria

 , , , , , and
, , , , , and
Abstract
:1. Introduction
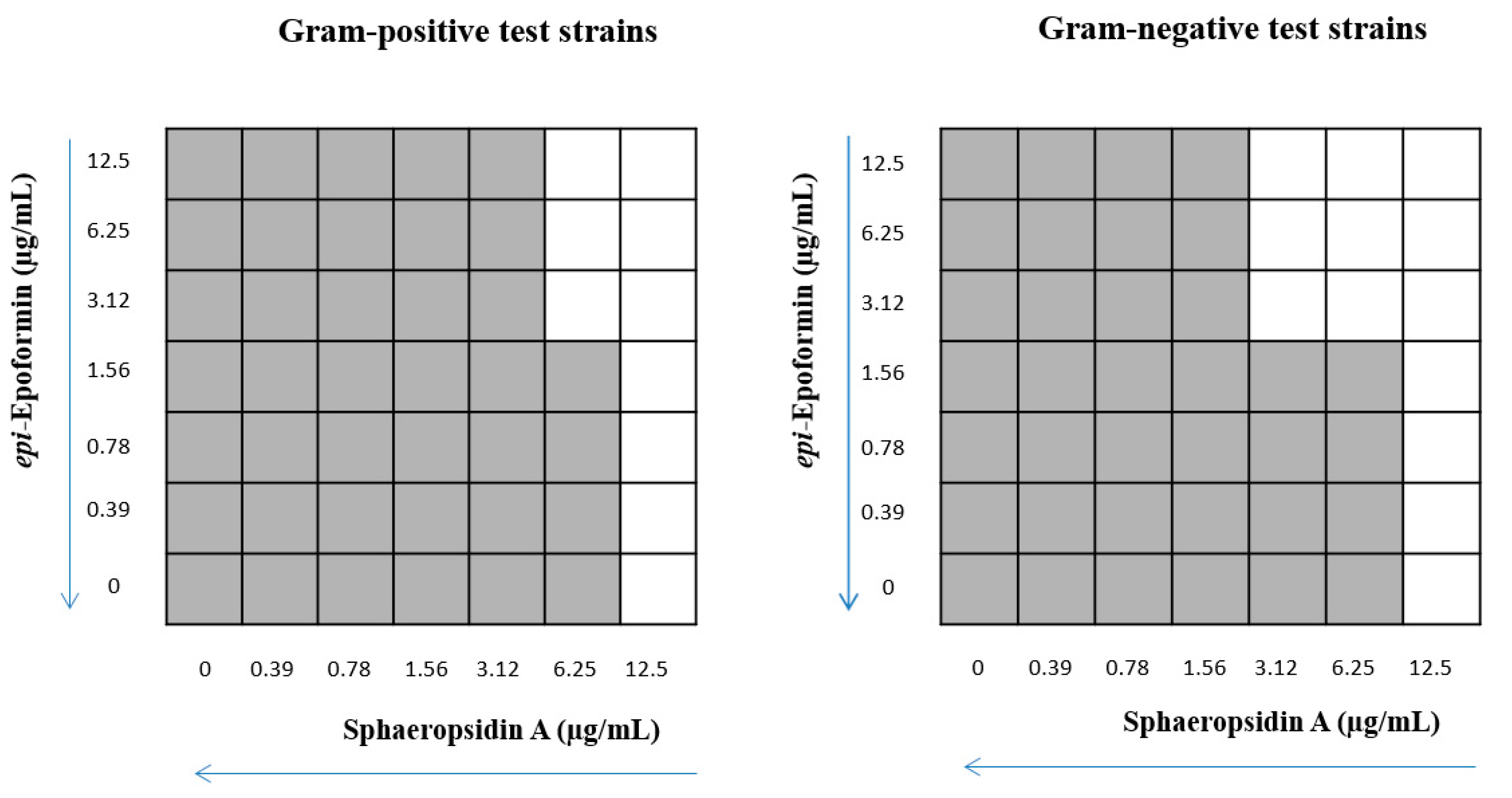
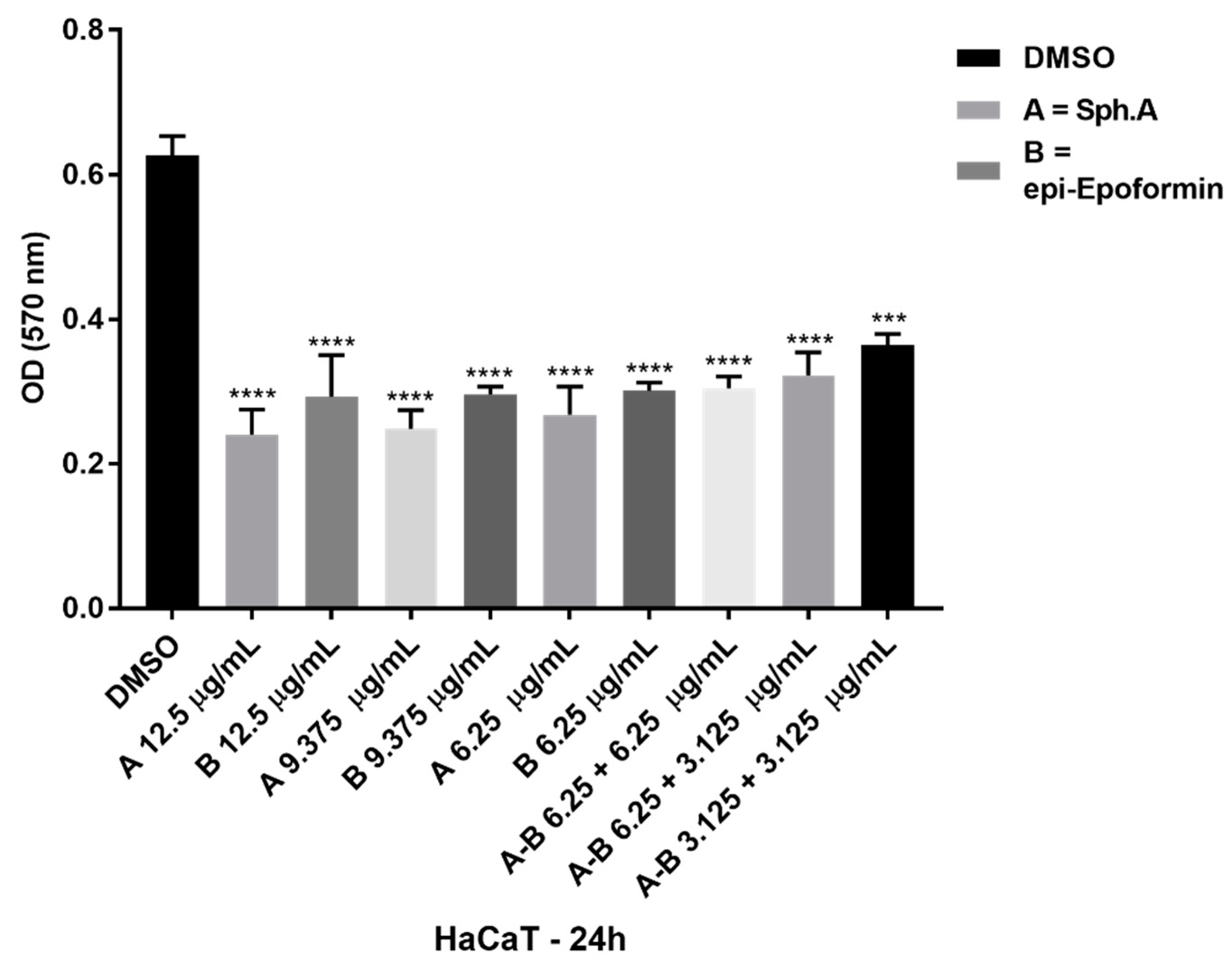
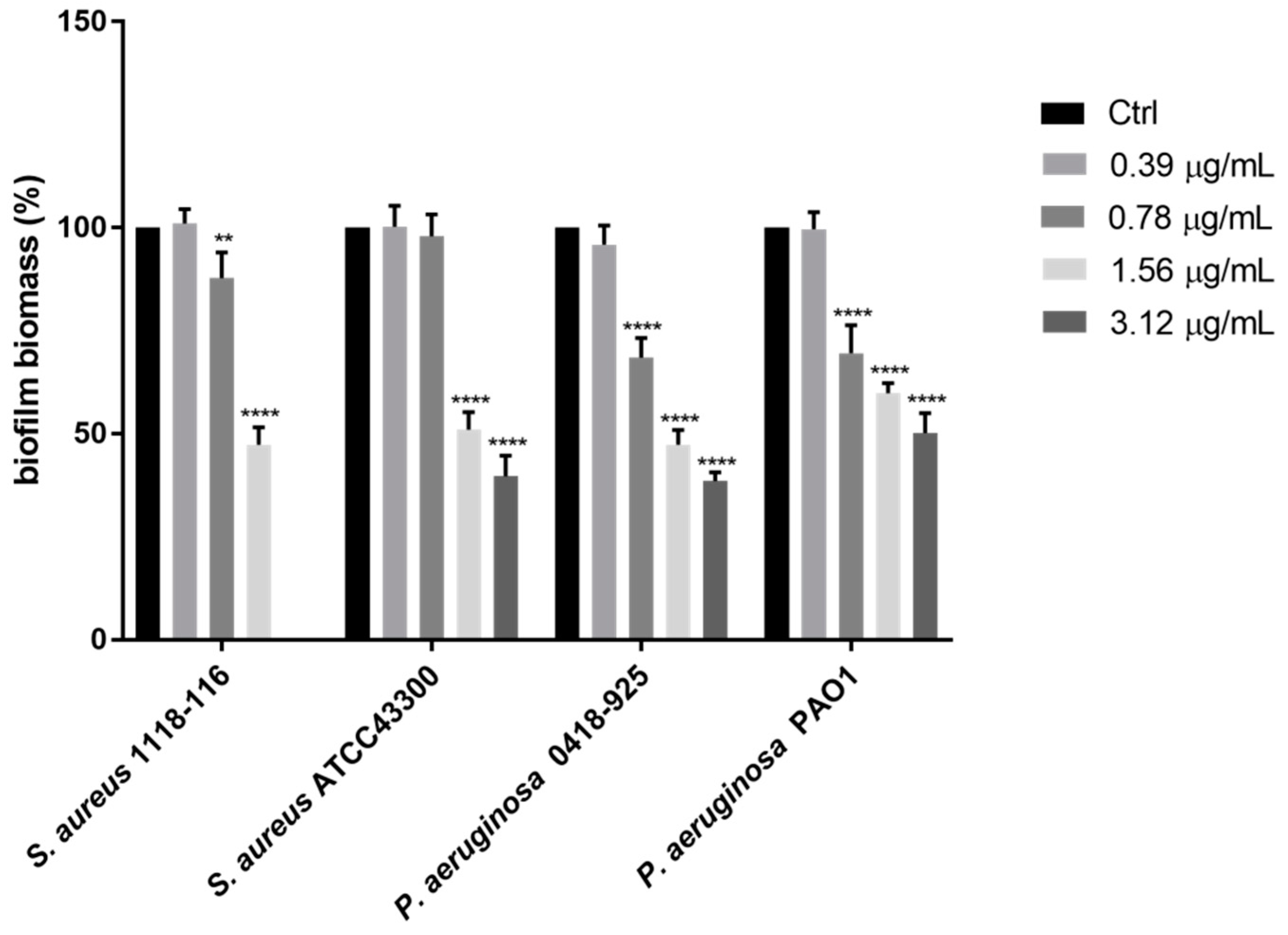
2. Results and Discussion
3. Conclusions
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Microbial Strains and Culture Conditions
4.3. Antimicrobial Assays
4.4. Synergy Assays
4.5. Biofilm Formation Inhibition Assay
4.6. Cytotoxicity Test
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO); Antimicrobial Resistance. No Time to Wait: Securing the Future from Drug-Resistant Infections. Report to the Secretary-General of the United Nations. Available online: https://www.who.int/antimicrobial-resistance/interagency-coordination-group/IACG_final_report_EN.pdf?ua=1 (accessed on 8 April 2020).
- Ciofu, O.; Mandsberg, L.F.; Wang, H.; Høiby, N. Phenotypes selected during chronic lung infection in cystic fibrosis patients: Implications for the treatment of Pseudomonas aeruginosa biofilm infections. FEMS Immunol. Med. Microbiol. 2012, 65, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Cantone, E.; Negri, R.; Roscetto, E.; Grassia, R.; Catania, M.R.; Capasso, P.; Maffei, M.; Soriano, A.A.; Leone, C.A.; Iengo, M.; et al. In vivo biofilm formation, Gram-negative infections and TAS2R38 polymorphisms in CRSw NP patients. Laryngoscope 2018, 128, E339–E345. [Google Scholar] [CrossRef] [PubMed]
- Gebreyohannes, G.; Nyerere, A.; Bii, C.; Sbhatu, D.B. Challenges of intervention, treatment, and antibiotic resistance of biofilm-forming microorganisms. Heliyon 2019, 5, e02192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, C.W.; Mah, T.F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, L. Photodynamic combinational therapy in cancer treatment. J. Buon. 2018, 23, 561–567. [Google Scholar]
- Barra, F.; Roscetto, E.; Soriano, A.A.; Vollaro, A.; Postiglione, I.; Pierantoni, G.M.; Palumbo, G.; Catania, M.R. Photodynamic and antibiotic therapy in combination to fight biofilms and resistant surface bacterial infections. Int. J. Mol. Sci. 2015, 16, 20417–20430. [Google Scholar] [CrossRef]
- Hu, X.; Huang, Y.Y.; Wang, Y.; Wang, X.; Hamblin, M.R. Antimicrobial photodynamic therapy to control clinically relevant biofilm infections. Front. Microbiol. 2018, 9, 1299. [Google Scholar] [CrossRef] [Green Version]
- Leaper, D.; Wilson, P.; Assadian, O.; Edmiston, C.; Kiernan, M.; Miller, A.; Bond-Smith, G.; Yap, J. The role of antimicrobial sutures in preventing surgical site infection. Ann. R. Coll. Surg. Engl. 2017, 99, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Balato, G.; Roscetto, E.; Vollaro, A.; Galasso, O.; Gasparini, G.; Ascione, T.; Catania, M.R.; Mariconda, M. Bacterial biofilm formation is variably inhibited by different formulations of antibiotic-loaded bone cement in vitro. Knee Surg. Sports Traumatol. Arthrosc. 2019, 27, 1943–1952. [Google Scholar] [CrossRef] [PubMed]
- Malek, A.E.; Raad, I.I. Preventing catheter-related infections in cancer patients: A review of current strategies. Expert Rev. Anti. Infect. Ther. 2020. [Google Scholar] [CrossRef] [PubMed]
- Artini, M.; Cicatiello, P.; Ricciardelli, A.; Papa, R.; Selan, L.; Dardano, P.; Tilotta, M.; Vrenna, G.; Tutino, M.L.; Giardina, P.; et al. Hydrophobin coating prevents Staphylococcus epidermidis biofilm formation on different surfaces. Biofouling 2017, 33, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Vila, J.; Moreno-Morales, J.; Ballesté-Delpierre, C. Current landscape in the discovery of novel antibacterial agents. Clin. Microbiol. Infect. 2019, 26, 596–603. [Google Scholar] [CrossRef]
- Ricciardelli, A.; Casillo, A.; Papa, R.; Monti, D.M.; Imbimbo, P.; Vrenna, G.; Artini, M.; Selan, L.; Corsaro, M.M.; Tutino, M.L.; et al. Pentadecanal inspired molecules as new antibiofilm agents against Staphylococcus epidermidis. Biofouling 2018, 34, 1110–1120. [Google Scholar] [CrossRef] [PubMed]
- Chung, P.Y.; Khanum, R. Antimicrobial peptides as potential anti-biofilm agents against multidrug-resistant bacteria. J. Microbiol. Immunol. Infect. 2017, 50, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Roscetto, E.; Contursi, P.; Vollaro, A.; Fusco, S.; Notomista, E.; Catania, M.R. Antifungal and anti-biofilm activity of the first cryptic antimicrobial peptide from an archaeal protein against Candida spp. clinical isolates. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- He, M.; Zhang, H.; Li, Y.; Wang, G.; Tang, B.; Zhao, J.; Huang, Y.; Zheng, J. Cathelicidin-derived antimicrobial peptides inhibit Zika virus through direct inactivation and interferon pathway. Front. Immunol. 2018, 9, 722. [Google Scholar] [CrossRef] [Green Version]
- Gaglione, R.; Pirone, L.; Farina, B.; Fusco, S.; Smaldone, G.; Aulitto, M.; Dell’Olmo, E.; Roscetto, E.; Del Gatto, A.; Fattorusso, R.; et al. Insights into the anticancer properties of the first antimicrobial peptide from Archaea. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 2155–2164. [Google Scholar] [CrossRef] [Green Version]
- Li, J.W.H.; Vederas, J.C. Drug discovery and natural products: End of an era or an endless frontier? Science 2009, 325, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Cragg, G.M.; Newman, D.J. Natural products: A continuing source of novel drug leads. Biochim. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 3670–3695. [Google Scholar] [CrossRef] [Green Version]
- Schueffler, A.; Anke, T. Fungal natural products in research and development. Nat. Prod. Rep. 2014, 31, 1425–1448. [Google Scholar] [CrossRef]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural products for drug discovery in the 21st century: Innovations for novel drug discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [Green Version]
- Wright, G.D. Unlocking the potential of natural products in drug discovery. Microb. Biotechnol. 2019, 12, 55–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masi, M.; Maddau, L.; Linaldeddu, B.T.; Scanu, B.; Evidente, A.; Cimmino, A. Bioactive metabolites from pathogenic and endophytic fungi of forest trees. Curr. Med. Chem. 2018, 25, 208–252. [Google Scholar] [CrossRef] [PubMed]
- Andolfi, A.; Maddau, L.; Basso, S.; Linaldeddu, B.T.; Cimmino, A.; Scanu, B.; Deidda, A.; Tuzi, A.; Evidente, A. Diplopimarane, a 20-nor-ent-pimarane produced by the oak pathogen Diplodia quercivora. J. Nat. Prod. 2014, 77, 2352–2360. [Google Scholar] [CrossRef] [PubMed]
- Graniti, A.; Sparapano, L.; Evidente, A. Cyclopaldic acid, a major phytotoxic metabolite of Seiridium cupressi, the pathogen of a canker disease of cypress. Plant Pathol. 1992, 41, 563–568. [Google Scholar] [CrossRef]
- Evidente, A.; Andolfi, A.; Maddau, L.; Franceschini, A.; Marras, F. Biscopyran, a phytotoxic hexasubstituted pyranopyran produced by Biscogniauxia mediterranea, a fungus pathogen of cork oak. J. Nat. Prod. 2005, 68, 568–571. [Google Scholar] [CrossRef]
- Evidente, A.; Sparapano, L.; Fierro, O.; Bruno, G.; Giordano, F.; Motta, A. Sphaeropsidins B and C, phytotoxic pimarane diterpenes from Sphaeropsis sapinea f. sp. cupressi and Diplodia mutila. Phytochemistry 1997, 45, 705–713. [Google Scholar] [CrossRef]
- Cimmino, A.; Maddau, L.; Masi, M.; Evidente, M.; Linaldeddu, B.T.; Evidente, A. Further secondary metabolites produced by Diplodia corticola, a fungal pathogen involved in cork oak decline. Tetrahedron 2016, 72, 6788–6793. [Google Scholar] [CrossRef]
- Evidente, A.; Maddau, L.; Scanu, B.; Andolfi, A.; Masi, M.; Motta, A.; Tuzi, A. Sphaeropsidones, phytotoxic dimedone methyl ethers produced by Diplodia cupressi: A structure-activity relationship study. J. Nat. Prod. 2011, 74, 757–763. [Google Scholar] [CrossRef]
- Masi, M.; Maddau, L.; Linaldeddu, B.T.; Cimmino, A.; D’Amico, W.; Scanu, B.; Evidente, M.; Tuzi, A.; Evidente, A. Bioactive secondary metabolites produced by the oak pathogen Diplodia corticola. J. Agric. Food Chem. 2016, 64, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A.; Randazzo, G.; Ballio, A. Structure determination of seiridin and isoseiridin, phytotoxic butenolides from culture filtrate of Seiridium cardinale. J. Nat. Prod. 1986, 49, 593–601. [Google Scholar] [CrossRef]
- Ballio, A.; Evidente, A.; Graniti, A.; Randazzo, G.; Sparapano, L. Seiricuprolide, a new phytotoxic macrolide from a strain of Seiridium cupressi infecting cypress. Phytochemistry 1988, 27, 3117–3121. [Google Scholar] [CrossRef]
- Cimmino, A.; Maddau, L.; Masi, M.; Linaldeddu, B.T.; Evidente, A. Secondary metabolites produced by Sardiniella urbana, a new emerging pathogen on European hackberry. Nat. Prod. Res. 2019, 33, 1862–1869. [Google Scholar] [CrossRef]
- Masi, M.; Di Lecce, R.; Tuzi, A.; Linaldeddu, B.T.; Montecchio, L.; Maddau, L.; Evidente, A. Hyfraxinic acid, a phytotoxic tetrasubstituted octanoic Acid, produced by the ash (Fraxinus excelsior L.) pathogen Hymenoscyphus fraxineus together with viridiol and some of its analogues. J. Agric. Food Chem. 2019, 67, 13617–13623. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A.; Maddau, L.; Spanu, E.; Franceschini, A.; Lazzaroni, S.; Motta, A. Diplopyrone, a new phytotoxic tetrahydropyranpyran-2-one produced by Diplodia mutila, a fungus pathogen of cork oak. J. Nat. Prod. 2003, 66, 313–315. [Google Scholar] [CrossRef] [PubMed]
- Sparapano, L.; Bruno, G.; Fierro, O.; Evidente, A. Studies on structure–activity relationship of sphaeropsidins A–F, phytotoxins produced by Sphaeropsis sapinea f. sp. cupressi. Phytochemistry 2004, 65, 189–198. [Google Scholar] [CrossRef]
- Reveglia, P.; Masi, M.; Evidente, A. Melleins—Intriguing natural compounds. Biomolecules 2020, 10, 772. [Google Scholar] [CrossRef] [PubMed]
- Barilli, E.; Cimmino, A.; Masi, M.; Evidente, M.; Rubiales, D.; Evidente, A. Inhibition of spore germination and appressorium formation of rust species by plant and fungal metabolites. Nat. Prod. Commun. 2016, 11, 1934578X1601100940. [Google Scholar] [CrossRef] [Green Version]
- Barilli, E.; Cimmino, A.; Masi, M.; Evidente, M.; Rubiales, D.; Evidente, A. Inhibition of early development stages of rust fungi by the two fungal metabolites cyclopaldic acid and epi-epoformin. Pest Manag. Sci. 2017, 73, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Andolfi, A.; Avolio, F.; Ali, A.; Tabanca, N.; Khan, I.A.; Evidente, A. Cyclopaldic Acid, seiridin, and sphaeropsidin a as fungal phytotoxins, and larvicidal and biting deterrents against Aedes aegypti (Diptera: Culicidae): Structure-activity relationships. Chem. Biodivers. 2013, 10, 1239–1251. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Masi, M.; Maddau, L.; Cimmino, A.; Evidente, M.; Rubiales, D.; Evidente, A. Induction of haustorium development by sphaeropsidones in radicles of the parasitic weeds Striga and Orobanche. A structure–activity relationship study. J. Agric. Food Chem. 2016, 64, 5188–5196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evidente, A.; Venturi, V.; Masi, M.; Degrassi, G.; Cimmino, A.; Maddau, L.; Andolfi, A. In vitro antibacterial activity of sphaeropsidins and chemical derivatives toward Xanthomonas oryzae pv. oryzae, the causal agent of rice bacterial blight. J. Nat. Prod. 2011, 74, 2520–2525. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, V.; Chantôme, A.; Lefranc, F.; Cimmino, A.; Miklos, W.; Paulitschke, V.; Mohr, T.; Maddau, L.; Kornienko, A.; Berger, W.; et al. Sphaeropsidin A shows promising activity against drug-resistant cancer cells by targeting regulatory volume increase. Cell. Mol. Life Sci. 2015, 72, 3731–3746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingels, A.; Dinhof, C.; Garg, A.D.; Maddau, L.; Masi, M.; Evidente, A.; Berger, W.; Dejaegher, B.; Mathieu, V. Computed determination of the in vitro optimal chemocombinations of sphaeropsidin A with chemotherapeutic agents to combat melanomas. Cancer Chemoth. Pharmacol. 2017, 79, 971–983. [Google Scholar] [CrossRef]
- Evidente, A.; Andolfi, A.; Cimmino, A. Relationships between the stereochemistry and biological activity of fungal phytotoxins. Chirality 2011, 23, 674–693. [Google Scholar] [CrossRef]
- Evidente, A.; Cimmino, A.; Andolfi, A. The effect of stereochemistry on the biological activity of natural phytotoxins, fungicides, insecticides and herbicides. Chirality 2013, 25, 59–78. [Google Scholar] [CrossRef]
- Masi, M.; Cimmino, A.; Maddau, L.; Kornienko, A.; Tuzi, A.; Evidente, A. Crystal structure and absolute configuration of sphaeropsidin A and its 6-o-p-bromobenzoate. Tetrahedron Lett. 2016, 57, 4592–4594. [Google Scholar] [CrossRef]
- Mazzeo, G.; Cimmino, A.; Masi, M.; Longhi, G.; Maddau, L.; Memo, M.; Evidente, A.; Abbate, S. Importance and difficulties in the use of chiroptical methods to assign the absolute configuration of natural products: The case of phytotoxic pyrones and furanones produced by Diplodia corticola. J. Nat. Prod. 2017, 80, 2406–2415. [Google Scholar] [CrossRef]
- Fusè, M.; Mazzeo, G.; Longhi, G.; Abbate, S.; Masi, M.; Evidente, A.; Puzzarini, C.; Barone, V. Unbiased determination of absolute configurations by vis-à-vis comparison of experimental and simulated spectra: The challenging case of diplopyrone. J. Phys. Chem. B 2019, 123, 9230–9237. [Google Scholar] [CrossRef]
- Fazli, M.; Bjarnsholt, T.; Kirketerp-Møller, K.; Jørgensen, B.; Andersen, A.S.; Krogfelt, K.A.; Givskov, M.; Tolker-Nielsen, T. Nonrandom distribution of Pseudomonas aeruginosa and Staphylococcus aureus in chronic wounds. J. Clin. Microbiol. 2009, 47, 4084–4089. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.M.; Zenilman, J.M.; Lazarus, G.S. Molecular microbiology: New dimensions for cutaneous biology and wound healing. J. Investig. Dermatol. 2010, 130, 38–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachiewicz, A.M.; Hauck, C.G.; Weber, D.J.; Cairns, B.A.; van Duin, D. Bacterial infections after burn injuries: Impact of multidrug resistance. Clin. Infect. Dis. 2017, 65, 2130–2136. [Google Scholar] [CrossRef] [PubMed]
- Eisner, R.; Lippmann, N.; Josten, C.; Rodloff, A.C.; Behrendt, D. Development of the bacterial spectrum and antimicrobial resistance in surgical site infections of trauma patients. Surg. Infect. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Singh, S. Prevalence of extended spectrum betalactamase (ESBL) and metallobetalactamase (MBL) producing Pseudomonas aeruginosa and Acinetobacter baumannii isolated from various clinical samples. J. Pathog. 2018, 2018, 6845985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royer, G.; Fourreau, F.; Boulanger, B.; Mercier-Darty, M.; Ducellier, D.; Cizeau, F.; Potron, A.; Podglajen, I.; Mongardon, N.; Decousser, J.W. Local outbreak of extended-spectrum β-lactamase SHV2a-producing Pseudomonas aeruginosa reveals the emergence of a new specific sub-lineage of the international ST235 high-risk clone. J. Hosp. Infect. 2020, 104, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, M.; Frees, D.; Ingmer, H. Antibiotic resistance and the MRSA problem. Gram Posit. Pathog. 2019, 7, 747–765. [Google Scholar]
- Serra, R.; Grande, R.; Butrico, L.; Rossi, A.; Settimio, U.F.; Caroleo, B.; Amato, B.; Gallelli, L.; de Franciscis, S. Chronic wound infections: The role of Pseudomonas aeruginosa and Staphylococcus aureus. Expert. Rev. Anti. Infect. Ther. 2015, 13, 605–613. [Google Scholar] [CrossRef] [PubMed]
- De Vor, L.; Rooijakkers, S.H.M.; van Strijp, J.A.G. Staphylococci evade the innate immune response by disarming neutrophils and forming biofilms. FEBS Lett. 2020. [Google Scholar] [CrossRef] [Green Version]
- Wolcott, R.D. Biofilms cause chronic infections. J. Wound Care 2017, 26, 423–425. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Santra, S.; Das, A.; Dixith, S.; Sinha, M.; Ghatak, S.; Ghosh, N.; Banerjee, P.; Khanna, S.; Mathew-Steiner, S.; et al. Staphylococcus aureus biofilm infection compromises wound healing by causing deficiencies in granulation tissue collagen. Ann. Surg. 2019. [Google Scholar] [CrossRef]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach, 3rd ed.; John Wiley & Sons: Chichester, UK, 2009. [Google Scholar]
- Le Calve, B.; Lallemand, B.; Perrone, C.; Lenglet, G.; Depauw, S.; Van Goietsenoven, G.; Bury, M.; Vurro, M.; Herphelin, F.; Andolfi, A.; et al. In vitro anticancer activity, toxicity and structure–activity relationships of phyllostictine A, a natural oxazatricycloalkenone produced by the fungus Phyllosticta cirsii. Toxicol. Appl. Pharmacol. 2011, 254, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Pillai, S.K.; Moellering, R.C.; Eliopoulos, G.M. Antimicrobial combinations. In Antibiotics in Laboratory. Medicine, 5th ed.; Lorian, V., Ed.; The Lippincott Williams & Wilkins Co.: Philadelphia, PA, USA, 2005; pp. 365–440. [Google Scholar]
- Stepanović, S.; Vuković, D.; Hola, V.; di Bonaventura, G.; Djukić, S.; Cirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by Staphylococci. Apmis 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Name | Chemical Family | Fungal Source | Ref. |
|---|---|---|---|---|
| 1 | epi-Epoformin | Cyclohexene oxide | Diplodia quercivora | [25] |
| 2 | Cyclopaldic acid | Isobenzofuranone | Seiridium cupressi | [26] |
| 3 | Biscopyran | Pyranopyran | Biscogniauxia mediterranea | [27] |
| 4 | Sphaeropsidin B | Diterpenoid | Diplodia cupressi | [28] |
| 5 | Sphaeropsidin C | Diterpenoid | D. cupressi | [28] |
| 6 | Sphaeropsidin G | Diterpenoid | Diplodia corticola | [29] |
| 7 | Sphaeropsidone | Cyclohexene oxide | D. cupressi | [30] |
| 8 | Sapinofuranone C | Furanone | D. corticola | [31] |
| 9 | (S,S)-Sapinofuranone B | Furanone | D. corticola | [31] |
| 10 | Seiridin | Butenolide | S. cupressi | [32] |
| 11 | Seiricuprolide | Macrolide | S. cupressi | [33] |
| 12 | Diplobifuranylone A | Furanone | D. corticola | [31] |
| 13 | Diplobifuranylone B | Furanone | D. corticola | [31] |
| 14 | Diplobifuranylone C | Furanone | D. corticola | [31] |
| 15 | (R)-Mellein | 3,4-Dihydroisocoumarin | Sardiniella urbana | [34] |
| 16 | cis-4-Hydroxymellein | 3,4-Dihydroisocoumarin | S. urbana | [34] |
| 17 | trans-4-Hydroxymellein | 3,4-Dihydroisocoumarin | S. urbana | [34] |
| 18 | Viridiol | Furanosteroid | Hymenoscyphus fraxineus | [35] |
| 19 | Diplopyrone | Pyranopyrone | D. corticola | [36] |
| 20 | Sphaeropsidin A | Diterpenoid | D. corticola | [31] |
| Compound | Bacterial Strain | |||||
|---|---|---|---|---|---|---|
| S. aureus ATCC 43300 | MRSA1118-116 | S. haemolyticus ATCC 29970 | S. haemolyticus VR 1219-118 | P. aeruginosa PAO1 | P. aeruginosa 0418-925 | |
| 1 | ≥90 | ≥90 | ≥90 | ≥90 | ≥90 | ≥90 |
| 2 | ≥90 | ≥90 | ≥90 | ≥90 | - | ≥90 |
| 3 | - | - | - | - | 50 | 60 |
| 4 | ≥90 | ≥90 | ≥90 | ≥90 | - | 60 |
| 5 | - | - | - | - | 60 | 60 |
| 6 | ≥90 | ≥90 | ≥90 | ≥90 | - | - |
| 7 | ≥90 | ≥90 | ≥90 | ≥90 | 50 | ≥90 |
| 8 | - | - | - | - | 60 | 60 |
| 9 | - | - | - | - | 60 | 60 |
| 10 | - | - | - | - | 50 | 60 |
| 11 | - | - | - | - | 60 | 50 |
| 12 | - | - | - | - | 50 | 50 |
| 13 | - | - | - | - | 50 | 60 |
| 14 | - | - | - | - | 50 | 60 |
| 15 | - | - | - | - | 60 | 60 |
| 16 | - | - | - | - | 60 | 70 |
| 17 | - | - | - | - | 60 | 60 |
| 18 | ≥90 | ≥90 | ≥90 | ≥90 | - | - |
| 19 | - | - | - | - | - | - |
| 20 | ≥90 | ≥90 | ≥90 | ≥90 | ≥90 | ≥90 |
| AK | nt | nt | nt | nt | >90 | >90 |
| TE | >90 | >90 | >90 | >90 | nt | nt |
| Bacterial Strain | Compound 1 | Compound 7 | Compound 20 | Amikacin | Teicoplanin | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | |
| S. aureus ATCC 43300 | 100 | 100 | 100 | >200 | 12.5 | 100 | nt | nt | 1 | 4 |
| MRSA 1118-116 | 100 | 100 | 100 | >200 | 6.25 | 25 | nt | nt | 0.5 | 4 |
| S. haemolyticus ATCC 29970 | 100 | 100 | 100 | >200 | 12.5 | 50 | nt | nt | 2 | >4 |
| S. haemolyticus VR 1219-118 | 100 | 100 | 100 | >200 | 12.5 | 50 | nt | nt | 2 | >4 |
| P. aeruginosa PAO1 | 50 | >200 | >100 | >200 | 12.5 | >200 | 4 | 32 | nt | nt |
| P. aeruginosa 0418-925 | 50 | >200 | 100 | >200 | 12.5 | >200 | 16 | >32 | nt | nt |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roscetto, E.; Masi, M.; Esposito, M.; Di Lecce, R.; Delicato, A.; Maddau, L.; Calabrò, V.; Evidente, A.; Catania, M.R. Anti-Biofilm Activity of the Fungal Phytotoxin Sphaeropsidin A against Clinical Isolates of Antibiotic-Resistant Bacteria. Toxins 2020, 12, 444. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070444
Roscetto E, Masi M, Esposito M, Di Lecce R, Delicato A, Maddau L, Calabrò V, Evidente A, Catania MR. Anti-Biofilm Activity of the Fungal Phytotoxin Sphaeropsidin A against Clinical Isolates of Antibiotic-Resistant Bacteria. Toxins. 2020; 12(7):444. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070444
Chicago/Turabian StyleRoscetto, Emanuela, Marco Masi, Matilde Esposito, Roberta Di Lecce, Antonella Delicato, Lucia Maddau, Viola Calabrò, Antonio Evidente, and Maria Rosaria Catania. 2020. "Anti-Biofilm Activity of the Fungal Phytotoxin Sphaeropsidin A against Clinical Isolates of Antibiotic-Resistant Bacteria" Toxins 12, no. 7: 444. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12070444