The Influence of NaCl and Glucose Content on Growth and Ochratoxin A Production by Aspergillus ochraceus, Aspergillus carbonarius and Penicillium nordicum


Abstract
:1. Introduction
2. Results
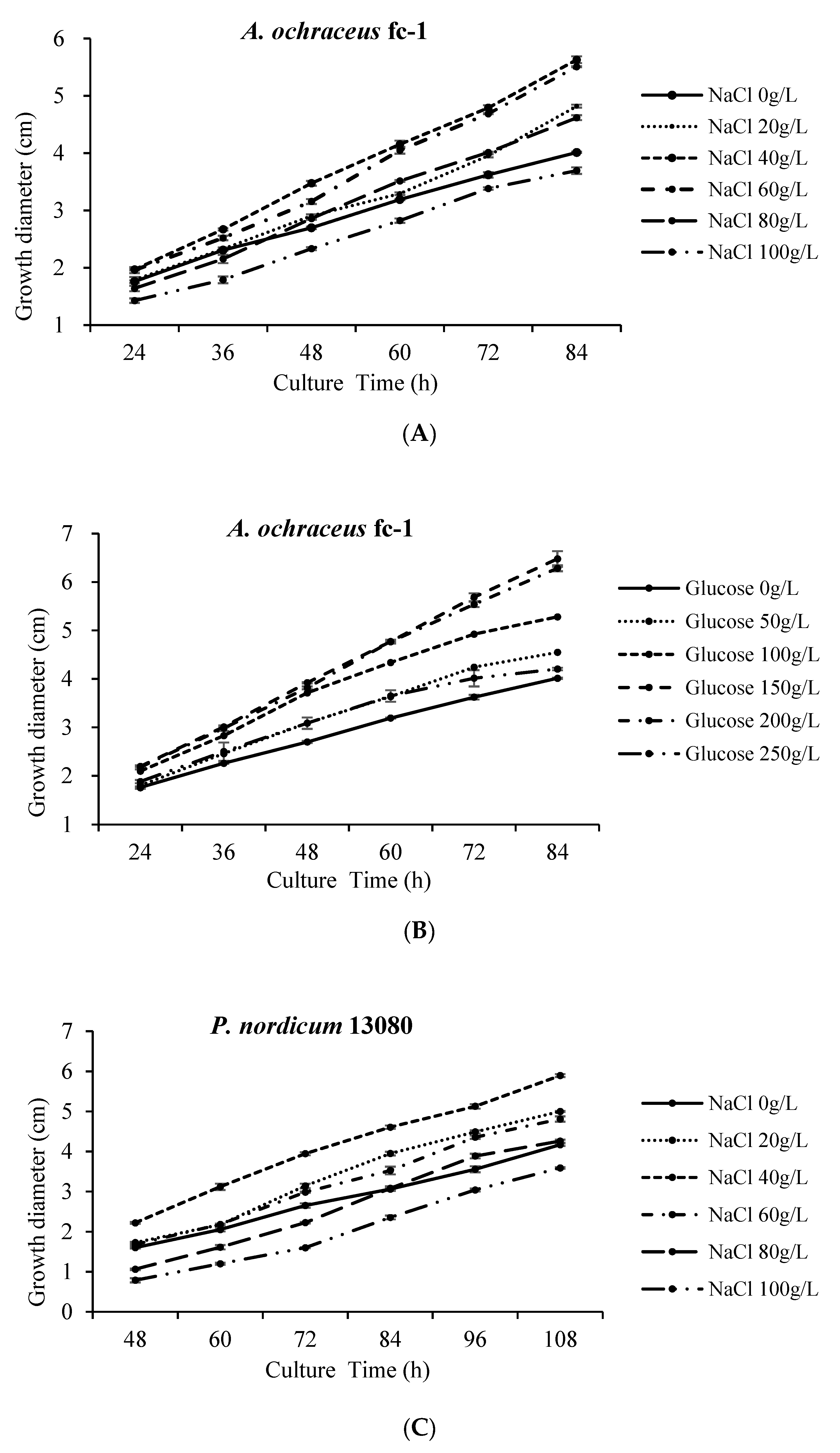
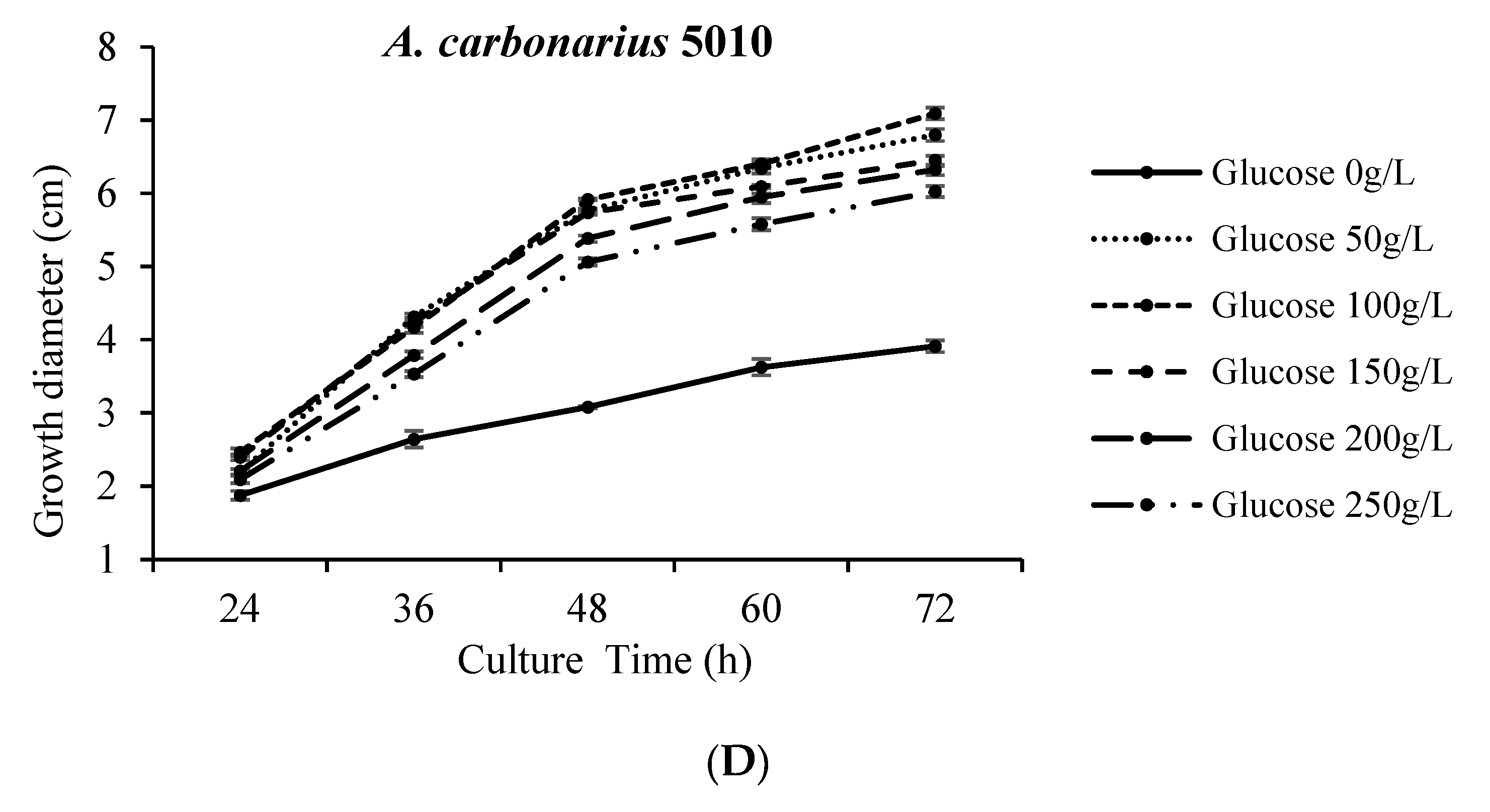
2.1. The Colony Morphology Are Affected by High Concentration of NaCl or Glucose
2.2. The Spore Production Are Affected by High Concentration of NaCl or Glucose
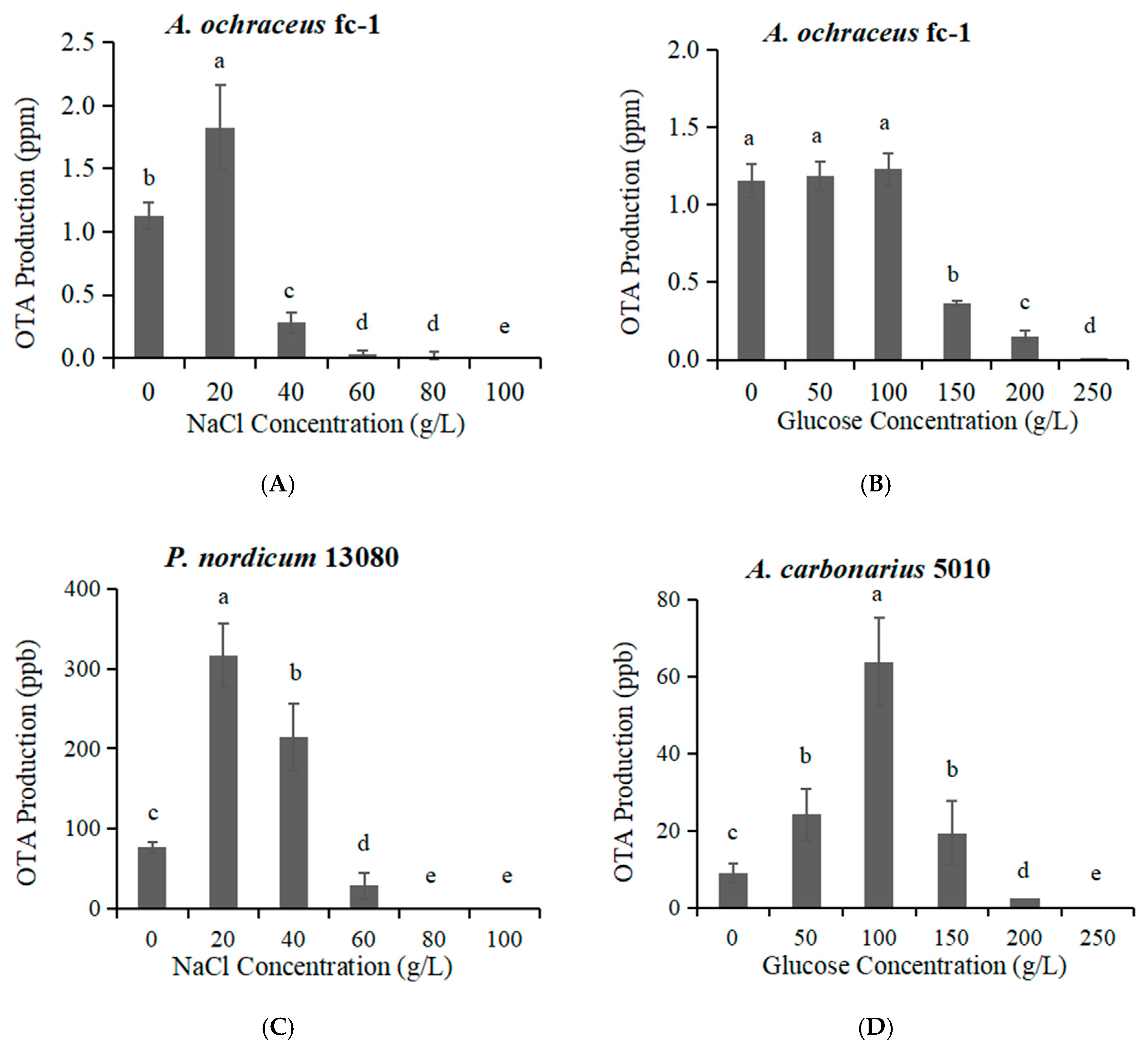
2.3. The OTA Production Effected by High Concentration of NaCl or Glucose
2.4. Relationship between Water Activity (Ionic Concentration) and OTA Production
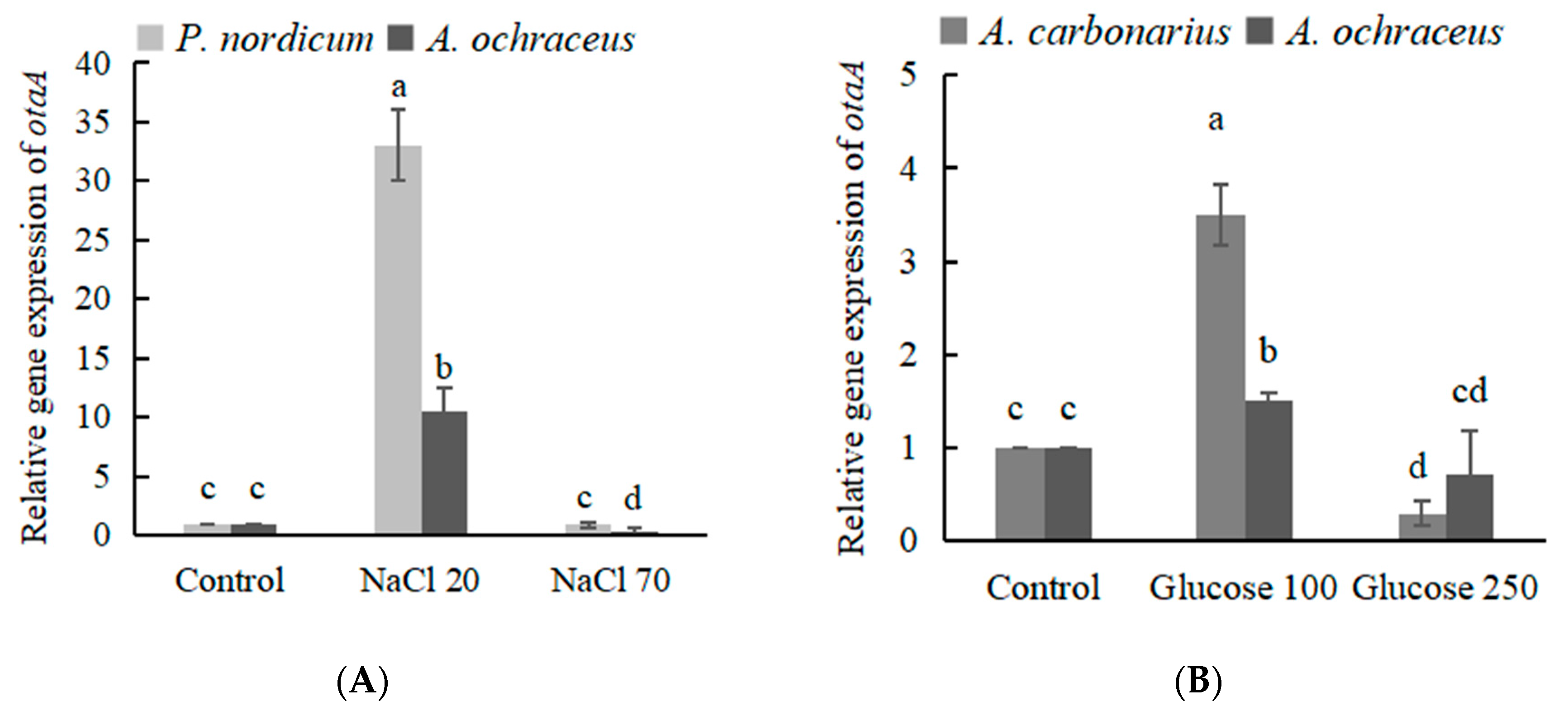
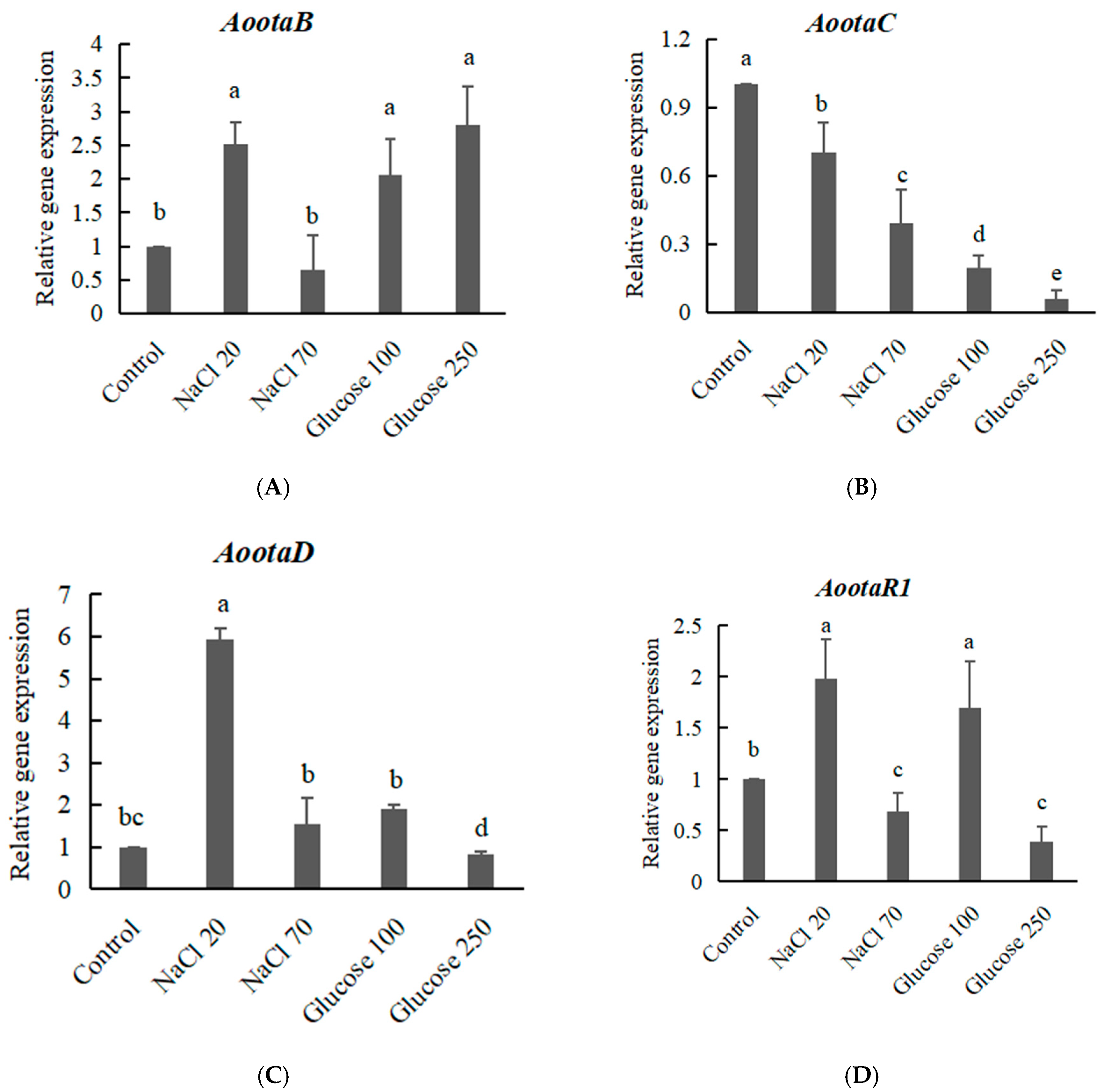
2.5. The Expression of OTA Biosynthetic Genes Effected by the Addition of NaCl or Glucose
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Strains and Media
5.2. Culture Media Preparation and Culture Conditions
5.3. Mycelial Growth Rate and Conidia Count
5.4. Extraction and Quantification of OTA
5.5. Gene Expression Studies
5.6. Data Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ostry, V.; Malir, F.; Ruprich, J. Producers and important dietary sources of ochratoxin A and citrinin. Toxins 2013, 5, 1574–1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, L.; Liu, F.; Wang, Q.; Selvaraj, J.N.; Xing, F.; Zhao, Y.; Liu, Y. Ochratoxin A producing fungi, biosynthetic pathway and regulatory mechanisms. Toxins 2016, 8, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, T.; Shen, X.L.; Chen, W.; Liao, X.; Yang, J.; Wang, Y.; Zou, Y.; Fang, C. Advances in research of nephrotoxicity and toxic antagonism of Ochratoxin A. Toxin Rev. 2017, 36, 39–44. [Google Scholar] [CrossRef]
- Northolt, M.D.; Van Egmond, H.P.; Paulsch, W.E. Ochratoxin A production by some fungal species in relation to water activity and temperature. J. Food Protect. 1979, 42, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.J.; Labernia, N.; Marín, S.; Sanchis, V.; Magan, N. Effect of water activity and temperature on growth and ochratoxin production by three strains of Aspergillus ochraceus on a barley extract medium and on barley grains. Int. J. Food Microbiol. 1998, 44, 133–140. [Google Scholar] [CrossRef]
- Somma, S.; Perrone, G.; Logrieco, A.F. Diversity of black Aspergilli and mycotoxin risks in grape, wine and dried vine fruits. Phytopathol. Mediterr. 2012, 51, 131–147. [Google Scholar]
- Cervini, C.; Gallo, A.; Piemontese, L.; Magistà, D.; Logrieco, A.F.; Ferrara, M.; Solfrizzo, M.; Perrone, G. Effects of temperature and water activity change on ecophysiology of ochratoxigenic Aspergillus carbonarius in field-simulating conditions. Int. J. Food Microbiol. 2020, 315, 108420. [Google Scholar] [CrossRef]
- Bejaoui, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Black aspergilli and ochratoxin A production in French vineyards. Int. J. Food Microbiol. 2006, 111 (Suppl. S1), 46–52. [Google Scholar] [CrossRef] [Green Version]
- Zadravec, M.; Vahčić, N.; Brnić, D.; Markov, K.; Frece, J.; Beck, R.; Lešić, T.; Pleadin, J. A study of surface moulds and mycotoxins in Croatian traditional dry-cured meat products. Int. J. Food Microbiol. 2020, 317, 108459. [Google Scholar] [CrossRef]
- Delgado, J.; da Cruz Cabral, L.; Rodríguez, A.; Rodríguez, M. Influence of ochratoxin A on adaptation of Penicillium nordicum on a NaCl-rich dry-cured ham-based medium. Int. J. Food Microbiol. 2018, 272, 22–28. [Google Scholar] [CrossRef]
- Jiang, C.; Shi, J.; Zhu, C. Fruit spoilage and ochratoxin a production by Aspergillus carbonarius in the berries of different grape cultivars. Food Control 2013, 30, 93–100. [Google Scholar] [CrossRef]
- Merla, C.; Andreoli, G.; Garino, C.; Vicari, N.; Tosi, G.; Guglielminetti, M.L.; Moretti, A.; Biancardi, A.; Arlorio, M.; Fabbi, M. Monitoring of ochratoxin A and ochratoxin-producing fungi in traditional salami manufactured in Northern Italy. Mycotoxin Res. 2018, 34, 107–116. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Wu, F.; Liu, F.; Wang, Q.; Zhang, X.; Selvaraj, J.N.; Zhao, Y.; Xing, F.; Yin, W.-B.; et al. A consensus ochratoxin A biosynthetic pathway: Insights from the genome sequence of Aspergillus ochraceus and a comparative genomic analysis. Appl. Environ. Microbiol. 2018, 84, e01009-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrara, M.; Perrone, G.; Gambacorta, L.; Epifani, F.; Solfrizzo, M.; Gallo, A. Identification of a halogenase involved in the biosynthesis of ochratoxin A in Aspergillus carbonarius. Appl. Environ. Microbiol. 2016, 82, 5631–5641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt-Heydt, M.; Graf, E.; Stoll, D.; Geisen, R. The biosynthesis of ochratoxin A by Penicillium as one mechanism for adaptation to NaCl rich foods. Food Microbiol. 2012, 29, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, M.; Magistà, D.; Lippolis, V.; Cervellieri, S.; Susca, A.; Perrone, G. Effect of Penicillium nordicum contamination rates on ochratoxin A accumulation in dry-cured salami. Food Control 2016, 67, 235–239. [Google Scholar] [CrossRef]
- Coton, M.; Coton, E.; Auffret, A.; Poirier, E.; Debaets, S.; Dantigny, P. Production and migration of ochratoxin A and citrinin in Comté cheese by an isolate of Penicillium verrucosum selected among Penicillium spp. mycotoxin producers in YES medium. Food Microbiol. 2019, 82, 551–559. [Google Scholar] [CrossRef]
- Zhang, X.; Li, J.; Zong, N.; Zhou, Z.; Ma, L. Ochratoxin A in dried vine fruits from Chinese markets. Food Addit. Contam. Part B 2014, 7, 157–161. [Google Scholar] [CrossRef]
- Zhang, X.; Li, J.; Cheng, Z.; Zhou, Z.; Ma, L. High-performance liquid chromatography-tandem mass spectrometry method for simultaneous detection of ochratoxin A and relative metabolites in Aspergillus species and dried vine fruits. Food Addit. Contam. Part A 2016, 33, 1355–1366. [Google Scholar]
- Altafini, A.; Fedrizzi, G.; Roncada, P. Occurrence of ochratoxin A in typical salami produced in different regions of Italy. Mycotoxin Res. 2018, 35, 141–148. [Google Scholar] [CrossRef]
- Geisen, R.; Touhami, N.; Schmidt-Heydt, M. Mycotoxins as adaptation factors to food related environments. Curr. Opin. Food Sci. 2017, 17, 1–8. [Google Scholar] [CrossRef]
- Geisen, R.; Stoll, D.; Schmidt-Heydt, M. Differences in the regulation of ochratoxin a by the HOG pathway in Penicillium and Aspergillus in response to high osmolar environments. Toxins 2013, 5, 1282–1298. [Google Scholar]
- Rodríguez, A.; Medina, Á.; Córdoba, J.J.; Magan, N. The influence of salt (NaCl) on ochratoxin A biosynthetic genes, growth and ochratoxin A production by three strains of Penicillium nordicum on a dry-cured ham-based medium. Int. J. Food Microbiol. 2014, 178, 113–119. [Google Scholar] [CrossRef]
- Rodríguez, A.; Capela, D.; Medina, Á.; Córdoba, J.J.; Magan, N. Relationship between ecophysiological factors, growth and ochratoxin A contamination of dry-cured sausage based matrices. Int. J. Food Microbiol. 2015, 194, 71–77. [Google Scholar] [CrossRef]
- Bernáldez, V.; Rodríguez, A.; Delgado, J.; Sánchez-Montero, L.; Córdoba, J.J. Gene expression analysis as a method to predict OTA accumulation in dry-cured meat products. Food Anal. Method 2018, 11, 2463–2471. [Google Scholar] [CrossRef]
- Ioannidis, A.-G.; Kogkaki, E.A.; Natskoulis, P.I.; Nychas, G.-J.E.; Panagou, E.Z. Modelling the influence of temperature, water activity and sodium metabisulphite on the growth and OTA production of Aspergillus carbonarius isolated from Greek wine grapes. Food Microbiol. 2015, 49, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Valero, A.; Marín, S.; Ramos, A.J.; Sanchis, V. Ochratoxin A-producing species in grapes and sun-dried grapes and their relation to ecophysiological factors. Lett. Appl. Microbiol. 2005, 41, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Fodil, S.; Delgado, J.; Varvaro, L.; Yaseen, T.; Rodríguez, A. Effect of potassium sorbate (E-202) and the antifungal PgAFP protein on Aspergillus carbonarius growth and ochratoxin A production in raisin simulating media. J. Sci. Food Agric. 2018, 98, 5785–5794. [Google Scholar] [CrossRef] [PubMed]
- Covarelli, L.; Beccari, G.; Marini, A.; Tosi, L. A review on the occurrence and control of ochratoxigenic fungal species and ochratoxin A in dehydrated grapes, non-fortified dessert wines and dried vine fruit in the Mediterranean area. Food Control 2012, 26, 347–356. [Google Scholar] [CrossRef]
- Lappa, I.K.; Kizis, D.; Panagou, E.Z. Monitoring the temporal expression of genes involved in ochratoxin a production of Aspergillus carbonarius under the influence of temperature and water activity. Toxins 2017, 9, 296. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, F.; Wang, L.; Wang, Q.; Selvaraj, J.N.; Zhao, Y.; Wang, Y.; Xing, F.; Liu, Y. pH-signaling transcription factor AopacC regulates ochratoxin A biosynthesis in Aspergillus ochraceus. J. Agric. Food Chem. 2018, 66, 4394–4401. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Knox, B.P.; Bruno, K.S.; Solfrizzo, M.; Baker, S.E.; Perrone, G. Identification and characterization of the polyketide synthase involved in ochratoxin A biosynthesis in Aspergillus carbonarius. Int. J. Food Microbiol. 2014, 179, 10–17. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Functions | P. nordicum | A. ochraceus | A. carbonarius | |||||
|---|---|---|---|---|---|---|---|---|---|
| NaCl (g/L) | NaCl (g/L) | Glc (g/L) | Glc (g/L) | ||||||
| 20 | 70 | 20 | 70 | 100 | 250 | 100 | 250 | ||
| otaA | type i iterative polyketide synthase (PKS) | + | N | + | − | + | N | + | − |
| otaB | nonribosomal peptide synthase (NRPS) | + | N | + | + | ||||
| otaC | cytochrome p450 monooxygenase | − | − | − | − | ||||
| otaD | halogenase | + | + | + | − | ||||
| otaR1 | bZIP transcription factor | + | − | + | − | ||||
| Primer Name | Sequence (5′ to 3′) | Strain |
|---|---|---|
| GADPH-F | TGCTCAAGTACGACAGCACC | Aspergillus ochraceus [13] |
| GADPH-R | CTCGGCGAAGAACTGAACCT | |
| AootaA-F | CGCCTCATCATCAATCCTT | |
| AootaA-R | CAACTCGGTCAAGCAGAT | |
| AootaB-F | ATACCACCAGAGCTCCAAA | |
| AootaB-R | GAGATGTTCGGTCTGTTCA | |
| AootaC-F | CTTAATACGGTGGTCTACGA | |
| AootaC-R | GAATGATAGGTCCGTATTTCT | |
| AootaD-F | CTATCCGGTGGTCTGTCAGC | |
| AootaD-R | TGAATGCATCGTCGAACCCA | |
| AootaR1-F | GCTTTCAAATCGAATGATTCC | |
| AootaR1-R | GATCGGTTGGAAGTGTAGAA | |
| β-tub F | GCCAGCGGTGACAAGTACGT | Penicillium nordicum [10] |
| β-tub R | TACCGGGCTCCAAATCGA | |
| PnotaA-F | CGCCGCTGCGGTTACT | |
| PnotaA-R | GGTAACAATCAACGCTCCCTCTT | |
| β-tub F | CAAACCGGCCAGTGTGGTA | Aspergillus carbonarius [32] |
| β-tub R | CGGAGGTGCCATTGTAAACA | |
| AcotaA-F | CGTGTCCGATACTGTCTGTGA | |
| AcotaA-R | GCATGGAGTCCTCAAGAACC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Yan, H.; Neng, J.; Gao, J.; Yang, B.; Liu, Y. The Influence of NaCl and Glucose Content on Growth and Ochratoxin A Production by Aspergillus ochraceus, Aspergillus carbonarius and Penicillium nordicum. Toxins 2020, 12, 515. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080515
Wang Y, Yan H, Neng J, Gao J, Yang B, Liu Y. The Influence of NaCl and Glucose Content on Growth and Ochratoxin A Production by Aspergillus ochraceus, Aspergillus carbonarius and Penicillium nordicum. Toxins. 2020; 12(8):515. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080515
Chicago/Turabian StyleWang, Yan, Hao Yan, Jing Neng, Jian Gao, Bolei Yang, and Yang Liu. 2020. "The Influence of NaCl and Glucose Content on Growth and Ochratoxin A Production by Aspergillus ochraceus, Aspergillus carbonarius and Penicillium nordicum" Toxins 12, no. 8: 515. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12080515