The Insecticidal Activity of Rhinella schneideri (Werner, 1894) Paratoid Secretion in Nauphoeta cinerea Cocroaches

 , , ,
, , ,
Abstract
:
1. Introduction
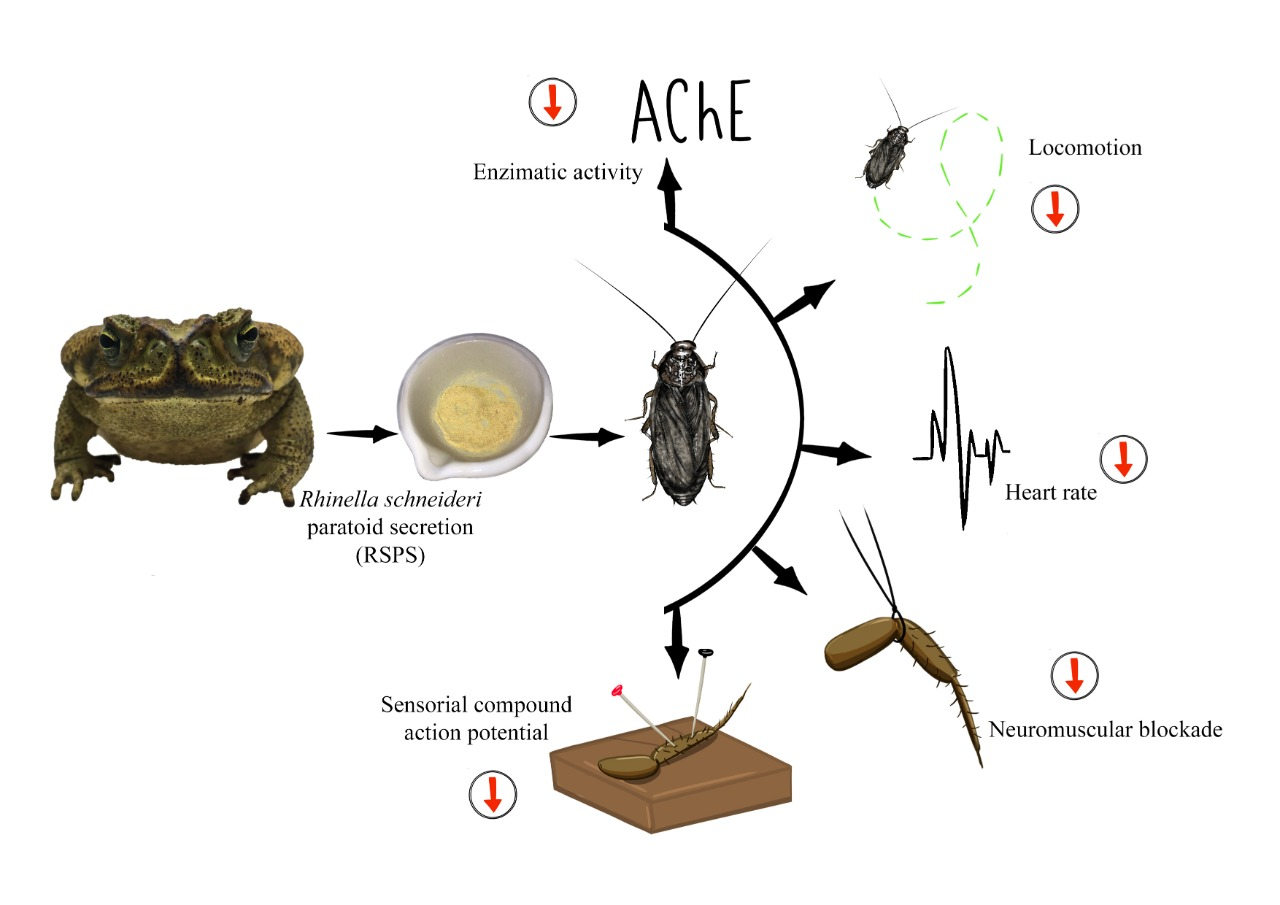
2. Results
2.1. High Resolution Mass Spectrometry
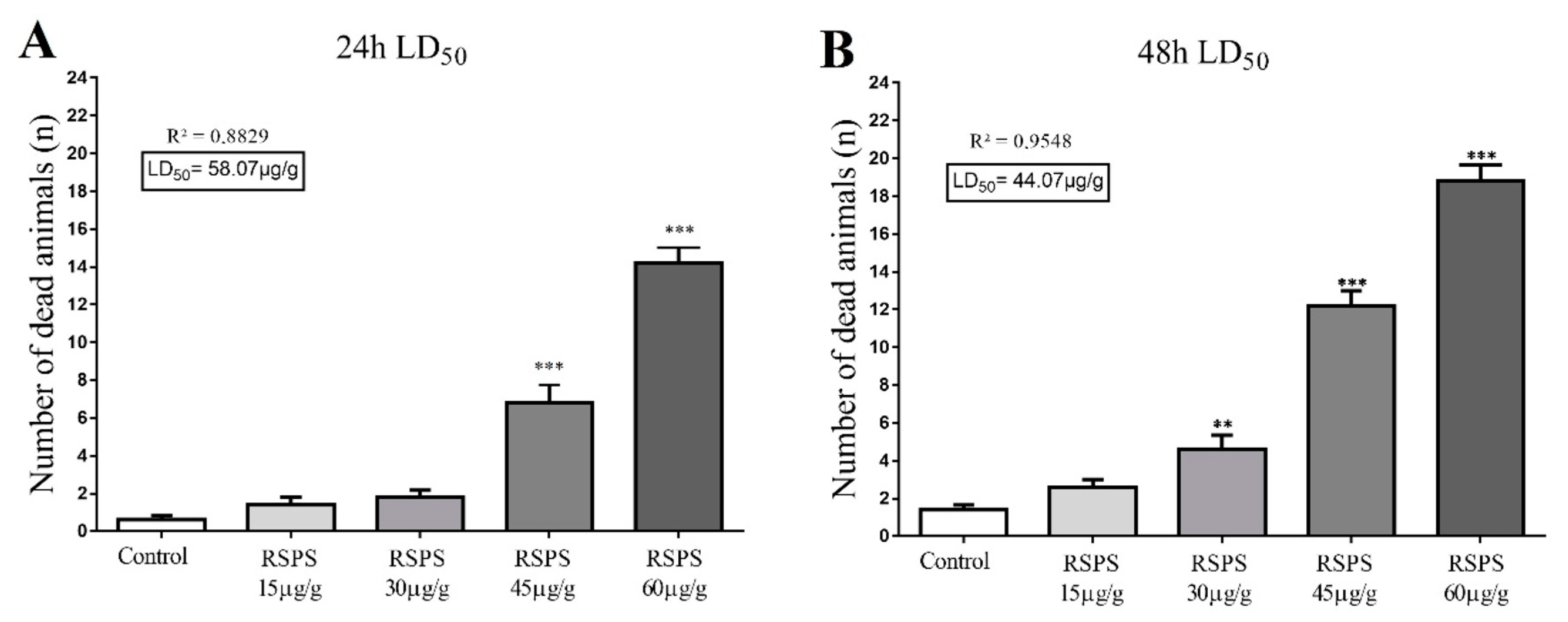
2.2. Lethality Dose Assay (LD50)
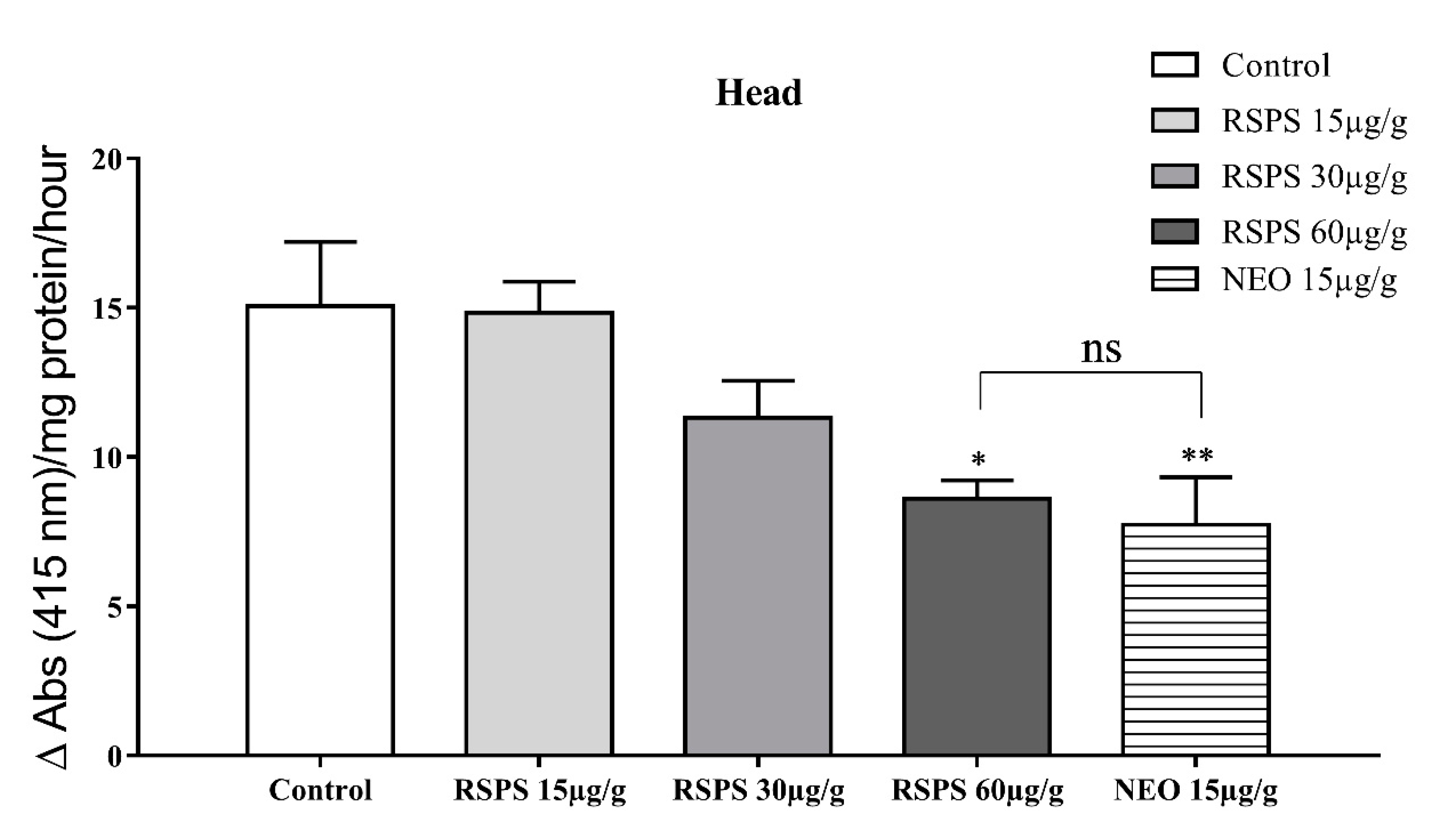
2.3. Acetylcholinesterase Activity
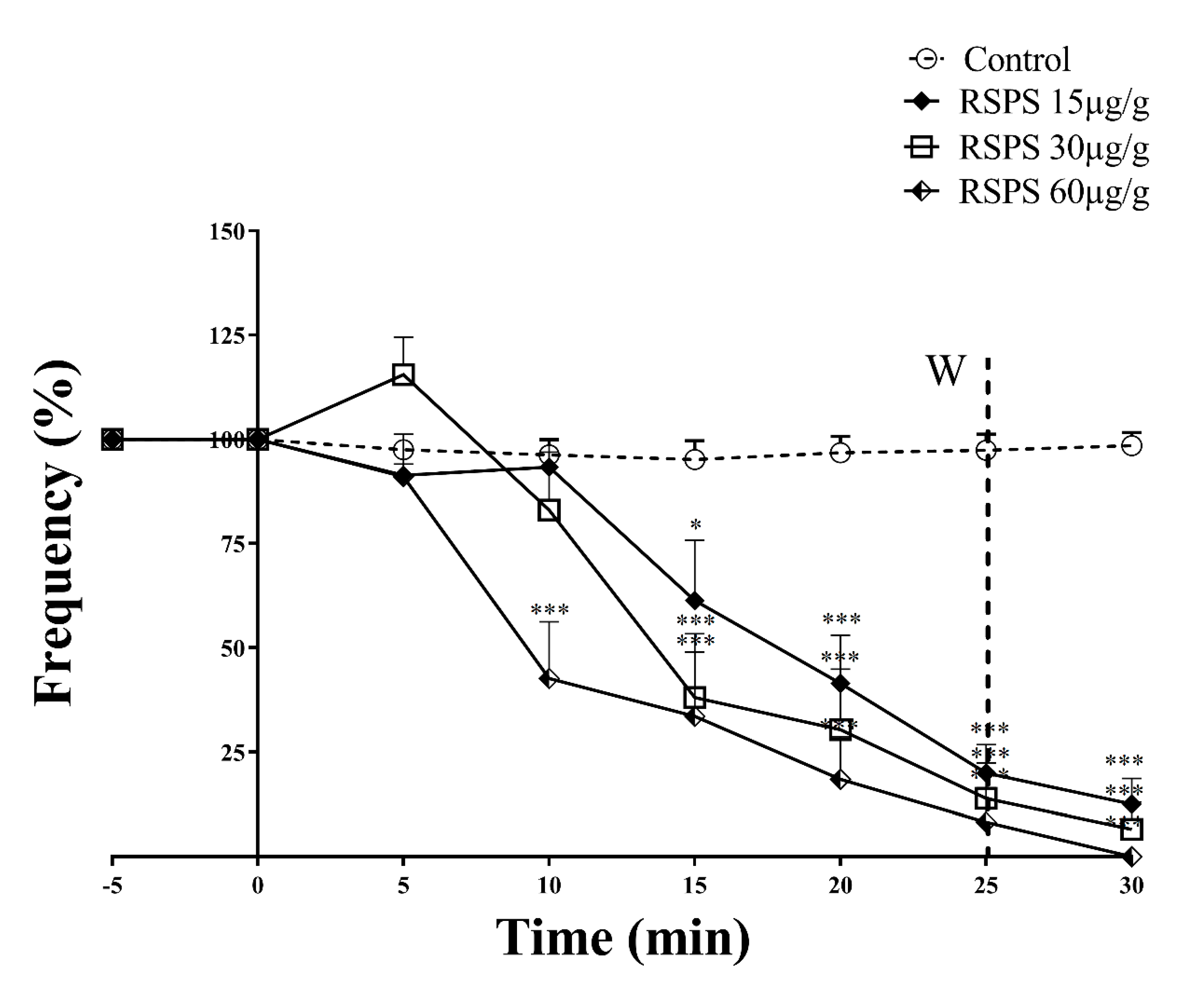
2.4. Locomotory Activity
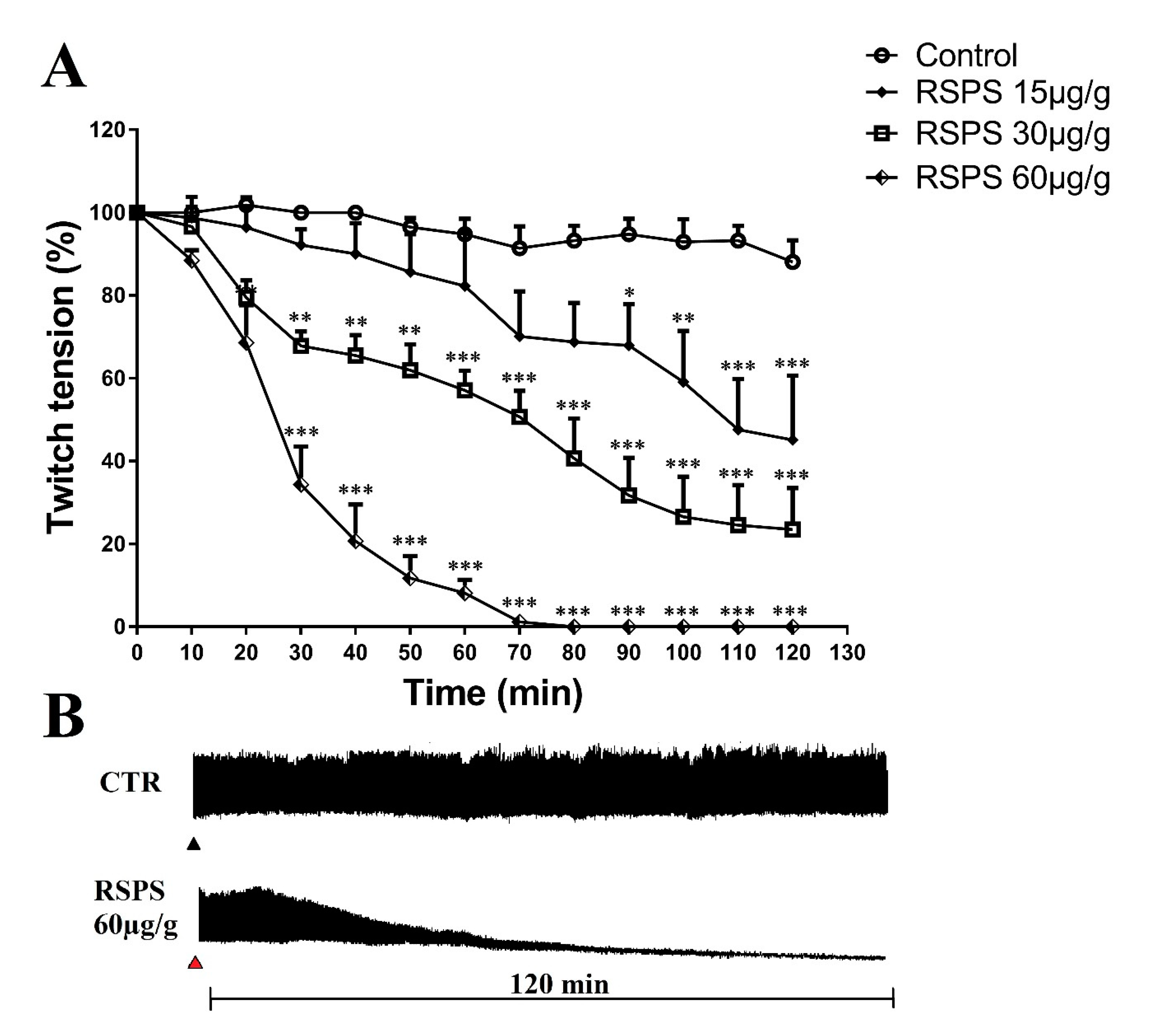
2.5. Effect of RSPS on Semi-Isolated Cockroach Heart Preparation
2.6. Effect of RSPS at Cockroach Metathoracic Coxal-Adductor Nerve-Muscle Preparation
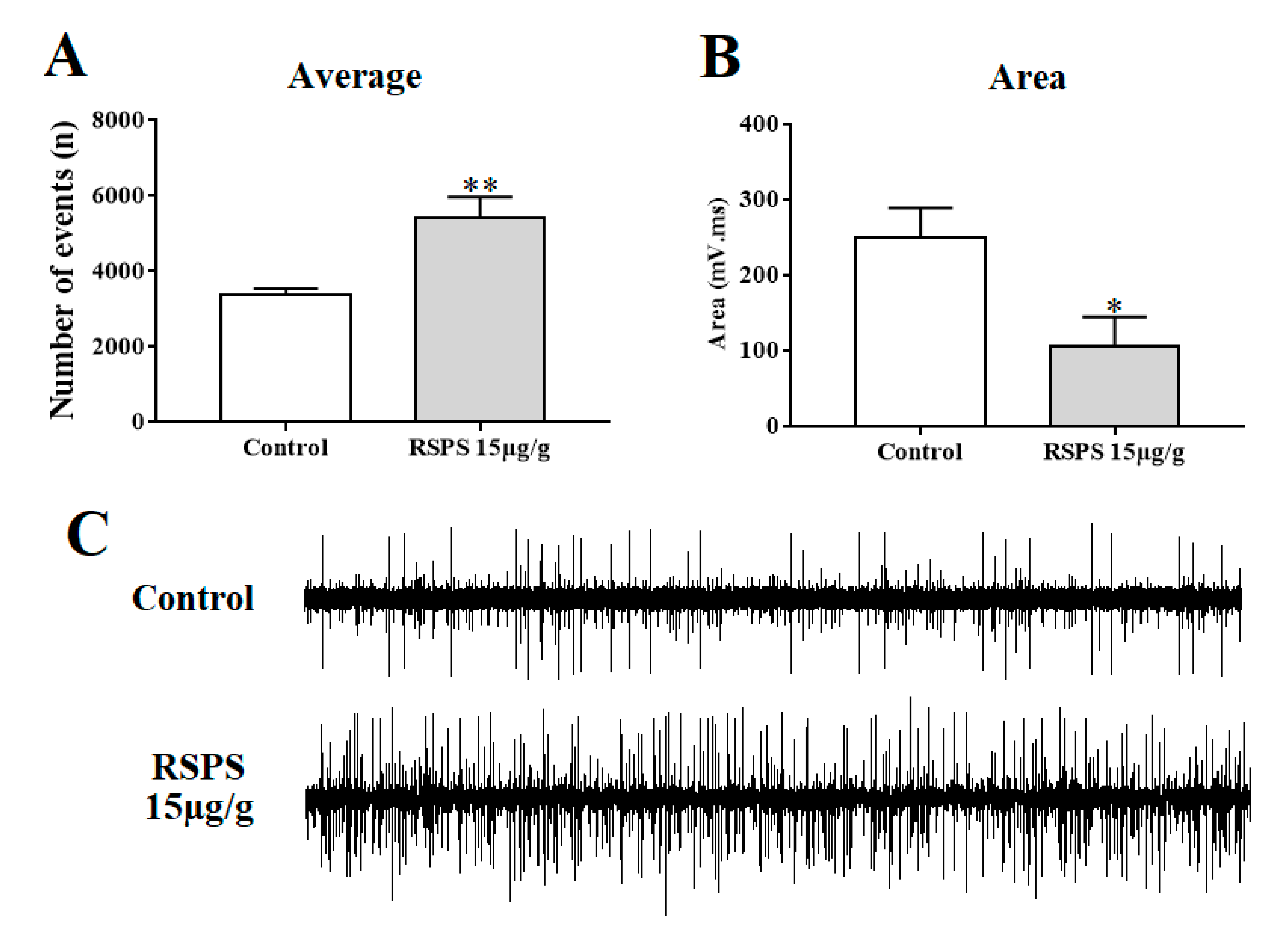
2.7. Recordings of Spontaneous Neural Compound Action Potentials (SNCAP)
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Experimental Animals
5.2. Rhinella Schneideri
5.3. Rhinella Schneideri Parotid Secretion (RSPS )
5.4. Biological Assays
5.5. Reagents and Solutions
5.6. Fractionation and Identification
5.6.1. Chemical fractionation
5.6.2. High Resolution Mass Spectrometry
5.7. Lethality Dose Assay (LD50)
5.8. Acetylcholinesterase Activity
5.9. Assay for Locomotory Activity
5.10. Semi-Isolated Cockroach Heart Preparation
5.11. Metathoracic Coxal-Adductor Nerve-Muscle Preparation
5.12. Recordings of Spontaneous Neural Compound Action Potentials (SNCAP)
5.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ngai, M.; McDowell, M.A. The search for novel insecticide targets in the post-genomics era, with a specific focus on G-protein coupled receptors. Mem. Inst. Oswaldo Cruz 2017, 112, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. A Global Brief on Vector-Borne Diseases; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Chen, Y.H.; Udalov, M.; Benkovskaya, G.; Lindström, L. Evolutionary Considerations in Potato Pest Management. Insect Pests Potato 2013, 19, 543–571. [Google Scholar]
- de Sousa, L.Q.; da Conceição Machado, K.; de Carvalho Oliveira, S.F.; da Silva Araújo, L.; Dos Santos Monção-Filho, E.; de Carvalho Melo-Cavalcante, A.A.; Vieira-Júnior, G.M.; Ferreira, P.M.P. Bufadienolides from amphibians: A promising source of anticancer prototypes for radical innovation, apoptosis triggering and Na+/K+-ATPase inhibition. Toxicon 2017, 127, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Briso, A.L.F.; Oda, F.H.; Campos, V.A. Rhinella schneideri, Cururu Toad. IUCN Red List Threat. Species 2013, 44, 130–131. [Google Scholar]
- Barravieira, B. Venenos animais: Uma visão integrada. Rev. Inst. Med. Trop. São Paulo 1994, 36, 300. [Google Scholar] [CrossRef]
- Chen, K.K.; Chen, A.L. notes on the poisonous secretions of twelve species of toads. J. Pharmacol. Exp. Ther. 1933, 47, 281–293. [Google Scholar]
- Balazs, T.; Hanig, J.P.; Herman, E.H. Toxic responses of the cardiovascular system. In Casarett Doull’s Toxicology: The Basic Science of Poisons; Klaassen, C.D., Amdur, M.O., Doull, J., Eds.; Macmillan Publishing Co. Inc.: New York, NY, USA, 1986; pp. 387–411. [Google Scholar]
- Duellman, W.E.; Trueb, L. Biology of Amphibians New York McGraw-Hill. Copeia 1986, 1986, 549–553. [Google Scholar]
- Brownlee, A.A.; Johnson, P.; Mills, I.H. Actions of bufalin and cinobufotalin, two bufadienolides respectively more active and less active than ouabain, on ouabain binding and 86Rb uptake by human erythrocytes. Clin. Sci. 1990, 78, 169–174. [Google Scholar] [CrossRef]
- Sciani, J.M.; Angeli, C.B.; Antoniazzi, M.M.; Jared, C.; Pimenta, D.C. Differences and similarities among parotoid macrogland secretions in South American toads: A preliminary biochemical delineation. Sci. World J. 2013, 2013, 937407. [Google Scholar] [CrossRef] [Green Version]
- Rostelato-Ferreira, S.; Dal Belo, C.A.; da Cruz-Höfling, M.A.; Hyslop, S.; Rodrigues-Simioni, L. Presynaptic effect of a methanolic extract of toad (Rhinella schneideri) poison in avian neuromuscular preparation. J. Venom Res. 2011, 2, 32–36. [Google Scholar]
- Shibao, P.Y.T.; Anjolette, F.A.P.; Lopes, N.P.; Arantes, E.C. First serine protease inhibitor isolated from Rhinella schneideri poison. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Yuan, B.; Takagi, N.; Wang, H.; Zhou, Y.; Si, N.; Yang, J.; Wei, X.; Zhao, H.; Bian, B. Comparative analysis of hydrophilic ingredients in toad skin and toad venom using the UHPLC-HR-MS/MS and UPLC-QqQ-MS/MS methods together with the anti-inflammatory evaluation of indolealkylamines. Molecules 2019, 24, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmeda-Hirschmann, G.; Quispe, C.; Arana, G.V.; Theoduloz, C.; Urra, F.A.; Cárdenas, C. Antiproliferative activity and chemical composition of the venom from the Amazonian toad Rhinella marina (Anura: Bufonidae). Toxicon 2016, 121, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Han, L.; Luo, M.; Bian, B.; Guan, M.; Yang, H.; Han, C.; Li, N.; Li, T.; Li, S. Multi-component identification and target cell-based screening of potential bioactive compounds in toad venom by UPLC coupled with high-resolution LTQ-Orbitrap MS and high-sensitivity Qtrap MS. Anal. Bioanal. Chem. 2018, 410, 4419–4435. [Google Scholar] [CrossRef] [PubMed]
- Schmeda-Hirschmann, G.; Gomez, C.V.; de Arias, A.R.; Burgos-Edwards, A.; Alfonso, J.; Rolon, M.; Brusquetti, F.; Netto, F.; Urra, F.A.; Cárdenas, C. The Paraguayan Rhinella toad venom: Implications in the traditional medicine and proliferation of breast cancer cells. J. Ethnopharmacol. 2017, 199, 106–118. [Google Scholar] [CrossRef]
- Matsukawa, M.; Akizawa, T.; Ohigashi, M.; Morris, J.F.; BUTLER, J.; Yoshioka, M. A novel bufadienolide, marinosin, in the skin of the giant toad, Bufo marinus. Chem. Pharm. Bull. 1997, 45, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Rattan, R.S. Mechanism of action of insecticidal secondary metabolites of plant origin. Crop Prot. 2010, 29, 913–920. [Google Scholar] [CrossRef]
- Costa, L.G. Current issues in organophosphate toxicology. Clin. Chim. Acta 2006, 366, 1–13. [Google Scholar] [CrossRef]
- Dahm, P.A. Toxic effects produced in insects by organophosphorus compounds. Bull. World Health Organ. 1971, 44, 215–219. [Google Scholar]
- Ritzmann, R.; Harley, C.; Daltorio, K.; Tietz, B.; Pollack, A.; Bender, J.; Guo, P.; Horomanski, A.; Kathman, N.; Nieuwoudt, C.; et al. Deciding Which Way to Go: How Do Insects Alter Movements to Negotiate Barriers? Front. Neurosci. 2012, 6, 97. [Google Scholar] [CrossRef] [Green Version]
- Endler, J.A. Some general comments on the evolution and design of animal communication systems. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1993, 340, 215–225. [Google Scholar]
- Adedara, I.A.; Rosemberg, D.B.; de Souza, D.; Farombi, E.O.; Aschner, M.; Souza, D.O.; Rocha, J.B.T. Neurobehavioral and biochemical changes in Nauphoeta cinerea following dietary exposure to chlorpyrifos. Pestic. Biochem. Physiol. 2016, 130, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Leal, A.P.; Oliveira, R.S.; Perin, A.P.A.; Borges, B.T.; de Brum Vieira, P.; dos Santos, T.G.; Vinadé, L.; Valsecchi, C.; Dal Belo, C.A. Entomotoxic activity of Rhinella icterica (Spix, 1824) toad skin secretion in Nauphoeta cinerea cockroaches: An octopamine-like modulation. Pestic. Biochem. Physiol. 2018, 148, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Tuthill, J.C.; Wilson, R.I. Mechanosensation and adaptive motor control in insects. Curr. Biol. 2016, 26, R1022–R1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stu, G.D.; de Freitas, T.C.; de Avila Heberle, M.; de Assis, D.R.; Vinadé, L.; Pereira, A.B.; Franco, J.L.; Dal Belo, C.A. Modulation of dopaminergic neurotransmission induced by sublethal doses of the organophosphate trichlorfon in cockroaches. Ecotoxicol. Environ. Saf. 2014, 109, 56–62. [Google Scholar]
- Farooqui, T. Review of octopamine in insect nervous systems. Open Access Insect Physiol. 2012, 4, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Rostelato-Ferreira, S.; Dal Belo, C.A.; Leite, G.B.; Hyslop, S.; Rodrigues-Simioni, L. Presynaptic neuromuscular action of a methanolic extract from the venom of Rhinella schneideri toad. J. Venom. Anim. Toxins Incl. Trop. Dis. 2014, 20, 30. [Google Scholar] [CrossRef] [Green Version]
- Gerschenfeld, H.M. Chemical transmission in invertebrate central nervous systems and neuromuscular junctions. Physiol. Rev. 1973, 53, 1–119. [Google Scholar] [CrossRef]
- Langeloh, H.; Wasser, H.; Richter, N.; Bicker, G.; Stern, M. Neuromuscular transmitter candidates of a centipede (Lithobius forficatus, Chilopoda). Front. Zool. 2018, 15, 28. [Google Scholar] [CrossRef] [Green Version]
- Carrazoni, T.; de Avila Heberle, M.; Perin, A.P.A.; Zanatta, A.P.; Rodrigues, P.V.; dos Santos, F.D.M.; de Almeida, C.G.M.; Vaz Breda, R.; dos Santos, D.S.; Pinto, P.M.; et al. Central and peripheral neurotoxicity induced by the Jack Bean Urease (JBU) in Nauphoeta cinerea cockroaches. Toxicology 2016, 368–369, 162–171. [Google Scholar] [CrossRef]
- Libersat, F.; Pflueger, H.-J. Monoamines and the Orchestration of Behavior. Bioscience 2004, 54, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Weisel-Eichler, A.; Haspel, G.; Libersat, F. Venom of a parasitoid wasp induces prolonged grooming in the cockroach. J. Exp. Biol. 1999, 202, 957–964. [Google Scholar] [PubMed]
- Hoyle, G.; Barker, D.L. Synthesis of octopamine by insect dorsal median unpaired neurons. J. Exp. Zool. Part A Ecol. Genet. Physiol. 1975, 193, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Biewener, A.; Patek, S. Animal Locomotion; Oxford University Press: Oxford, UK, 2018; ISBN 0191060852. [Google Scholar]
- Pamnani, M.B.; Chen, S.; Bryant, H.J.; Schooley, J.F., Jr.; Eliades, D.C.; Yuan, C.M.; Haddy, F.J. Effects of three sodium-potassium adenosine triphosphatase inhibitors. Hypertension 1991, 18, 316–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirai, Y.; Morishita, S.; Ito, C.; Sakanashi, M. Effects of bufadienolides and some kinds of cardiotonics on guinea pig hearts. Nihon Yakurigaku Zasshi 1992, 100, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.; Manunta, P.; Chen, S.; Hamlyn, J.M.; Haddy, F.J.; Pamnani, M.B. Role of ouabain-like factors in hypertension: Effects of ouabain and certain endogenous ouabain-like factors in hypertension. J. Cardiovasc. Pharmacol. 1993, 22, S10–S12. [Google Scholar] [CrossRef]
- Bagrov, A.Y.; Roukoyatkina, N.I.; Pinaev, A.G.; Dmitrieva, R.I.; Fedorova, O. V Effects of two endogenous Na+, K+-ATPase inhibitors, marinobufagenin and ouabain, on isolated rat aorta. Eur. J. Pharmacol. 1995, 274, 151–158. [Google Scholar] [CrossRef]
- Matsukawa, M.; Akizawa, T.; Morris, J.F.; BUTLER, J.; YOSHIOKA, M. Marinoic acid, a novel bufadienolide-related substance in the skin of the giant toad, Bufo marinus. Chem. Pharm. Bull. 1996, 44, 255–257. [Google Scholar] [CrossRef] [Green Version]
- Sokolove, P.G.; Cooke, I.M. Inhibition of impulse activity in a sensory neuron by an electrogenic pump. J. Gen. Physiol. 1971, 57, 125–163. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, R.S.; Borges, B.T.; Leal, A.P.; Lailowski, M.M.; de Castro Figueiredo Bordon, K.; de Souza, V.Q.; Vinadé, L.; dos Santos, T.G.; Hyslop, S.; Moura, S. Chemical and Pharmacological Screening of Rhinella icterica (Spix 1824) Toad Parotoid Secretion in Avian Preparations. Toxins 2020, 12, 396. [Google Scholar] [CrossRef]
- Kagabu, S.; Murase, Y.; Imai, R.; Ito, N.; Nishimura, K. Effect of substituents at the 5-position of the pyridine ring of imidacloprid on insecticidal activity against Periplaneta americana. Pest Manag. Sci. 2007, 63, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Carrazoni, T.; de Brum Vieira, P.; da Silva, P.A. Mechanism of the Entomotoxic Activity Induced by Araucaria Angustifolia Methanolic Extract in Nauphoeta Cinerea Lobster Cockroaches. J. Bot. Res. 2017, 1, 38–49. [Google Scholar]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–90. [Google Scholar] [CrossRef]
- Rodríguez, V.; Mori, B.; Dörr, F.A.; Dal Belo, C.A.; Colepicolo, P.; Pinto, E. Effects of a cyanobacterial extract containing-anatoxin-a (s) on the cardiac rhythm of Leurolestes circunvagans. Rev. Bras. Farmacogn. 2012, 22, 775–781. [Google Scholar] [CrossRef] [Green Version]
- Martinelli, A.H.S.; Kappaun, K.; Ligabue-Braun, R.; Defferrari, M.S.; Piovesan, A.R.; Staniscuaski, F.; Demartini, D.R.; Dal Belo, C.A.; Almeida, C.G.M.; Follmer, C.; et al. Structure-function studies on jaburetox, a recombinant insecticidal peptide derived from jack bean (Canavalia ensiformis) urease. Biochim. Biophys. Acta 2014, 1840, 935–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Precursor Ion m/z | Extract | Identification | Elem. Comp. | Diff Ppm | Comp. Type | Ref. |
|---|---|---|---|---|---|---|---|
| Extracts analysis in positive mode ESI (+) | |||||||
| 1 | 191.1173 | SCH Aq | n-methyl-5-hydroxy-tryptamine | C11H14N2O | 5.76 | Alkaloid | [14] |
| 2 | 387.2523 | SCH Aq | 3,14-dihydroxybufa-20,22-dienolide (Bufalin) | C24H34O4 | 3.09 | Steroid | [15] |
| 3 | 401.2321 401.2325 401.2322 | SCH Aq SCH SCH Et | 14,15-epoxy-3,5-dihydroxybufa-20,22-dienolide (marinobufagin) | C24H32O5 | 1.74 0.75 1.49 | Steroid | [15] |
| 4 | 713.4089 | SCH Aq | 3-(n-suberoyl argininyl) marinobufagin; (Mari- nobufotoxin) | C36H53N4O9 | 5.05 | Steroid | [15,16,17] |
| Extracts analysis in negative mode ESI (-) | |||||||
| 5 | 445.2220 445.2269 | SCH Aq SCH Et | 11,19-epoxy-19-methoxy-Telocinobufagin (Marinosin) | C25H34O7 | 1.34 9.65 | Steroid | [18] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leal, A.; Karnopp, E.; Barreto, Y.C.; Oliveira, R.S.; Rosa, M.E.; Borges, B.T.; Goulart, F.L.; Souza, V.Q.d.; Laikowski, M.M.; Moura, S.; et al. The Insecticidal Activity of Rhinella schneideri (Werner, 1894) Paratoid Secretion in Nauphoeta cinerea Cocroaches. Toxins 2020, 12, 630. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12100630
Leal A, Karnopp E, Barreto YC, Oliveira RS, Rosa ME, Borges BT, Goulart FL, Souza VQd, Laikowski MM, Moura S, et al. The Insecticidal Activity of Rhinella schneideri (Werner, 1894) Paratoid Secretion in Nauphoeta cinerea Cocroaches. Toxins. 2020; 12(10):630. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12100630
Chicago/Turabian StyleLeal, Allan, Etiely Karnopp, Yuri Correia Barreto, Raquel Soares Oliveira, Maria Eduarda Rosa, Bruna Trindade Borges, Flávia Luana Goulart, Velci Queiróz de Souza, Manuela Merlin Laikowski, Sidnei Moura, and et al. 2020. "The Insecticidal Activity of Rhinella schneideri (Werner, 1894) Paratoid Secretion in Nauphoeta cinerea Cocroaches" Toxins 12, no. 10: 630. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12100630