New Insights in Saccharomyces cerevisiae Response to the Cyanotoxin Microcystin-LR, Revealed by Proteomics and Gene Expression

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Saccharomyces cerevisiae Cells Viability When Exposed to Different Concentrations of MC-LR
2.2. Proteomic Analysis
2.2.1. Protein Expression
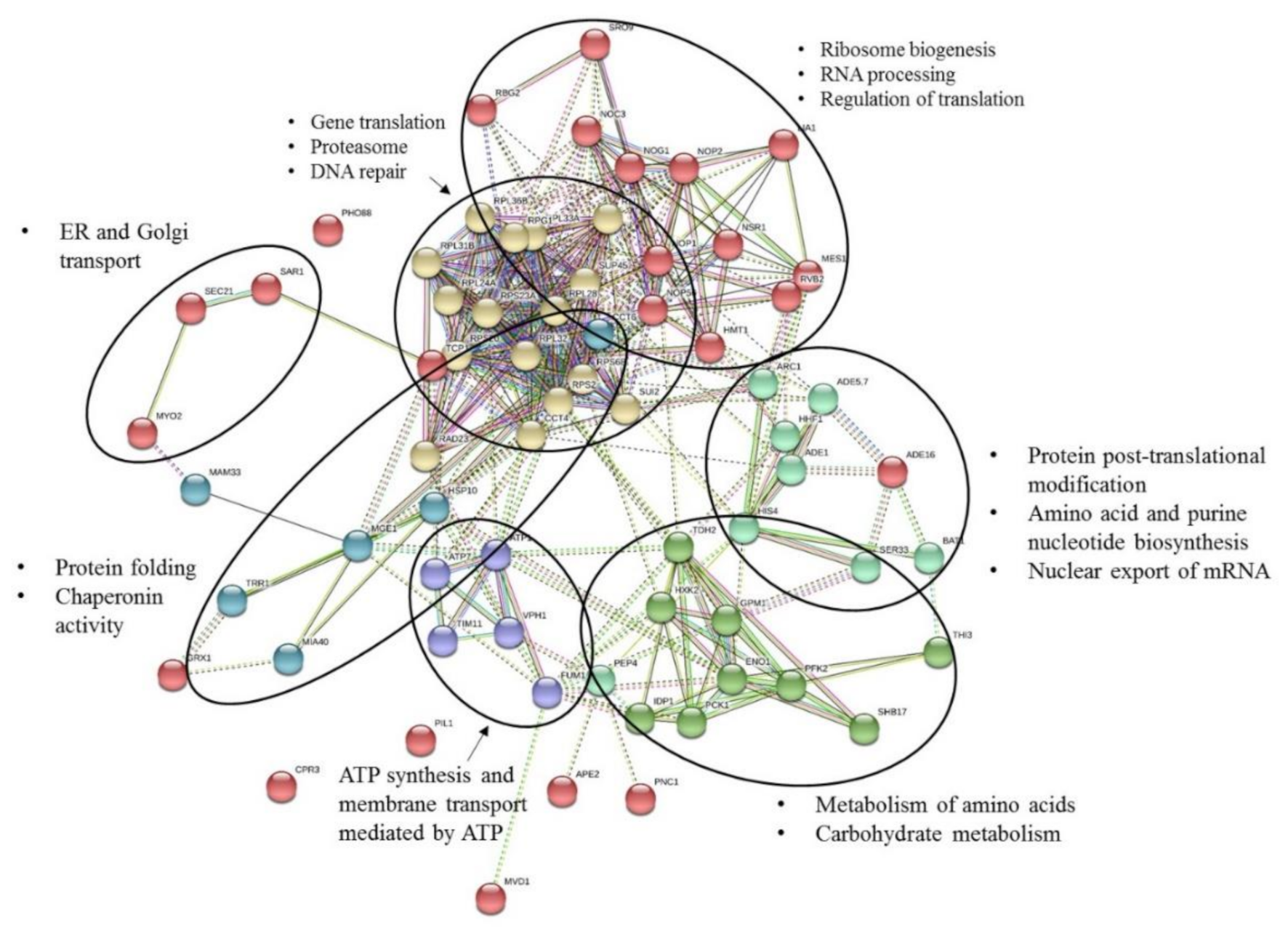
2.2.2. Protein Functional Analysis
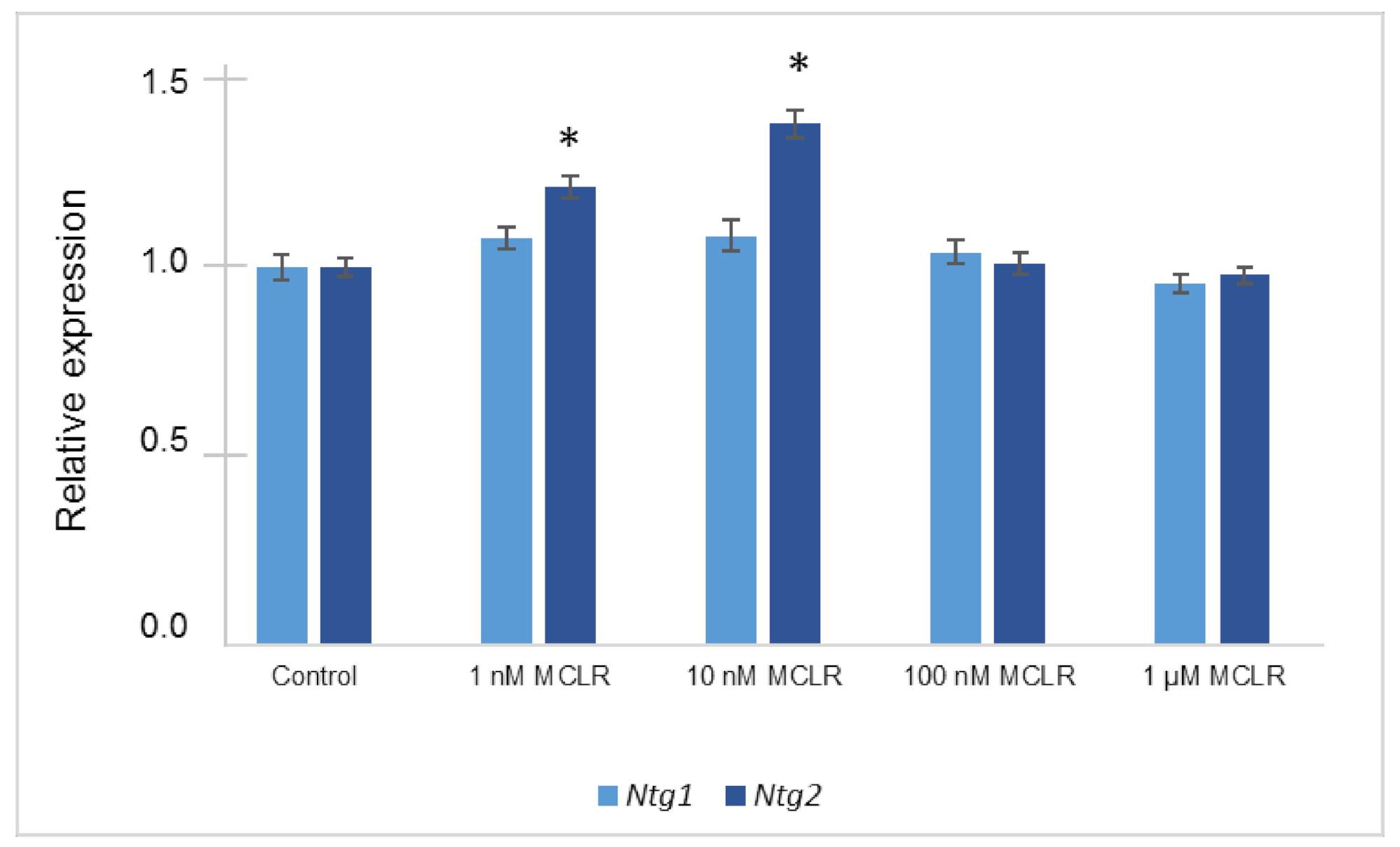
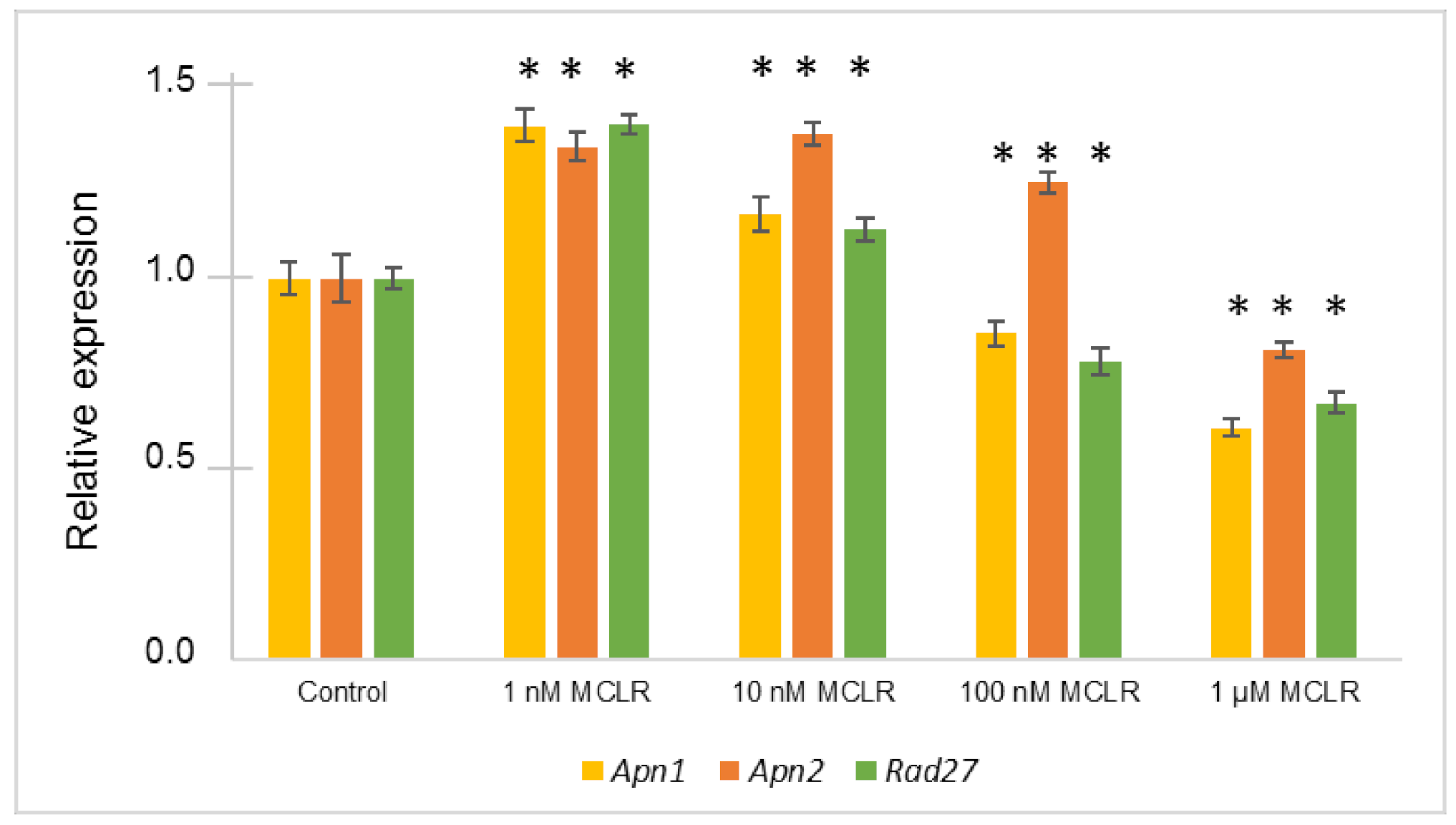
2.3. Relative Gene Expression Alterations of BER Genes in Saccharomyces cerevisiae Exposed to Different MC-LR Concentrations
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Saccharomyces cerevisiae Cultures
5.2. Analysis of Saccharomyces cerevisiae Viability When Exposed to Different Concentrations of MC-LR
5.3. Proteomic Analysis
5.3.1. Protein Extraction and Sample Preparation
5.3.2. LC-MS/MS and Protein Identification
5.4. Real-Time RT-qPCR Assays
5.4.1. Saccharomyces cerevisiae Studied Genes and Primers Design
5.4.2. Saccharomyces cerevisiae RNA Extraction and Purification
5.4.3. Evaluation of RT-qPCR Parameters
5.5. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Catherine, A.; Bernard, C.; Spoof, L.; Bruno, M. Microcystins and Nodularins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis, 1st ed.; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 109–126. [Google Scholar]
- van Apeldoorn, M.E.; van Egmond, H.P.; Speijers, G.J.; Bakker, G.J. Toxins of cyanobacteria. Mol. Nutr. Food Res. 2007, 51, 7–60. [Google Scholar] [CrossRef]
- Campos, A.; Vasconcelos, V. Molecular Mechanisms of Microcystin Toxicity in Animal Cells. Int. J. Mol. Sci. 2010, 11, 268–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valério, E.; Vasconcelos, V.; Campos, A. new insights on the mode of action of microcystins in animal cells—A Review. Mini Rev. Med. Chem. 2016, 16, 1032–1041. [Google Scholar] [CrossRef] [PubMed]
- Welten, R.D.; Meneely, J.P.; Elliott, C.T. A Comparative review of the effect of microcystin-LR on the proteome. Expo Health 2020, 12, 111–129. [Google Scholar] [CrossRef] [Green Version]
- McLellan, N.L.; Manderville, R.A. Toxic mechanisms of microcystins in mammals. Toxicol. Res. (Camb.) 2017, 6, 391–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Tu, W.; Xu, J. Mechanisms of microcystin-LR-induced cytoskeletal disruption in animal cells. Toxicon 2015, 101, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Shan, Z.; Xu, W.; Wang, X.; Zhou, J.; Kong, D.; Xu, J. Microcystin-LR induced reactive oxygen species mediate cytoskeletal disruption and apoptosis of hepatocytes in Cyprinus carpio L. PloS ONE 2013, 8, e84768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puerto, M.; Pichardo, S.; Jos, A.; Prieto, A.I.; Sevilla, E.; Frias, J.E.; Camean, A.M. Differential oxidative stress responses to pure Microcystin-LR and Microcystin-containing and non-containing cyanobacterial crude extracts on Caco-2 cells. Toxicon 2010, 55, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Zong, W.; Zhang, S.; Wang, Q.; Teng, Y.; Liu, Y.; Du, Y. Evaluation of the direct and indirect regulation pathways of glutathione target to the hepatotoxicity of Microcystin-LR. BioMed. Res. Int. 2018, 5672637. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.X.; Nam Ong, C. Role of oxidative stress and mitochondrial changes in cyanobacteria-induced apoptosis and hepatotoxicity. FEMS Microbiol. Lett. 2003, 220, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Lu, G.; Chen, G.; Huang, B.; Zhang, X.; Shen, K.; Wu, S. Microcystin-LR induces apoptosis via NF-κB/iNOS pathway in INS-1 cells. Int. J. Mol. Sci. 2011, 12, 4722–4734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Huang, P.; Liu, Y.; Du, H.; Wang, X.; Wang, M.; Wang, Y.; Hei, T.K.; Wu, L.; Xu, A. Role of nitric oxide in the genotoxic response to chronic microcystin-LR exposure in human–hamster hybrid cells. J. Environ. Sci. 2015, 29, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Huang, M.; Xu, P.; Wang, M.; Ye, S.; Wang, Q.; Zeng, S.; Chen, X.; Gao, W.; Chen, J.; et al. Microcystins-LR induced apoptosis via S-nitrosylation of GAPDH in colorectal cancer cells. Ecotoxicol. Environ. Saf. 2020, 190, 110096. [Google Scholar] [CrossRef] [PubMed]
- Valério, E.; Vilares, A.; Campos, A.; Pereira, P.; Vasconcelos, V. Effects of microcystin-LR on Saccharomyces cerevisiae growth, oxidative stress and apoptosis. Toxicon 2014, 90, 191–198. [Google Scholar] [CrossRef]
- Menezes, C.; Valério, E.; Dias, E. The kidney Vero-E6 cell line: A suitable model to study the toxicity of microcystins. In New Insights into Toxicity and Drug Testing; Gowder, S., Ed.; InTech.: London, UK, 2013; ISBN 978-953-51-0946-4. [Google Scholar]
- Valério, E.; Campos, A.; Osório, H.; Vasconcelos, V. Proteomic and Real-Time PCR analyses of Saccharomyces cerevisiae VL3 exposed to microcystin-LR reveals a set of protein alterations transversal to several eukaryotic models. Toxicon 2016, 112, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Sun, M.; Tang, Y.; Chen, Y.; Zhu, C.; Yang, Y.; Wang, C.; Yu, G.; Tang, Z. Responses of the proteome in testis of mice exposed chronically to environmentally relevant concentrations of Microcystin-LR. Ecotoxicol. Environ. Saf. 2020, 187, 109824. [Google Scholar] [CrossRef]
- Du, B.; Liu, G.; Ke, M.; Zhang, Z.; Zheng, M.; Lu, T.; Sun, L.; Qian, H. Proteomic analysis of the hepatotoxicity of Microcystis aeruginosa in adult zebrafish (Danio rerio) and its potential mechanisms. Environ. Pollut. 2019, 254, 113019. [Google Scholar] [CrossRef]
- He, J.; Li, G.; Chen, J.; Lin, J.; Zeng, C.; Chen, J.; Deng, J.; Xie, P. Prolonged exposure to low-dose microcystin induces nonalcoholic steatohepatitis in mice: A systems toxicology study. Arch. Toxicol. 2017, 91, 465–480. [Google Scholar] [CrossRef]
- Friedberg, E.C.; Walker, G.C.; Siede, W.; Wood, R.D.; Schultz, R.A.; Ellenberger, T. DNA Repair and Mutagenesis; ASM Press: Washington, DC, USA, 2006. [Google Scholar]
- Boiteux, S.; Jinks-Robertson, S. DNA repair mechanisms and the bypass of DNA damage in Saccharomyces cerevisiae. Genetics 2013, 193, 1025–1064. [Google Scholar] [CrossRef] [Green Version]
- Humpage, A.R.; Falconer, I.R. Microcystin-LR and liver tumor promotion: Effects on cytokinesis, ploidy, and apoptosis in cultured hepatocytes. Environ. Toxicol. 1999, 14, 61–75. [Google Scholar] [CrossRef]
- Dias, E.; Matos, P.; Pereira, P.; Batoréu, M.C.C.; Silva, M.J.; Jordan, P. Microcystin-LR activates the ERK1/2 kinases and stimulates the proliferation of the monkey kidney-derived cell line Vero-E6. Toxicol. In Vitro 2010, 24, 1689–1695. [Google Scholar] [CrossRef] [PubMed]
- Alverca, E.; Andrade, M.; Dias, E.; Sam Bento, F.; Batoréu, M.C.; Jordan, P.; Silva, M.J.; Pereira, P. Morphological and ultrastructural effects of microcystin-LR from Microcystis aeruginosa extract on a kidney cell line. Toxicon 2010, 54, 283–294. [Google Scholar] [CrossRef]
- Menezes, C.; Alverca, E.; Dias, E.; Sam-Bento, F.; Pereira, P. Involvement of endoplasmic reticulum and autophagy in microcystin-LR toxicity in Vero-E6 and HepG2 cell lines. Toxicol. In Vitro 2013, 27, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, E.; Andrade, M.; Alverca, E.; Pereira, P.; Batoréu, M.C.C.; Jordan, P.; Silva, M.J. Comparative study of the cytotoxic effect of microcistin-LR and purified extracts from Microcystis aeruginosa on a kidney cell line. Toxicon 2009, 53, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Chong, M.W.K.; Gu, K.D.; Lam, P.K.S.; Yang, M.; Fong, W.F. Study on the cytotoxicity of microcystin-LR on cultured cells. Chemosphere 2000, 41, 143–147. [Google Scholar] [CrossRef]
- Alhebshi, A.; Sideri, T.C.; Holland, S.L.; Avery, S.V. The essential iron-sulfur protein Rli1 is an important target accounting for inhibition of cell growth by reactive oxygen species. Mol. Biol. Cell. 2012, 23, 3582–3590. [Google Scholar] [CrossRef]
- Aitken, C.E.; Beznosková, P.; Vlčkova, V.; Chiu, W.-L.; Zhou, F.; Valášek, L.S.; Hinnebusch, A.G.; Lorsch, J.R. Eukaryotic translation initiation factor 3 plays distinct roles at the mRNA entry and exit channels of the ribosomal preinitiation complex. eLife 2016, 5, e20934. [Google Scholar] [CrossRef]
- Xie, Z.; Liu, S.; Zhang, Y.; Wang, Z. Roles of Rad23 protein in yeast nucleotide excision repair. Nucleic Acids Res. 2004, 32, 5981–5990. [Google Scholar] [CrossRef] [Green Version]
- Ross, S.J.; Findlay, V.J.; Malakasi, P.; Morgan, B.A. Thioredoxin peroxidase is required for the transcriptional response to oxidative stress in budding yeast. Mol. Biol. Cell. 2000, 11, 2631–2642. [Google Scholar] [CrossRef] [Green Version]
- Andreyev, A.Y.; Kushnareva, Y.E.; Starkov, A.A. Mitochondrial metabolism of reactive oxygen species. Biochemistry (Mosc.) 2005, 70, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Yadav, L.; Tamene, F.; Göös, H.; van Drogen, A.; Katainen, R.; Aebersold, R.; Gstaiger, M.; Varjosalo, M. Systematic analysis of human protein phosphatase interactions and dynamics. Cell Syst. 2017, 4, 430–444.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maclean, M.J.; Aamodt, R.; Harris, N.; Alseth, I.; Seeberg, E.; Bjørås, M.; Piper, P.W. Base excision repair activities required for yeast to attain a full chronological life span. Aging Cell 2003, 2, 93–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherman, F. Getting started with yeast. Methods Enzymol. 1991, 194, 3–21. [Google Scholar]
- Seidel, C. OD660 vs Number of Cells. Available online: http://www.pangloss.com/seidel/Protocols/ODvsCells.html (accessed on 30 June 2019).
- Wiśniewski, J.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Campos, A.; Danielsson, G.; Farinha, A.P.; Kuruvilla, J.; Warholm, P.; Cristobal, S. Shotgun proteomics to unravel marine mussel (Mytilus edulis) response to long-term exposure to low salinity and propranolol in a Baltic Sea microcosm. J. Proteom. 2016, 137, 97–106. [Google Scholar] [CrossRef]
- Wu, X.; Wang, Z. Relationships between yeast Rad27 and Apn1 in response to apurinic/apyrimidinic (AP) sites in DNA. Nucleic Acids Res. 1999, 27, 956–962. [Google Scholar] [CrossRef] [Green Version]
- Teste, M.; Duquenne, M.; François, J.M.; Parrou, J. Validation of reference genes for quantitative expression analysis by real-time RT-PCR in Saccharomyces cerevisiae. BMC Mol. Biol. 2009, 10, 99. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, F.H. Chapter 9-The real-time polymerase chain reaction (RT-PCR). In Calculations for Molecular Biology and Biotechnology, 2nd ed.; Frank, H., Stephenson, Eds.; Academic Press: Cambridge, MA, USA, 2010; pp. 211–311. ISBN 9780123756909. [Google Scholar]
- Multiple Experiment Viewer. Available online: https://www.mybiosoftware.com/mev-4-6-2-multiple-experiment-viewer.html (accessed on 31 January 2018).
- Pfaffl, M. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Cell Viability (%) |
|---|---|
| Control (cells grown in YPD) | 100.0 |
| With 50 mM H2O2 | 40.6 * |
| With 5% SDS | 10.8 * |
| With 0.2 nM MC-LR | 100.2 |
| With 0.4 nM MC-LR | 96.2 |
| With 1 nM MC-LR | 108.3 |
| With 10 nM MC-LR | 101.5 |
| With 100 nM MC-LR | 109.1 |
| With 1 µM MC-LR | 105.2 |
| MC-LR | ||||||
|---|---|---|---|---|---|---|
| Mechanisms Associated (a) | Proteins (b) | Control | 1 nM | 10 nM | 100 nM | 1 µM |
| Protein abundance increases in response to DNA replication stress | ENO1 | 0.0019 ± 0.00024 | 0.0054 ± 0.00078 * | 0.0038 ± 0.00233 | 0.0073 ± 0.00347 | 0.0031 ± 0.00005 * |
| PIL1 | 0.0 (c) | 0.0005 ± 0.00005 * | 0.0002 ± 0.00032 | 0.0006 ± 0.00033 | 0.0004 ± 0.0003 * | |
| TDH2 | 0.0009 ± 0.00103 | 0.0042 ± 0.00098 * | 0.0017 ± 0.00291 | 0.0045 ± 0.00053 * | 0.0036 ± 0.00038 * | |
| ARC1 | 0.0016 ± 0.00040 | 0.0014 ± 0.00013 | 0.0018 ± 0.00033 | 0.0011 ± 0.00032 | 0.0005 ± 0.00050 * | |
| TRR1 ≈ TRR2 | 0.0005 ± 0.00081 | 0.0031 ± 0.00122 * | 0.0012 ± 0.00202 | 0.0027 ± 0.00075 * | 0.0040 ± 0.00041 * | |
| Nuclear Excision Repair Factor 2 (NEF2) | RAD23 | 0.0007 ± 0.00019 | 0.0006 ± 0.00029 | 0.0006 ± 0.00019 | 0.0012 ± 0.00010 * | 0.0013 ± 0.00016 * |
| Gene upregulated in cancer | NOP2 | 0.0025 ± 0.00041 | 0.0010 ± 0.00023 * | 0.0011 ± 0.00050 * | 0.0012 ± 0.00044 * | 0.0011 ± 0.00014 * |
| Responds to oxidative stress | MGE1 | 0.0012 ± 0.00034 | 0.0 * | 0.0 * | 0.0007 ± 0.00006 | 0.0009 ± 0.00009 |
| Involved in actin cytoskeleton maintenance | TCP1 | 0.0 | 0.0003 ± 0.00029 * | 0.0005 ± 0.00014 | 0.0002 ± 0.00016 | 0.0 |
| Several ribosomal proteins | RPL36B | 0.0034 ± 0.0011 | 0.0053 ± 0.00137 | 0.0109 ± 0.00463 | 0.00474 ± 0.00146 | 0.0073 ± 0.00186 * |
| RPL32 | 0.0116 ± 0.00202 | 0.0065 ± 0.00136 * | 0.0080 ± 0.00228 | 0.0077 ± 0.00093 | 0.0082 ± 0.00090 | |
| RPL24A | 0.0022 ± 0.00063 | 0.0009 ± 0.00081 | 0.0 * | 0.0009 ± 0.00077 | 0.0005 ± 0.00086 | |
| RPS6B | 0.0099 ± 0.00124 | 0.0057 ± 0.00101 * | 0.0099 ± 0.00190 | 0.0059 ± 0.00086 * | 0.0069 ± 0.00189 | |
| RPS20 | 0.0156 ± 0.00120 | 0.0100 ± 0.00488 | 0.0153 ± 0.00272 | 0.0080 ± 0.00186 * | 0.0121 ± 0.00077 * | |
| RPS2 | 0.0011 ± 0.00031 | 0.0039 ± 0.00106 * | 0.0021 ± 0.00079 | 0.0041 ± 0.00095 * | 0.0029 ± 0.00068 * | |
| RPS23A | 0.0 | 0.0020 ± 0.00116 | 0.0010 ± 0.00095 | 0.0025 ± 0.00064 * | 0.0018 ± 0.00071 * | |
| Activity in nuclear ribosomal-subunit export impaired by mild oxidative stress | RLI1 | 0.0005 ± 0.00013 | 0.0 * | 0.0004 ± 0.00013 | 0.0 * | 0.0001 ± 0.00018 |
| Involved in ER to Golgi transport | SEC21 | 0.0 | 0.0001 ± 0.00020 | 0.0001 ± 0.00011 | 0.0003 ± 0.00003 * | 0.0 |
| SAR1 | 0.0 | 0.0009 ± 0.00008 * | 0.0 | 0.00053 ± 0.00046 | 0.0003 ± 0.00057 | |
| Decreases upon DNA replication stress | PNC1 | 0.0 | 0.0008 ± 0.00007 * | 0.0003 ± 0.00050 | 0.0012 ± 0.00035 * | 0.0009 ± 0.00010 * |
| VPH1 | 0.0 | 0.0002 ± 0.00002 * | 0.0001 ± 0.00013 | 0.0004 ± 0.00020 | 0.0001 ± 0.00016 | |
| Specific translational activator for the mitochondrial COX1 mRNA | MAM33 | 0.0016 ± 0.00019 | 0.0011 ± 0.00035 | 0.0012 ± 0.00020 | 0.0004 ± 0.00033 * | 0.0013 ± 0.00050 |
| Enzyme of the ‘de novo’ purine nucleotide biosynthetic pathway | ADE5,7 | 0.0 | 0.0001 ± 0.00014 | 0.0003 ± 0.00010 * | 0.0001 ± 0.00011 | 0.0 |
| Polypeptide release factor (eRF1) in translation termination | SUP45 | 0.0 | 0.0005 ± 0.00018 * | 0.0001 ± 0.00025 | 0.0007 ± 0.00006 * | 0.0002 ± 0.00030 |
| Interaction with Top1p and nucleolar localization are negatively regulated by polyphosphorylation | NSR1 | 0.0078 ± 0.00074 | 0.0065 ± 0.00097 | 0.0091 ± 0.00155 | 0.0061 ± 0.00025 * | 0.0061 ± 0.00052 * |
| Required for the assembly of actin and tubulins in vivo | CCT4 | 0.0001 ± 0.00024 | 0.0012 ± 0.00018 * | 0.0002 ± 0.00041 | 0.0010 ± 0.00060 | 0.0006 ± 0.00025 |
| Heat Shock Protein | HSP10 | 0.0072 ± 0.00106 | 0.0028 ± 0.00091 * | 0.0059 ± 0.00213 | 0.0030 ± 0.00026 * | 0.0044 ± 0.00085 * |
| Involved in glycolysis and gluconeogenesis | GPM1 | 0.0105 ± 0.00076 | 0.0081 ± 0.00131 | 0.0078 ± 0.00037 * | 0.0079 ± 0.00037 * | 0.0091 ± 0.00103 |
| Involved in TCA cycle | FUM1 | 0.0 | 0.0001 ± 0.00022 | 0.0001 ± 0.00022 | 0.0003 ± 0.00003 * | 0.0006 ± 0.00031 |
| Catalyzes the oxidation of isocitrate to alpha-ketoglutarate | IDP1 | 0.0 | 0.0010 ± 0.00014 * | 0.0 | 0.0010 ± 0.00028 * | 0.0006 ± 0.00051 |
| Gene | Primer Sequence | Molecular Function | Primers Designed in |
|---|---|---|---|
| Apn1 | F: 5’-TGG GTT TCT CCG CAG TAT-3’ | AP endonuclease/3′-diesterases, enzymes whose function is to excise mutagenic and cytotoxic AP or 3′ phosphate/phosphoglycolate groups from DNA [36] | This study |
| R: 5’-GCC TAT CCC TAA TTG CTC AC-3’ | |||
| Apn2 | F: 5’-TGC TAA TGG GCG ACG TAA AT-3’ | This study | |
| R: 5’-GGC GTG TCC GGA TTG ATA ATA-3’ | |||
| Rad27 | F: 5’-CCG CAG CAA GTG AAG ATA TG-3’ | 5′→3′ exonuclease and a flap endonuclease [41] | This study |
| R: 5’-CCA ACA CCT CTG ATG CTT TC-3’ | |||
| Ntg1 | F: 5’-CAT TCC TGT AAC GGT TGC CT-3’ | DNA glycosylases/AP lyases. These excise primarily oxidized pyrimidines [36] | [17] |
| R: 5’-TTG TGT GGA ACC CAA CTG AA-3’ | |||
| Ntg2 | F: 5’-AAC ACT GCA AAA AGG TTG GG-3’ | [17] | |
| R: 5’-GAC CAA ATC CAA CCA AAA CG-3’ |
| Gene | Annealing Temperature (°C) | Efficiency (%) | Slope | Y Intercept | R2 | |
|---|---|---|---|---|---|---|
| BER genes | Apn1 | 52 | 98 | −3.361 | 24.083 | 0.997 |
| Apn2 | 52 | 100 | −3.314 | 26.196 | 0.992 | |
| Rad27 | 52 | 96 | −3.416 | 24.524 | 0.998 | |
| Ntg1 | 48 | 88 | −3.649 | 25.863 | 0.993 | |
| Ntg2 | 48 | 94 | −3.484 | 21.605 | 0.993 | |
| Reference gene | Alg9 | 58 | 97 | −3.398 | 23.906 | 0.989 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valério, E.; Barreiros, S.; Rodrigues, S.; Turkina, M.V.; Vasconcelos, V.M.; Campos, A. New Insights in Saccharomyces cerevisiae Response to the Cyanotoxin Microcystin-LR, Revealed by Proteomics and Gene Expression. Toxins 2020, 12, 667. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12100667
Valério E, Barreiros S, Rodrigues S, Turkina MV, Vasconcelos VM, Campos A. New Insights in Saccharomyces cerevisiae Response to the Cyanotoxin Microcystin-LR, Revealed by Proteomics and Gene Expression. Toxins. 2020; 12(10):667. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12100667
Chicago/Turabian StyleValério, Elisabete, Sara Barreiros, Sara Rodrigues, Maria V. Turkina, Vitor M. Vasconcelos, and Alexandre Campos. 2020. "New Insights in Saccharomyces cerevisiae Response to the Cyanotoxin Microcystin-LR, Revealed by Proteomics and Gene Expression" Toxins 12, no. 10: 667. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12100667