Degradation and Detoxification of Aflatoxin B1 by Tea-Derived Aspergillus niger RAF106

Abstract
:1. Introduction
2. Results
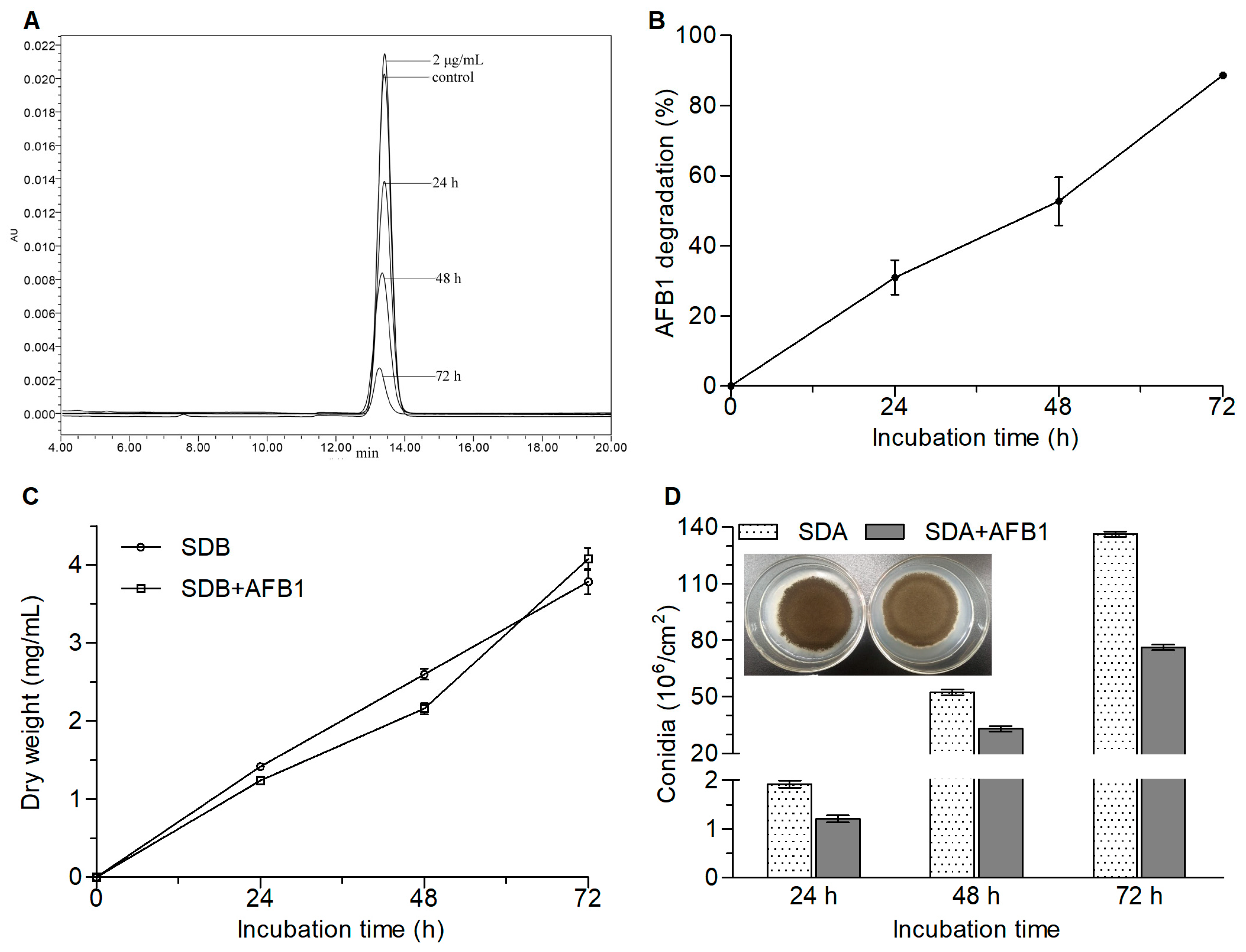
2.1. AFB1 Degradation by A. niger RAF106 and the Effect of AFB1 on Fungal Development
2.2. Effects of the Fermentation Conditions on AFB1 Degradation
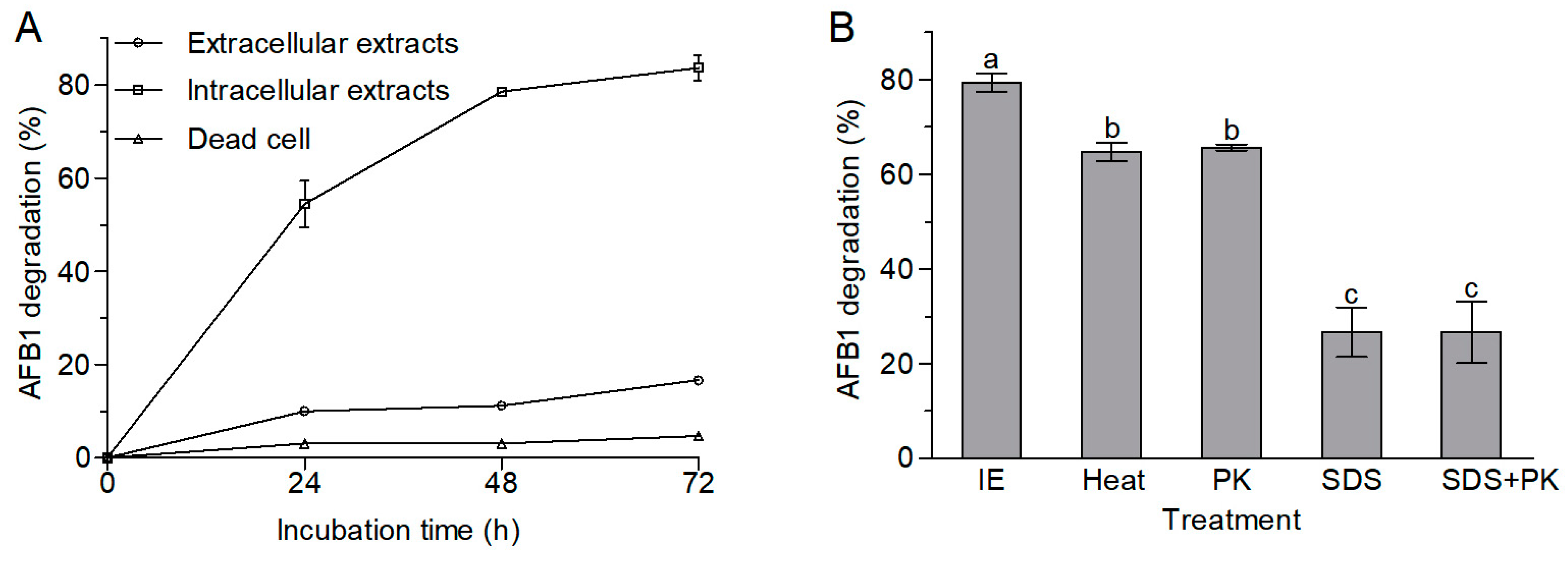
2.3. AFB1 Degradation by Extracellular and Intracellular Extracts and Dead Cells of A. niger RAF106
2.4. The Ames Test for Mutagenicity
2.5. Analysis of the AFB1 Degradation Products
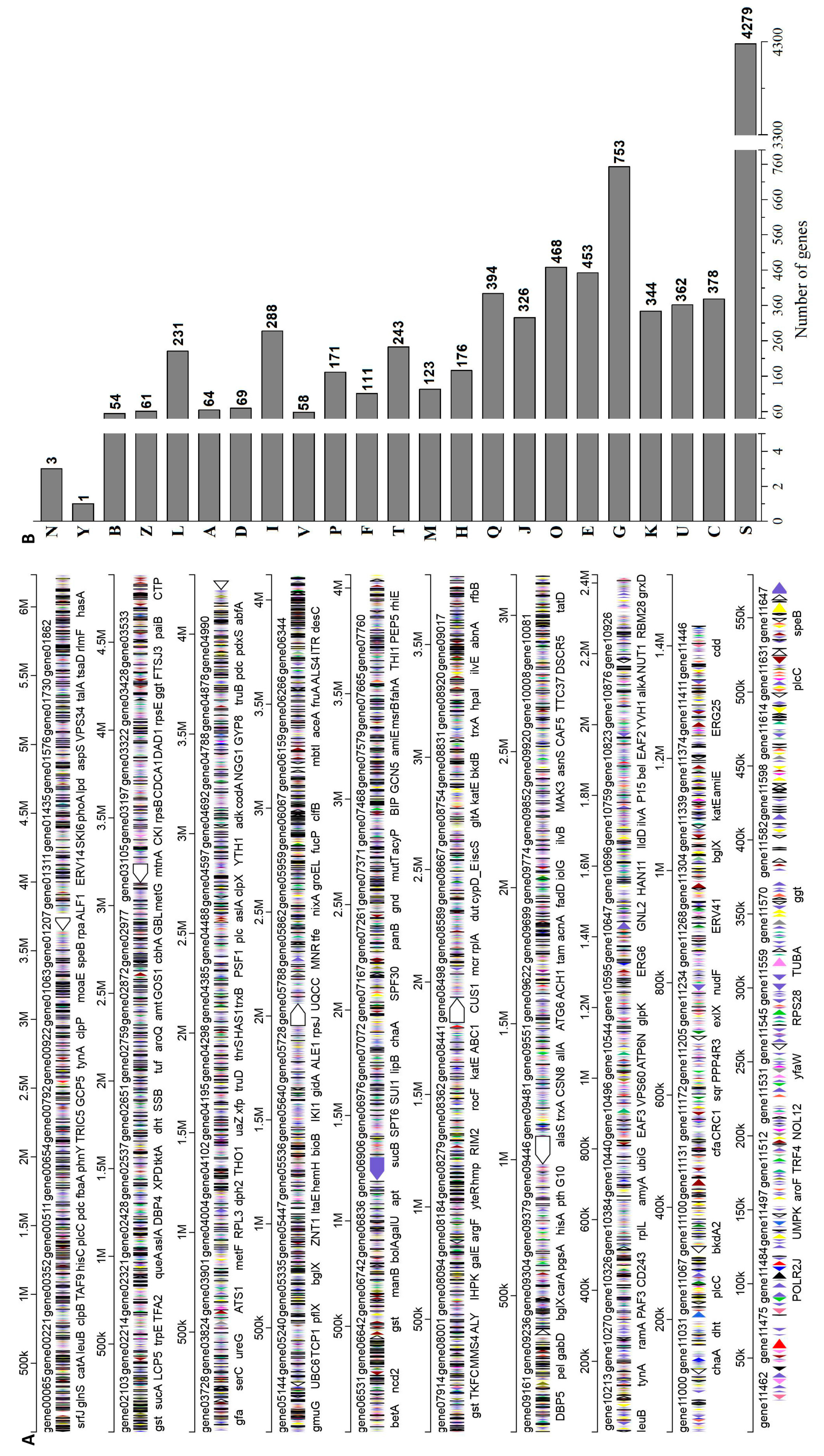
2.6. Genomic Characteristics of A. niger RAF106
3. Discussion
4. Conclusions
5. Material and Methods
5.1. Fungal Strains and Culture Conditions
5.2. AFB1 Degradation by A. niger RAF106 in Liquid Culture
5.3. Quantification of AFB1 Using HPLC
5.4. Changes in Fungal Growth and Aerial Conidiation Caused by AFB1 in A. niger RAF106
5.5. Effects of Nutrients, pH, and Temperature on AFB1 Degradation and Fungal Growth
5.6. AFB1 Degradation by Extracellular Extracts, Intracellular Extracts, and Dead Cells
5.7. Effects of Proteinase K, SDS, and Heat Treatment on the Degradation of AFB1
5.8. Analysis of the AFB1 Metabolites Using HPLC-Q-TOF-MS
5.9. Ames Mutagenicity Assay
5.10. Genome Sequencing and Analysis
5.11. Data Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Lee, H.J.; Ryu, D. Worldwide occurrence of mycotoxins in cereals and cereal-derived food products: Public health perspectives of their co-occurrence. J. Agric. Food Chem. 2017, 65, 7034–7051. [Google Scholar] [CrossRef] [PubMed]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food Sci. 2019, 1549–7852. [Google Scholar] [CrossRef] [PubMed]
- Nugraha, A.; Khotimah, K.; Rietjens, I.M.C.M. Risk assessment of aflatoxin B1 exposure from maize and peanut consumption in Indonesia using the margin of exposure and liver cancer risk estimation approaches. Food Chem. Toxicol. 2018, 113, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Cui, A.; Hua, H.; Shao, T.; Song, P.; Kong, Q.; Luo, T.; Jiang, Y. Aflatoxin B1 induces Src phosphorylation and stimulates lung cancer cell migration. Tumour Biol. 2015, 36, 6507–6513. [Google Scholar] [CrossRef]
- Lahoum, A.; Verheecke-Vawssen, C.; Bouras, N.; Sabaou, N.; Mathieu, F. Taxonomy of mycelial actinobacteria isolated from Saharan soils and their effiency to reduce aflatoxin B1 content in a solid-based medium. Ann. Microbiol. 2017, 67, 231–237. [Google Scholar] [CrossRef]
- Lee, L.S.; Dunn, J.J.; Delucca, A.J.; Ciegler, A. Role of lactone ring of aflatoxin B1 in toxicity and mutagenicity. Experientia 1981, 37, 16–17. [Google Scholar] [CrossRef]
- Wogan, G.; Edwards, G.; Newberne, P. Structure-activity relationships in toxicity and carcinogenicity of aflatoxins and analogs. Cancer Res. 1971, 31, 1936–1942. [Google Scholar]
- Cullen, J.M.; Ruebner, B.H.; Hsieh, L.S.; Hyde, D.M.; Hsieh, D.P. Carcinogenicity of dietary aflatoxin M1 in male fischer rats compared to aflatoxin B1. Cancer Res. 1987, 47, 1913–1917. [Google Scholar]
- Luongo, D.; Russo, R.; Balestrieri, A.; Marzocco, S.; Bergamo, P.; Severino, L. In vitro study of AFB1 and AFM1 effects on human lymphoblastoid Jurkat T-cell model. J. Immunotoxicol. 2013, 11, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Pitt, J.I.; Miller, J.D. A concise history of mycotoxin research. J. Agric. Chem. 2017, 65, 7021–7033. [Google Scholar] [CrossRef]
- Wu, F.; Mitchell, N.J. How climate change and regulations can affect the economics of mycotoxins. World Mycotoxin J. 2016, 9, 653–663. [Google Scholar] [CrossRef]
- Williams, J.H.; Phillips, T.D.; Jolly, P.E.; Stiles, J.K.; Jolly, C.M.; Aggarwal, D. Human aflatoxicosis in developing countries: A review of toxicology, exposure, potential health consequences, and interventions. Am. J. Clin. Nutr. 2004, 80, 1106–1122. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.R.; Yang, S.M.; Cho, S.M.; Kim, M.; Hong, S.Y.; Chung, S.H. Aflatoxin B1 detoxification by Aspergillus oryzae from Meju, a traditional Korean fermented soybean starter. J. Microbiol. Biotechnol. 2017, 27, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Mishra, H.N.; Das, C. A review on biological control and metabolism of aflatoxin. Front. Pharmacol. 2003, 43, 245–264. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Jezkova, A.; Yuan, Z.; Pavlikova, L.; Dohnal, V.; Kuca, K. Biological degradation of aflatoxins. Drug Metab. Rev. 2009, 41, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Kong, Q.; Chi, C.; Shan, S.; Guan, B. Biotransformation of aflatoxin B-1 and aflatoxin G(1) in peanut meal by anaerobic solid fermentation of Streptococcus thermophilus and Lactobacillus delbrueckii subsp. bulgaricus. Int. J. Food Microbiol. 2015, 211, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Samuel, M.S.; Sivaramakrishna, A.; Mehta, A. Degradation and detoxification of aflatoxin B1 by Pseudomonas putida. Int. Biodeter. Biodegrad. 2014, 86, 202–209. [Google Scholar] [CrossRef]
- Eshelli, M.; Harvey, L.; Edrada-Ebel, R.; McNeil, B. Metabolomics of the bio-degradation process of aflatoxin B1 by actinomycetes at an initial pH of 6.0. Toxins 2015, 7, 439–456. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Ogata, M.; Hirai, H.; Kawagishi, H. Detoxification of aflatoxin B1 by manganese peroxidase from the white-rot fungus Phanerochaete sordida YK-624. FEMS Microbiol. Lett. 2011, 314, 164–169. [Google Scholar] [CrossRef]
- Ji, C.; Fan, Y.; Zhao, L. Review on biological degradation of mycotoxins. Anim. Nutr. 2016, 2, 127–133. [Google Scholar] [CrossRef]
- Shu, X.; Wang, Y.; Zhou, Q.; Li, M.; Hu, H.; Ma, Y.; Chen, X.; Ni, J.; Zhao, W.; Huang, S.; et al. Biological degradation of Aflatoxin B-1 by cell-free extracts of Bacillus velezensis DY3108 with broad pH stability and excellent thermostability. Toxins 2018, 10, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adebo, O.A.; Njobeh, P.B.; Sidu, S.; Tlou, M.G.; Mavumengwana, V. Aflatoxin B1 degradation by liquid cultures and lysates of three bacterial strains. Int. J. Food Microbiol. 2016, 233, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Cairns, T.C.; Nai, C.; Meyer, V. How a fungus shapes biotechnology: 100 years of Aspergillus niger research. Fungal Biol. Biotechnol. 2018, 5, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.S.; Jun, S.C.; Han, K.H.; Hong, S.B.; Yu, J.H. Diversity, application, and synthetic biology of industrially important Aspergillus fungi. Adv. Appl. Microbiol. 2017, 100, 161–202. [Google Scholar]
- Ma, Y.; Ling, T.; Su, X.; Jiang, B.; Nian, B.; Chen, L.; Liu, M.; Zhang, Z.; Wang, D.; Mu, Y.; et al. Integrated proteomics and metabolomics analysis of tea leaves fermented by Aspergillus niger, Aspergillus tamarii and Aspergillus fumigatus. Food Chem. 2020, 334, 127560. [Google Scholar] [CrossRef]
- Zhang, W.; Xue, B.; Li, M.; Mu, Y.; Shan, A. Screening a strain of Aspergillus niger and optimization of fermentation conditions for degradation of Aflatoxin B1. Toxins 2014, 6, 3157–3172. [Google Scholar] [CrossRef]
- Sun, X.; Sun, C.; Zhang, X.; Zhang, H.; Tang, L. Aflatoxin B1 decontamination by UV-mutated live and immobilized Aspergillus niger. Food Control 2015, 61, 235–242. [Google Scholar] [CrossRef]
- Mann, R.; Rehm, H.J. Degradation products from aflatoxin B1 by Corynebacterium rubrum, Aspergillus niger, Trichoderma viride and Mucor ambiguus. Appl. Microbiol. Biotechnol. 1976, 2, 297–306. [Google Scholar]
- Andersen, M.R.; Salazar, M.P.; Schaap, P.J.; Van de Vondervoort, P.J.I.; Culley, D.; Thykaer, J.; Frisvad, J.C.; Nielsen, K.F.; Albang, R.; Albermann, K.; et al. Comparative genomics of citric-acid-producing Aspergillus niger ATCC 1015 versus enzyme-producing CBS 513.88. Genome Res. 2011, 21, 885–897. [Google Scholar] [CrossRef] [Green Version]
- Frisvad, J.C.; Møller, L.L.H.; Larsen, T.O.; Kumar, R.; Arnau, J. Safety of the fungal workhorses of industrial biotechnology: Update on the mycotoxin and secondary metabolite potential of Aspergillus niger, Aspergillus oryzae, and Trichoderma reesei. Appl. Microbiol. Biotechnol. 2018, 102, 9481–9515. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Du, M.; Liu, T.; Fang, Q.; Liao, Z.; Zhong, Q.; Chen, J.; Meng, X.; Zhou, S.; Wang, J. Changes in the biotransformation of green tea catechins induced by different carbon and nitrogen sources in Aspergillus niger RAF106. Front. Microbiol. 2019, 10, 2521. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.W.; Pryor, B.M. Degradation of aflatoxin B1 from naturally contaminated maize using the edible fungus Pleurotus ostreatus. AMB Express 2017, 7, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mwakinyali, S.E.; Ming, Z.; Xie, H.; Zhang, Q.; Li, P. Investigation and characterization of Myroides odoratimimus Strain 3J2MO aflatoxin B1 degradation. J. Agric. Food Chem. 2019, 67, 4595–4602. [Google Scholar] [CrossRef] [PubMed]
- Verheecke, C.; Liboz, T.; Mathieu, F. Microbial degradation of aflatoxin B1: Current status and future advances. Int. J. Food Microbiol. 2016, 237, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brzonkalik, K.; Herrling, T.; Syldatk, C.; Neumann, A. The influence of different nitrogen and carbon sources on mycotoxin production in Alternaria alternate. Int. J. Food Microbiol. 2011, 147, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Farzaneh, M.; Shi, Z.; Ghassempour, A.; Sedaghat, N.; Ahmadzadeh, M.; Mirabolfathy, M.; Javan-Nikkhah, M. Aflatoxin B1 degradation by Bacillus subtilis UTBSP1 isolated from pistachio nuts of Iran. Food Control 2012, 23, 100–106. [Google Scholar] [CrossRef]
- Müller, R.; Antranikian, G.; Maloney, S.; Sharp, R. Thermophilic degradation of environmental pollutants. In: Biotechnology of Extremophiles. Adv. Biochem. Eng. Biotechnol. 2007, 61, 155–169. [Google Scholar]
- Guan, S.; Cheng, J.; Ting, Z.; Junxia, L.; Qiugang, M.; Tiangui, N. Aflatoxin B1 degradation by Stenotrophomonas Maltophilia and other microbes selected using coumarin medium. Int. J. Mol. Sci. 2008, 9, 1489–1503. [Google Scholar] [CrossRef] [Green Version]
- Alborch, L.; Bragulat, M.R.; Abarca, M.L.; Cabañes, F.J. Effect of water activity, temperature and incubation time on growth and ochratoxin a production by Aspergillus niger and Aspergillus carbonarius on maize kernels. Int. J. Food Microbiol. 2011, 147, 53–57. [Google Scholar] [CrossRef]
- Héctor, P.; Hiromi, T.M.; Minoru, H.J.; De, M.H.C. Growth of Aspergillus ochraceus, A. carbonarius and A. niger on culture media at different water activities and temperatures. Braz. J. Microbiol. 2005, 36, 24–28. [Google Scholar]
- Colin, V.; Baigorí, M.; Pera, L. Tailoring fungal morphology of Aspergillus niger MYA 135 by altering the hyphal morphology and the conidia adhesion capacity: Biotechnological applications. AMB Express 2013, 3, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, C.H.; Lung, M.Y. Effect of pH on the production and molecular weight distribution of exopolysaccharide by Antrodia camphorate in batch cultures. Process Biochem. 2004, 39, 931–937. [Google Scholar] [CrossRef]
- Kong, Q.; Zhai, C.; Guan, B.; Li, C.; Shan, S.; Yu, J. Mathematic modeling for optimum conditions on aflatoxin B1 degradation by the aerobic bacterium Rhodococcus erythropolis. Toxins 2012, 4, 1181–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.H.; Guan, S.; Gao, X.; Ma, Q.G.; Lei, Y.P.; Bai, X.M.; Ji, C. Preparation, purification and characteristics of an aflatoxin degradation enzyme from Myxococcus fulvus ANSM068. J. Appl. Microbiol. 2010, 110, 147–155. [Google Scholar] [CrossRef]
- Hathout, A.S.; Aly, S.E. Biological detoxification of mycotoxins: A review. Ann. Microbiol. 2014, 64, 905–919. [Google Scholar] [CrossRef]
- Li, J.; Huang, J.; Jin, Y.; Wu, C.; Shen, D.; Zhang, S.; Zhou, R. Aflatoxin B1 degradation by salt tolerant Tetragenococcus halophilus CGMCC 3792. Food Chem. Toxicol. 2018, 121, 430–436. [Google Scholar] [CrossRef]
- Li, J.; Huang, J.; Jin, Y.; Wu, C.; Shen, D.; Zhang, S.; Zhou, R. Mechanism and kinetics of degrading aflatoxin B1 by salt tolerant Candida versatilis CGMCC 3790. J. Hazard. Mater. 2018, 359, 382–387. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.; Yan, H.; Yin, C.; Liu, Y.; Xu, Q.; Liu, X.; Zhang, Z. Effective biodegradation of aflatoxin B1 using the Bacillus licheniformis (BL010) strain. Toxins 2018, 10, 497. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wu, J.; Liu, Z.; Shi, Y.; Liu, J.; Xu, X.; Hao, S.; Mu, P.; Deng, F.; Deng, Y. Aflatoxin B1 degradation and detoxification by Escherichia coli CG1061 isolated from chicken. Front. Pharmacol. 2019, 9. [Google Scholar] [CrossRef]
- Rao, K.R.; Vipin, A.V.; Hariprasad, P.; Appaiah, K.A.A.; Venkateswaran, G. Biological detoxification of aflatoxin B1 by Bacillus licheniformis CFR1. Food Control 2016, 71, 234–241. [Google Scholar]
- Alberts, J.F.; Engelbrecht, Y.; Steyn, P.S.; Holzapfel, W.H.; Van Zyl, W.H. Biological degradation of aflatoxin B1 by Rhodococcus erythropolis cultures. Int. J. Food Microbiol. 2006, 109, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Pel, H.J.; De Winde, J.H.; Archer, D.B.; Dyer, P.S.; Hofmann, G.; Schaap, P.J.; Turner, G.; De Vries, R.P.; Albang, R.; Albermann, K.; et al. Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nat. Biotechnol. 2007, 25, 221–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyake, Y.; Ito, C.; Itoigawa, M.; Osawa, T. Isolation of the antioxidant pyranonigrin-A from rice mold starters used in the manufacturing process of fermented foods. J. Agric. Chem. Soc. Jpn. 2007, 71, 2515–2521. [Google Scholar] [CrossRef] [PubMed]
- Inglis, D.O.; Binkley, J.; Skrzypek, M.S.; Arnaud, M.B.; Cerqueira, G.C.; Shah, P.; Wymore, F.; Wortman, J.R.; Sherlock, G. Comprehensive annotation of secondary metabolite biosynthetic genes and gene clusters of Aspergillus nidulans, A. fumigatus, A. niger and A. oryzae. BMC Microbiol. 2013, 13, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurome, T.; Inami, K.; Inoue, T.; Ikai, K.; Takesako, K.; Kato, I.; Shiba, T. Total synthesis of an antifungal cyclic depsipeptide aureobasidin A. Tetrahedron 1996, 52, 4327–4346. [Google Scholar] [CrossRef]
- Choi, J.S.; Lee, H.J.; Park, K.Y.; Ha, J.O.; Kang, S.S. In vitro antimutagenic effects of anthraquinone aglycones and naphthopyrone glycosides from Cassia tora. Planta Med. 1997, 63, 11–14. [Google Scholar] [CrossRef]
- Walton, J.D. HC-toxin. Phytochemistry 2006, 67, 1406. [Google Scholar] [CrossRef] [Green Version]
- Godio, R.P.; Fouces, R.; Gudiña, E.J.; Martín, J.F. Agrobacterium tumefaciens-mediated transformation of the antitumor clavaric acid-producing basidiomycete Hypholoma sublateritium. Curr. Genet. 2004, 46, 287–294. [Google Scholar] [CrossRef]
- Yin, C.; Wang, B.; He, P.; Lin, Y.; Pan, L. Genomic analysis of the aconidial and high-performance protein producer, industrially relevant Aspergillus niger SH2 strain. Gene 2014, 541, 107–114. [Google Scholar] [CrossRef]
- Gong, W.; Cheng, Z.; Zhang, H.; Liu, L.; Gao, P.; Wang, L. Draft genome sequence of Aspergillus niger strain An76. Genome Announc. 2016, 4, e01700-15. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Shin, H.; Li, J.; Du, G.; Liu, L.; Chen, J. Comparative genomics and transcriptome analysis of Aspergillus niger and metabolic engineering for citrate production. Sci. Rep. 2017, 7, 41040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malekpour, A.; Bayati, S. Stimultaneous determination of aflatoxins in pistachio using ultrasonically stabilized chloroform/water emulsion and HPLC. Food Anal. Methods 2015, 9, 805–811. [Google Scholar] [CrossRef]
- Ames, B.N.; Durston, W.E.; Yamasaki, E.; Lee, F.D. Carcinogens are mutagens: A simple test system combining liver homogenates for activation and bacteria for detection. Proc. Natl. Acad. Sci. USA 1973, 70, 2281–2285. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster Type | MIBiG ID * | Similarity | Cluster Type | MIBiG ID | Similarity |
|---|---|---|---|---|---|
| Scaffold 1 | Scaffold 2 | ||||
| Aflatrem | BGC0000629 | 11% | Aflavarin | BGC0001304 | 40% |
| Notoamide | BGC0000818 | 11% | Nidulanin A | BGC0001699 | 75% |
| Pyripyropene A | BGC0001068 | 25% | Fusarielin H | BGC0001600 | 37% |
| Scaffold 3 | Pyranonigrin E | BGC0001124 | 100% | ||
| Fumonisin | BGC0000063 | 11% | |||
| Scaffold 5 | Scaffold 6 | ||||
| Azaphilone | BGC0000027 | 16% | HC-toxin | BGC0000370 | 100% |
| Pyranonigrin E | BGC0001124 | 100% | Clavaric acid | BGC0001248 | 100% |
| TAN-1612 | BGC0000156 | 100% | Naphthopyrone | BGC0000107 | 100% |
| Wortmanamide A | BGC0001466 | 100% | Aflatrem | BGC0000629 | 17% |
| Communesin | BGC0001205 | 12% | |||
| Shearinine D | BGC0001776 | 13% | Scaffold 9 | ||
| Scaffold 7 | Aureobasidin A1 | BGC0000307 | 100% | ||
| Citrinin | BGC0001338 | 18% | HC-toxin | BGC0000370 | 100% |
| Aflatrem | BGC0000629 | 25% | Duclauxin | BGC0001578 | 42% |
| Neurosporin A | BGC0001697 | 40% | |||
| Gene ID | Annotation | Similarity |
|---|---|---|
| Scaffold 1 | ||
| gene00348 | Secondary metabolism regulator laeA | 83.1% |
| gene00577 | 40S ribosomal protein S27a | 100% |
| gene00593 | Aspartic protease Pep2 | 87.7% |
| gene00993 | Pyrophosphate phospho-hydrolase | 93.1% |
| gene01168 | Cell division control protein 42 | 80.1% |
| gene01307 | Phosphatidate cytidylyltransferase | 88.7% |
| gene01444 | Transcriptional regulatory protein RPD3 | 87.8% |
| gene01632 | Chitin synthase C | 80.9% |
| gene01911 | Fatty acid desaturase | 92.8% |
| Scaffold 2 | ||
| gene02216 | 1,3-beta-glucan synthase component FKS1 | 89% |
| gene02229 | Translation initiation factor eif-2b epsilon subunit | 83.4% |
| gene02256 | Mitogen-activated protein kinase | 93.4% |
| gene02307 | GTP-binding nuclear protein GSP1/Ran | 86% |
| gene02534 | Histone H3 | 97.7% |
| gene02609 | Guanine nucleotide-binding protein subunit α | 94.1% |
| gene02632 | Mitogen-activated protein kinase hog1 | 95.1% |
| gene02634 | G protein complex alpha subunit GpaB | 91.6% |
| gene02843 | β-tubulin | 97% |
| gene03109 | Glucosamine-fructose-6-phosphate aminotransferase | 96.4% |
| gene03500 | Guanine nucleotide-binding protein subunit β | 85.2% |
| Scaffold 3 | ||
| gene03862 | Bifunctional tryptophan synthase TRPB | 85.2% |
| gene03939 | Secretory pathway GDP dissociation inhibitor | 84.6% |
| gene03951 | GlcNAc-1-P transferase | 81.3% |
| gene03962 | Ribosomal RNA assembly protein mis3 | 88.3% |
| gene04302 | Mitogen-activated protein kinase Fus3 | 85.5% |
| gene04382 | Thioredoxin reductase Trr1/Trr2 | 85.3% |
| gene04618 | α-Tubulin | 92.3% |
| gene04834 | Ubiquitin | 95.9% |
| Scaffold 4 | ||
| gene05492 | Tubulin α-1 chain | 90.8% |
| gene05524 | Ferrochelatase | 87.6% |
| gene05596 | Citrate synthase Cit1 | 88.4% |
| gene05603 | Malate synthase | 81.8% |
| gene05712 | Imidazoleglycerol-phosphate dehydratase | 86.9% |
| gene05752 | Mitochondrial import receptor subunit tom40 | 86.5% |
| gene05805 | Ornithine-N5-oxygenase | 81.4% |
| gene05876 | Peptidyl-prolyl cis-trans isomerase ssp-1 | 81.6% |
| Scaffold 5 | ||
| gene07009 | Rho-related protein rac1 | 85.1% |
| gene07037 | Phosphoadenosine phosphosulfate reductase | 85.1% |
| gene07271 | U1 snRNP splicing complex subunit | 94.6% |
| gene07607 | Cell division control protein 2 | 90% |
| Scaffold 6 | ||
| gene08487 | Secretion related GTPase | 82.6% |
| gene08506 | GTP-binding protein | 99% |
| gene08568 | S-adenosylmethionine decarboxylase proenzyme | 83.4% |
| gene08576 | Glutamyl-tRNA synthetase | 83.4% |
| gene08706 | Golgi transport protein Sly1 | 84.4% |
| Scaffold 7 | ||
| gene09163 | 1,3-β-glucanosyltransferase Gel1 | 85.4% |
| gene09291 | Homoserine O-acetyltransferase | 87.2% |
| gene09449 | GTP-binding protein rhb1 | 95.2% |
| gene09466 | Acetyl-CoA acetyltransferase | 92.2% |
| gene09749 | Eukaryotic translation initiation factor 3 subunit | 80.6% |
| gene09874 | Aureobasidin resistance protein Aur1 | 92% |
| gene09879 | Sec14 cytosolic factor | 84.8% |
| gene09963 | DNA primase subunit Pri1 | 80.4% |
| Scaffold 8 | ||
| Gene10341 | Orotate phosphoribosyltransferase | 80.9% |
| Gene10380 | T-complex protein 1 | 82.8% |
| Gene10446 | Polyubiquitin | 98.7% |
| Gene10497 | Ribose-phosphate pyrophosphokinase | 86.1% |
| Gene10549 | Saccharopine dehydrogenase | 89.5% |
| Gene10566 | Mannose-1-phosphate guanyltransferase | 93.7% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, Q.; Du, M.; Chen, J.; Liu, T.; Zheng, Y.; Liao, Z.; Zhong, Q.; Wang, L.; Fang, X.; Wang, J. Degradation and Detoxification of Aflatoxin B1 by Tea-Derived Aspergillus niger RAF106. Toxins 2020, 12, 777. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12120777
Fang Q, Du M, Chen J, Liu T, Zheng Y, Liao Z, Zhong Q, Wang L, Fang X, Wang J. Degradation and Detoxification of Aflatoxin B1 by Tea-Derived Aspergillus niger RAF106. Toxins. 2020; 12(12):777. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12120777
Chicago/Turabian StyleFang, Qian’an, Minru Du, Jianwen Chen, Tong Liu, Yong Zheng, Zhenlin Liao, Qingping Zhong, Li Wang, Xiang Fang, and Jie Wang. 2020. "Degradation and Detoxification of Aflatoxin B1 by Tea-Derived Aspergillus niger RAF106" Toxins 12, no. 12: 777. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12120777