New Insectotoxin from Tibellus Oblongus Spider Venom Presents Novel Adaptation of ICK Fold

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
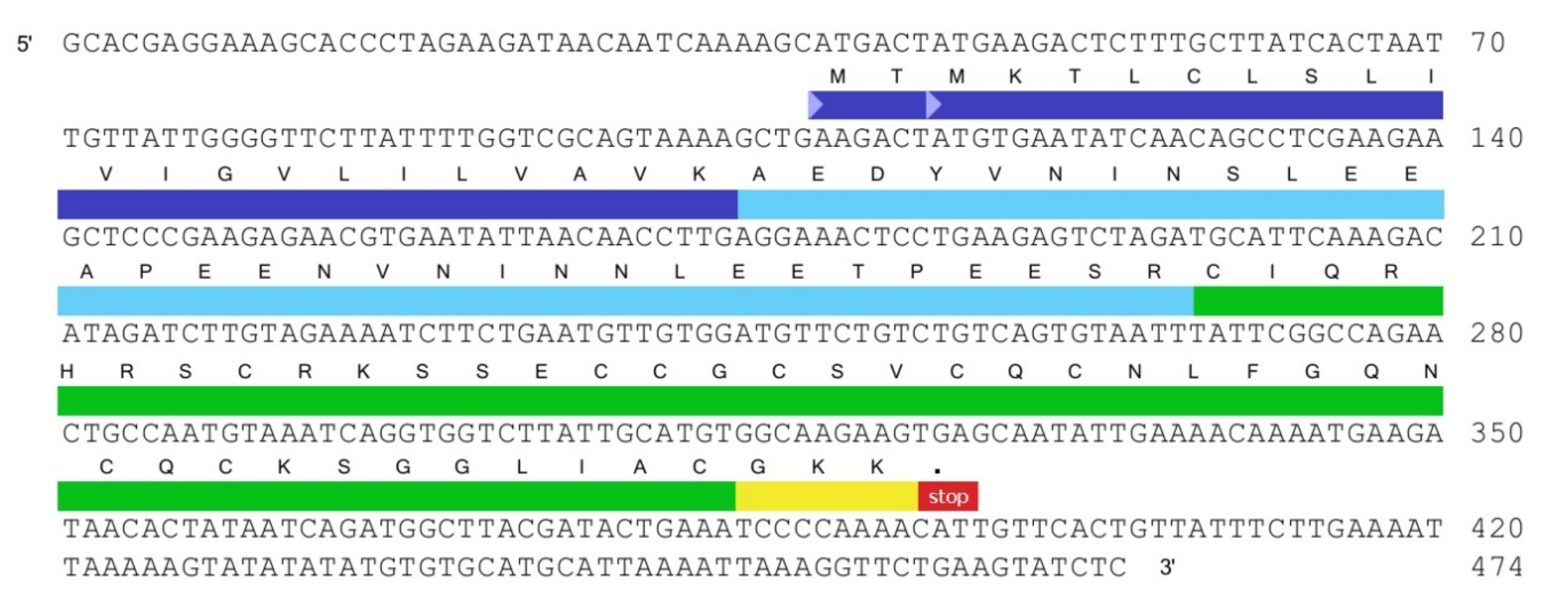
2.1. Tbo-IT2 Primary Structure
2.1.1. Transcriptome Analysis of cDNA Sequencing Data from the Venom Glands of Tibellus Oblongus
2.1.2. Tbo-IT2 Precursor Determination
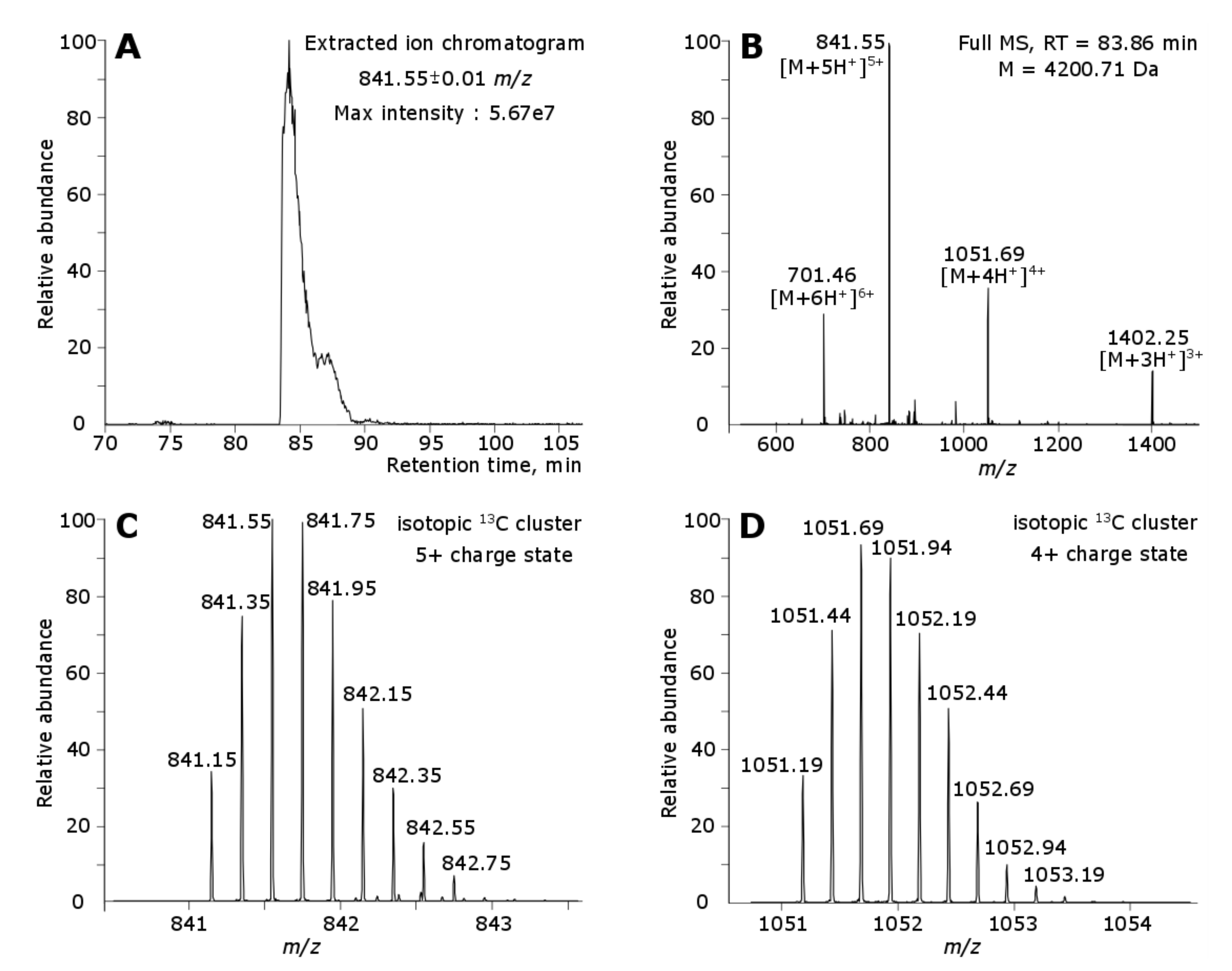
2.2. Identification of Tbo-IT2 in the Spider Venom
2.2.1. Proteomic Experiments Overview
2.2.2. Proteomic Identification of Tbo-IT2
2.3. Production of Recombinant Peptide
2.4. Study of the Biological Activity of Recombinant Tbo-IT2
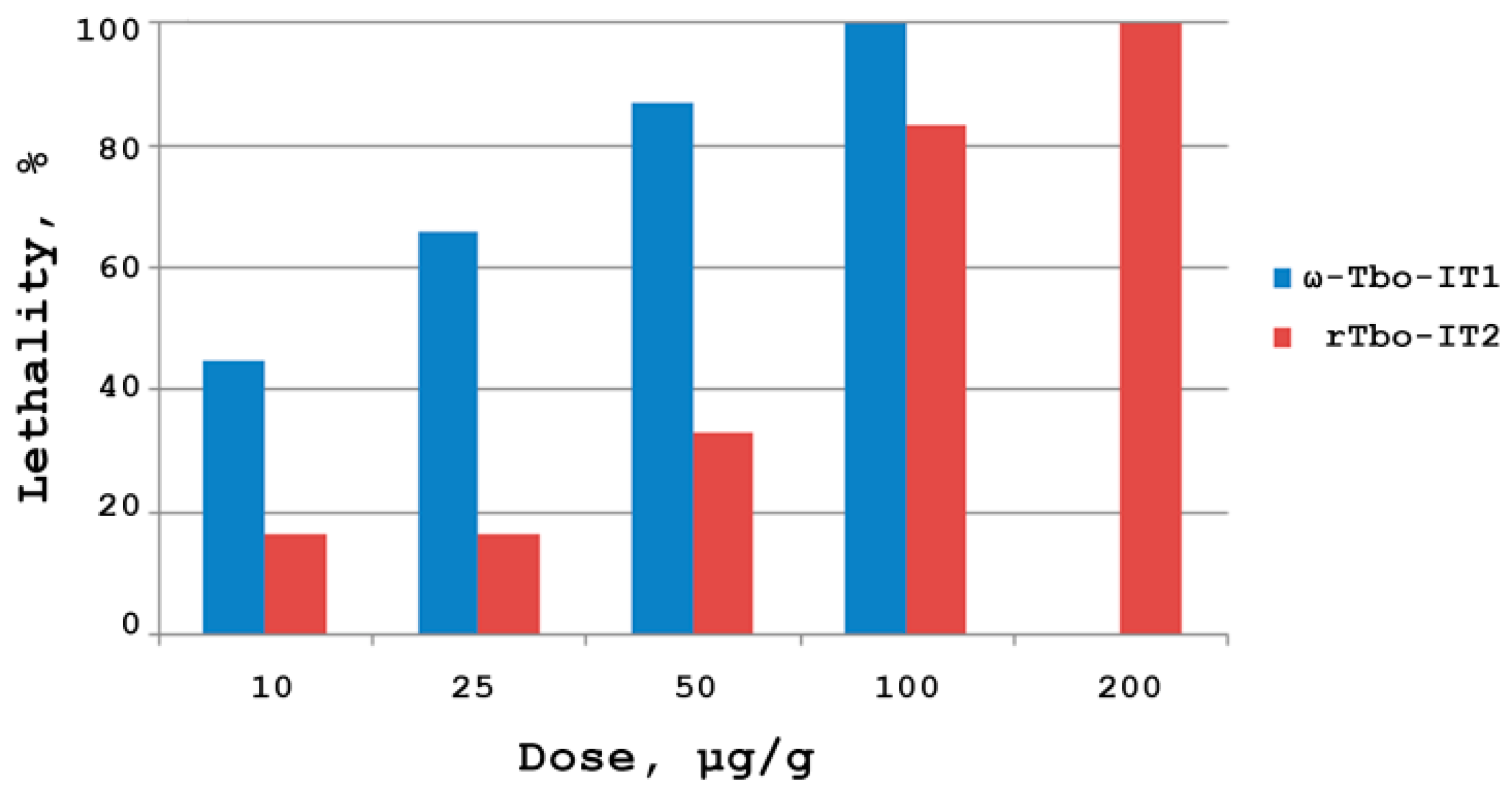
2.4.1. Insectotoxicity
2.4.2. Action on Neuronal Receptors
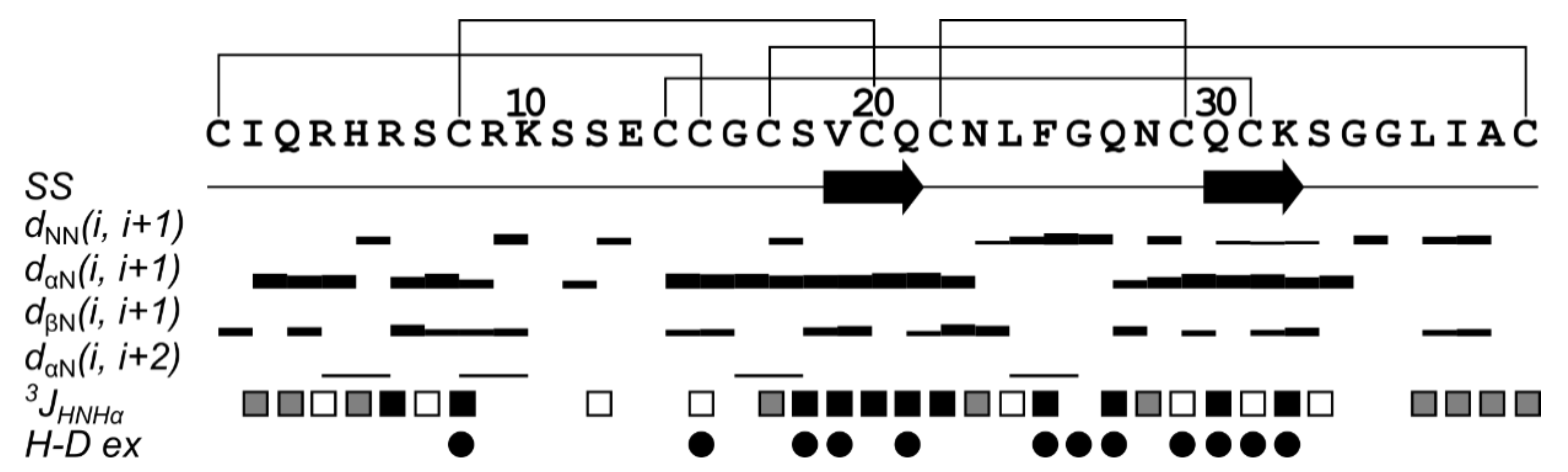
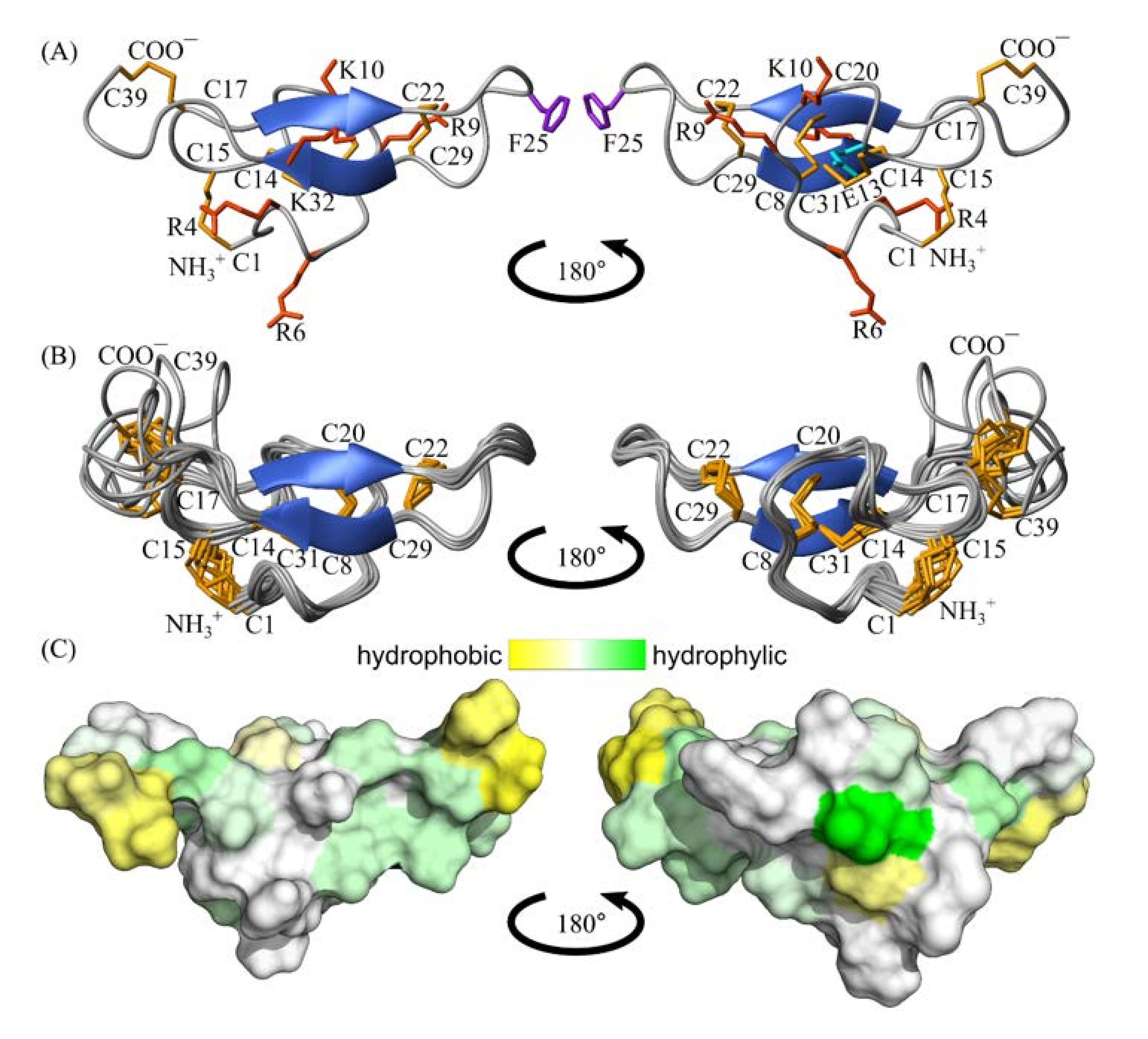
2.5. Spatial Structure of Tbo-IT2
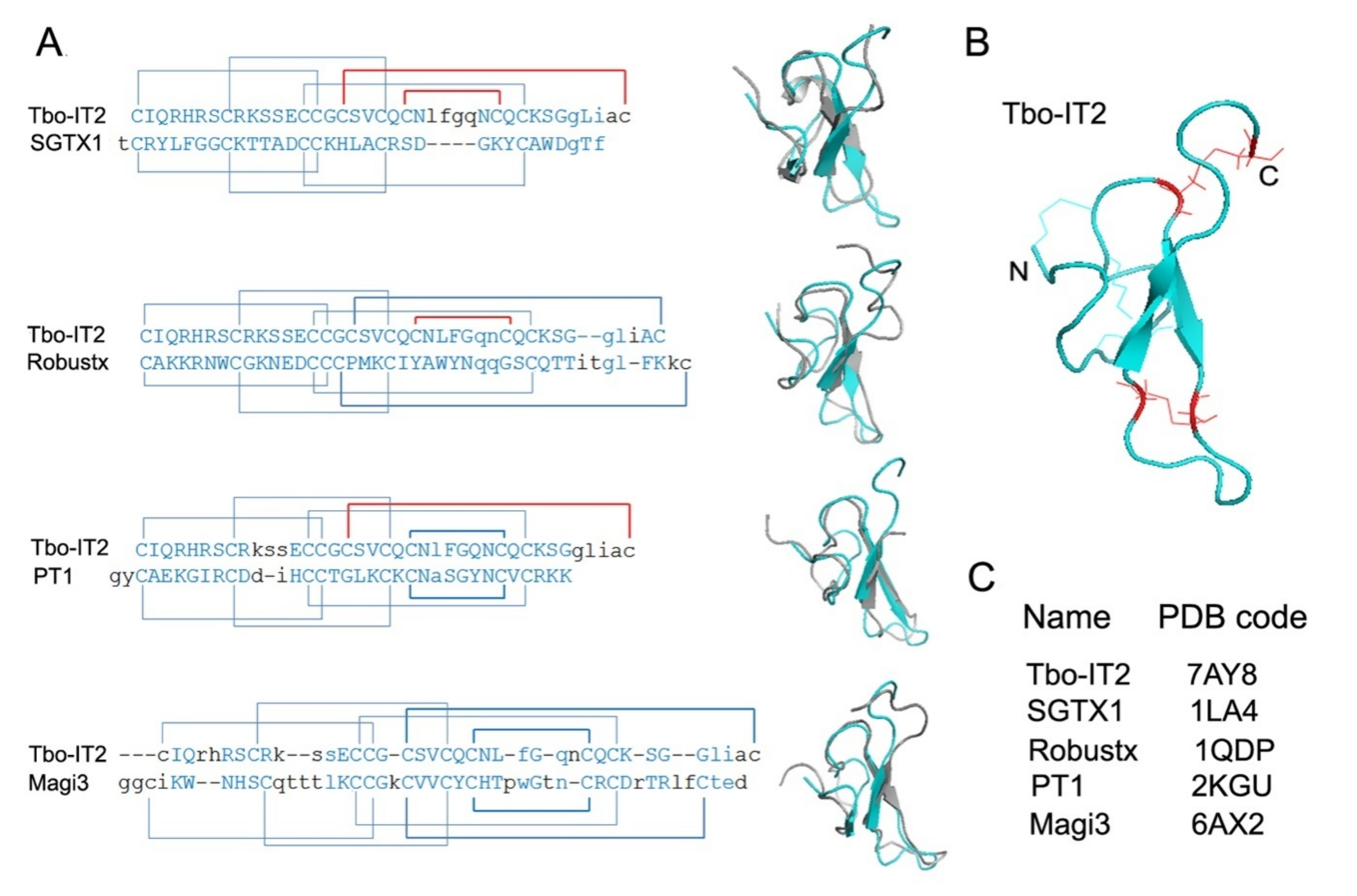
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animal Handling
5.2. EST Library Construction, Sequencing, and Data Processing
5.3. Venom Peptide Fraction Purification
5.4. Peptide Digestion
5.5. LC-MS/MS Analysis of Peptide Digests
5.6. LC-MS Analysis of Intact Toxins
5.7. Proteomics Data Analysis
5.8. Gene Construction and Recombinant Analog Production
5.9. Insectotoxicity Test
5.10. Activity Testing on Expressed Channels
5.11. NMR Spectroscopy and Calculation of Spatial Structure
5.12. Computation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Langenegger, N.; Nentwig, W.; Kuhn-Nentwig, L. Spider venom: Components, modes of action, and novel strategies in transcriptomic and proteomic analyses. Toxins 2019, 11, 611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gremski, L.H.; Da Justa, H.C.; Da Silva, T.P.; Polli, N.L.C.; Antunes, B.C.; Minozzo, J.C.; Wille, A.C.M.; Senff-Ribeiro, A.; Arni, R.K.; Veiga, S.S. Forty years of the description of brown spider venom phospholipases-D. Toxins 2020, 12, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ushkaryov, Y. α-Latrotoxin: From structure to some functions. Toxicon 2002, 40, 1–5. [Google Scholar] [CrossRef]
- Dongo, Y.; Cardoso, F.C.; Lewis, R.J. Spider knottin pharmacology at voltage-gated sodium channels and their potential to modulate pain Pathways. Toxins 2019, 11, 626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, F.C.; Lewis, R.J. Structure-function and therapeutic potential of spider venom-derived cysteine knot peptides targeting sodium channels. Front. Pharm. 2019, 10, 366. [Google Scholar] [CrossRef]
- Jiménez-Vargas, J.M.; Possani, L.D.; Luna-Ramírez, K. Arthropod toxins acting on neuronal potassium channels. Neuropharmacology 2017, 127, 139–160. [Google Scholar] [CrossRef]
- Norton, R.S.; Chandy, K.G. Venom-derived peptide inhibitors of voltage-gated potassium channels. Neuropharmacology 2017, 127, 124–138. [Google Scholar] [CrossRef]
- Bourinet, E.; Zamponi, G.W. Block of voltage-gated calcium channels by peptide toxins. Neuropharmacology 2017, 127, 109–115. [Google Scholar] [CrossRef]
- Sousa, S.R.; Vetter, I.; Lewis, R.J. Venom peptides as a rich source of Cav2.2 channel blockers. Toxins 2013, 5, 286–314. [Google Scholar] [CrossRef] [Green Version]
- Cristofori-Armstrong, B.; Rash, L.D. Acid-sensing ion channel (ASIC) structure and function: Insights from spider, snake and sea anemone venoms. Neuropharmacology 2017, 127, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Geron, M.; Hazan, A.; Priel, A. Animal toxins providing insights into TRPV1 activation mechanism. Toxins 2017, 9, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonello, R.; Fusi, C.; Materazzi, S.; Marone, I.M.; De Logu, F.; Benemei, S.; Gonçalves, M.C.; Coppi, E.; Castro-Junior, C.J.; Gomez, M.V.; et al. The peptide Phα1β, from spider venom, acts as a TRPA1 channel antagonist with antinociceptive effects in mice. Br. J. Pharm. 2017, 174, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Grishin, E.V.; Savchenko, G.A.; Vassilevski, A.A.; Korolkova, Y.V.; Boychuk, Y.A.; Viatchenko-Karpinski, V.Y.; Nadezhdin, K.D.; Arseniev, A.S.; Pluzhnikov, K.A.; Kulyk, V.B.; et al. Novel peptide from spider venom inhibits P2X3 receptors and inflammatory pain. Ann. Neurol. 2010, 67, 680–683. [Google Scholar] [CrossRef] [PubMed]
- Oparin, P.B.; Nadezhdin, K.D.; Berkut, A.A.; Arseniev, A.S.; Grishin, E.V.; Vassilevski, A.A. Structure of purotoxin-2 from Wolf spider: Modular design and membrane-Assisted mode of action in arachnid toxins. Biochem. J. 2016, 473, 3113–3126. [Google Scholar] [CrossRef] [PubMed]
- King, G.F. Tying pest insects in knots: The deployment of spider-venom-derived knottins as bioinsecticides. Pest. Manag. Sci. 2019, 75, 2437–2445. [Google Scholar] [CrossRef]
- Wu, T.; Wang, M.; Wu, W.; Luo, Q.; Jiang, L.; Tao, H.; Deng, M. Spider venom peptides as potential drug candidates due to their anticancer and antinociceptive activities. J. Venom. Anim. Toxins Incl. Trop. Dis. 2019, 25, e146318. [Google Scholar] [CrossRef]
- Mikov, A.N.; Fedorova, I.M.; Potapieva, N.N.; Maleeva, E.E.; Andreev, Y.A.; Zaitsev, A.V.; Kim, K.K.; Bocharov, E.V.; Bozin, T.N.; Altukhov, D.A.; et al. ω-Tbo-IT1-new inhibitor of insect calcium channels isolated from spider venom. Sci. Rep. 2015, 5, 17232. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, S.A.; Vassilevski, A.A.; Grishin, E.V. Secreted protein and peptide biosynthesis: Precursor structures and processing mechanisms. In Protein Biosynthesis; Esterhouse, T.E., Petrinos, L.B., Eds.; Nova Biomedical Books: New York, NY, USA, 2009; pp. 225–248. [Google Scholar]
- Kozlov, S.A.; Grishin, E.V. The universal algorithm of maturation for secretory and excretory protein precursors. Toxicon 2007, 49, 721–726. [Google Scholar] [CrossRef]
- Levitsky, L.I.; Ivanov, M.V.; Lobas, A.A.; Bubis, J.A.; Tarasova, I.A.; Solovyeva, E.M.; Pridatchenko, M.L.; Gorshkov, M.V. IdentiPy: An extensible search engine for protein identification in shotgun proteomics. J. Proteome Res. 2018, 17, 2249–2255. [Google Scholar] [CrossRef]
- Levitsky, L.I.; Klein, J.A.; Ivanov, M.V.; Gorshkov, M.V. Pyteomics 4.0: Five years of development of a python proteomics framework. J. Proteome Res. 2019, 18, 709–714. [Google Scholar] [CrossRef]
- LaVallie, E.R.; DiBlasio, E.A.; Kovacic, S.; Grant, K.L.; Schendel, P.F.; McCoy, J.M. A thioredoxin gene fusion expression system that circumvents inclusion body formation in the E. coli cytoplasm. Bio Technol. 1993, 11, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Andreev, Y.A.; Kozlov, S.A.; Vassilevski, A.A.; Grishin, E.V. Cyanogen bromide cleavage of proteins in salt and buffer solutions. Anal. Biochem. 2010, 407, 144–146. [Google Scholar] [CrossRef] [PubMed]
- White, S.H.; Wimley, W.C. Hydrophobic interactions of peptides with membrane interfaces. Biochim. Biophys. Acta Rev. Biomembr. 1998, 1376, 339–352. [Google Scholar] [CrossRef] [Green Version]
- Quistad, G.B.; Skinner, W.S. Isolation and sequencing of insecticidal peptides from the primitive hunting spider, Plectreurys tristis (Simon)—PubMed. J. Biol. Chem. 1994, 269, 11098–11101. [Google Scholar]
- Branton, W.D.; Kolton, L.; Jan, Y.N.; Jan, L.Y. Neurotoxins from Plectreurys spider venom are potent presynaptic blockers in Drosophila. J. Neurosci. 1987, 7, 4195–4200. [Google Scholar] [CrossRef] [Green Version]
- Titaux-Delgado, G.; Carrillo, E.; Mendoza, A.; Mayorga-Flores, M.; Escobedo-González, F.C.; Cano-Sánchez, P.; López-Vera, E.; Corzo, G.; del Rio-Portilla, F. Successful refolding and NMR structure of rMagi3: A disulfide-rich insecticidal spider toxin. Protein Sci. 2018, 27, 692–701. [Google Scholar] [CrossRef]
- Andreev, Y.A.; Osmakov, D.I.; Koshelev, S.G.; Maleeva, E.E.; Logashina, Y.A.; Palikov, V.A.; Palikova, Y.A.; Dyachenko, I.A.; Kozlov, S.A. Analgesic activity of acid-Sensing ion channel 3 (ASIC3) inhibitors: Sea anemones peptides Ugr9-1 and APETx2 versus low molecular weight compounds. Mar. Drugs 2018, 16, 500. [Google Scholar] [CrossRef] [Green Version]
- Logashina, Y.A.; Solstad, R.G.; Mineev, K.S.; Korolkova, Y.V.; Mosharova, I.V.; Dyachenko, I.A.; Palikov, V.A.; Palikova, Y.A.; Murashev, A.N.; Arseniev, A.S.; et al. New disulfide-stabilized fold provides sea anemone peptide to exhibit both antimicrobial and TRPA1 potentiating properties. Toxins 2017, 9, 154. [Google Scholar] [CrossRef] [Green Version]
- Logashina, Y.A.; Mosharova, I.V.; Korolkova, Y.V.; Shelukhina, I.V.; Dyachenko, I.A.; Palikov, V.A.; Palikova, Y.A.; Murashev, A.N.; Kozlov, S.A.; Stensvåg, K.; et al. Peptide from sea anemone metridium senile affects transient receptor potential ankyrin-repeat 1 (TRPA1) function and produces analgesic effect. J. Biol. Chem. 2017, 292, 2992–3004. [Google Scholar] [CrossRef] [Green Version]
- Daly, N.L.; Craik, D.J. Bioactive cystine knot proteins. Curr. Opin. Chem. Biol. 2011, 15, 362–368. [Google Scholar] [CrossRef]
- Mouhat, S.; Jouirou, B.; Mosbah, A.; De Waard, M.; Sabatier, J.M. Diversity of folds in animal toxins acting on ion channels. Biochem. J. 2004, 378, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Ferrat, G.; Bosmans, F.; Tytgat, J.; Pimentel, C.; Chagot, B.; Gilles, N.; Nakajima, T.; Darbon, H.; Corzo, G. Solution structure of two insect-specific spider toxins and their pharmacological interaction with the insect voltage-gated Na+ channel. Proteins Struct. Funct. Genet. 2005, 59, 368–379. [Google Scholar] [CrossRef] [PubMed]
- McNulty, J.C.; Jackson, P.J.; Thompson, D.A.; Chai, B.; Gantz, I.; Barsh, G.S.; Dawson, P.E.; Millhauser, G.L. Structures of the agouti signaling protein. J. Mol. Biol. 2005, 346, 1059–1070. [Google Scholar] [CrossRef] [PubMed]
- Bolin, K.A.; Anderson, D.J.; Trulson, J.A.; Thompson, D.A.; Wilken, J.; Kent, S.B.H.; Gantz, I.; Millhauser, G.L. NMR structure of a minimized human agouti related protein prepared by total chemical synthesis. FEBS Lett. 1999, 451, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Krissinel, E.; Henrick, K. Secondary-structure matching (SSM), a new tool for fast protein structure alignment in three dimensions. Acta Cryst. Sect. D Biol. Cryst. 2004, 60, 2256–2268. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Kim, S.; Roh, S.H.; Endoh, H.; Kodera, Y.; Maeda, T.; Kohno, T.; Wang, J.M.; Swartz, K.J.; Kim, J. Il Solution structure and functional characterization of SGTxl, a modifier of Kv2.1 channel gating. Biochemistry 2004, 43, 890–897. [Google Scholar] [CrossRef] [PubMed]
- Sigoillot, M.; Brockhoff, A.; Neiers, F.; Poirier, N.; Belloir, C.; Legrand, P.; Charron, C.; Roblin, P.; Meyerhof, W.; Briand, L. The crystal structure of gurmarin, a sweet taste–suppressing protein: Identification of the amino acid residues essential for inhibition. Chem. Senses 2018, 43, 635–643. [Google Scholar] [CrossRef]
- Escoubas, P.; Bernard, C.; Lambeau, G.; Lazdunski, M.; Darbon, H. Recombinant production and solution structure of PcTx1, the specific peptide inhibitor of ASIC1a proton-gated cation channels. Protein Sci. 2003, 12, 1332–1343. [Google Scholar] [CrossRef] [Green Version]
- Pallaghy, P.K.; Alewood, D.; Alewood, P.F.; Norton, R.S. Solution structure of robustoxin, the lethal neurotoxin from the funnel-web spider Atrax robustus. FEBS Lett. 1997, 419, 191–196. [Google Scholar] [CrossRef] [Green Version]
- King, G.F.; Gentz, M.C.; Escoubas, P.; Nicholson, G.M. A rational nomenclature for naming peptide toxins from spiders and other venomous animals. Toxicon 2008, 52, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Prudencio, G.; Possani, L.D.; Becerril, B.; Ortiz, E. The dual α-amidation system in scorpion venom glands. Toxins 2019, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzmenkov, A.I.; Grishin, E.V.; Vassilevski, A.A. Diversity of potassium channel ligands: Focus on scorpion toxins. Biochemistry 2015, 80, 1764–1799. [Google Scholar] [CrossRef] [PubMed]
- Coelho, V.A.; Cremonez, C.M.; Anjolette, F.A.P.; Aguiar, J.F.; Varanda, W.A.; Arantes, E.C. Functional and structural study comparing the C-terminal amidated β-neurotoxin Ts1 with its isoform Ts1-G isolated from Tityus serrulatus venom. Toxicon 2014, 83, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Benkhadir, K.; Kharrat, R.; Cestèle, S.; Mosbah, A.; Rochat, H.; El Ayeb, M.; Karoui, H. Molecular cloning and functional expression of the alpha-scorpion toxin BotIII: Pivotal role of the C-terminal region for its interaction with voltage-dependent sodium channels. Peptides 2004, 25, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez De La Vega, R.C.; Possani, L.D. Overview of scorpion toxins specific for Na+ channels and related peptides: Biodiversity, structure-function relationships and evolution. Toxicon 2005, 46, 831–844. [Google Scholar] [CrossRef]
- Cardoso, F.C.; Dekan, Z.; Rosengren, K.J.; Erickson, A.; Vetter, I.; Deuis, J.R.; Herzig, V.; Alewood, P.F.; King, G.F.; Lewis, R.J. Identification and characterization of ProTx-III [μ-TRTX-Tp1a], a new voltage-gated sodium channel inhibitor from venom of the tarantula Thrixopelma pruriens. Mol. Pharm. 2015, 88, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Yang, G.; Li, B.; Chi, C.; Wu, X. Cloning, co-expression with an amidating enzyme, and activity of the scorpion toxin BmK ITa1 cDNA in insect cells. Appl. Biochem. Biotechnol. Part. B Mol. Biotechnol. 2003, 24, 21–26. [Google Scholar] [CrossRef]
- Sermadiras, I.; Revell, J.; Linley, J.E.; Sandercock, A.; Ravn, P. Recombinant expression and in Vitro characterisation of active huwentoxin-IV. PLoS ONE 2013, 8, e83202. [Google Scholar] [CrossRef] [Green Version]
- Devaux, C.; Knibiehler, M.; Defendini, M.-L.; Mabrouk, K.; Rochat, H.; Van Rietschoten, J.; Baty, D.; Granier, C. Recombinant and chemical derivatives of apamin: Implication of post-transcriptional c-terminal amidation of apamin in biological activity. Eur. J. Biochem. 1995, 231, 544–550. [Google Scholar] [CrossRef]
- Sabatier, J.M.; Zerrouk, H.; Darbon, H.; Mabrouk, K.; Benslimane, A.; Rochat, H.; Martin-Eauclaire, M.F.; Van Rietschoten, J. P05, a new leiurotoxin i-like scorpion toxin: Synthesis and structure-activity relationships of the α-amidated analog, a ligand of Ca2+-activated K+ channels with increased affinity. Biochemistry 1993, 32, 2763–2770. [Google Scholar] [CrossRef]
- Saucedo, A.L.; Del Rio-Portilla, F.; Picco, C.; Estrada, G.; Prestipino, G.; Possani, L.D.; Delepierre, M.; Corzo, G. Solution structure of native and recombinant expressed toxin CssII from the venom of the scorpion Centruroides suffusus suffusus, and their effects on Nav1.5 Sodium channels. Biochim. Biophys. Acta Proteins Proteom. 2012, 1824, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Legros, C.; Céard, B.; Vacher, H.; Marchot, P.; Bougis, P.E.; Martin-Eauclaire, M.F. Expression of the standard scorpion alpha-toxin AaH II and AaH II mutants leading to the identification of some key bioactive elements. Biochim. Biophys. Acta Gen. Subj. 2005, 1723, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Estrada, G.; Restano-Cassulini, R.; Ortiz, E.; Possani, L.D.; Corzo, G. Addition of positive charges at the C-terminal peptide region of CssII, a mammalian scorpion peptide toxin, improves its affinity for sodium channels Nav1.6. Peptides 2011, 32, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Corzo, G.; Escoubas, P.; Stankiewicz, M.; Pelhate, M.; Kristensen, C.P.; Nakajima, T. Isolation, synthesis and pharmacological characterization of δ-palutoxins IT, novel insecticidal toxins from the spider Paracoelotes luctuosus (Amaurobiidae). Eur. J. Biochem. 2000, 267, 5783–5795. [Google Scholar] [CrossRef]
- Ostrow, K.L.; Mammoser, A.; Suchyna, T.; Sachs, F.; Oswald, R.; Kubo, S.; Chino, N.; Gottlieb, P.A. cDNA sequence and in vitro folding of GsMTx4, a specific peptide inhibitor of mechanosensitive channels. Toxicon 2003, 42, 263–274. [Google Scholar] [CrossRef]
- Vassilevski, A.A.; Kozlov, S.A.; Grishin, E.V. Molecular diversity of spider venom. Biochemistry 2009, 74, 1505–1534. [Google Scholar] [CrossRef]
- Leisy, D.J.; Alto, P.; Quistad, G.B.; View, M.; Skinner, W.S.; Valley, P. Insecticidal Plectoxins from Plectreurystristis. U.S. Patent No. 6,265,376 Bl, 24 July 2001. [Google Scholar]
- Corzo, G.; Gilles, N.; Satake, H.; Villegas, E.; Dai, L.; Nakajima, T.; Haupt, J. Distinct primary structures of the major peptide toxins from the venom of the spider Macrothele gigas that bind to sites 3 and 4 in the sodium channel. FEBS Lett. 2003, 547, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, G.M.; Walsh, R.; Little, M.J.; Tyler, M.I. Characterisation of the effects of robustoxin, the lethal neurotoxin from the Sydney funnel-web spider Atrax robustus, on sodium channel activatiom and inactivation. Pflug. Arch. Eur. J. Physiol. 1998, 436, 117–126. [Google Scholar] [CrossRef]
- Herzig, V.; Sunagar, K.; Wilson, D.T.R.; Pineda, S.S.; Israel, M.R.; Dutertre, S.; McFarland, B.S.; Undheim, E.A.B.; Hodgson, W.C.; Alewood, P.F.; et al. Australian funnel-web spiders evolved human-lethal δ-hexatoxins for defense against vertebrate predators. Proc. Natl. Acad. Sci. USA 2020, 117, 24920–24928. [Google Scholar] [CrossRef]
- Chen, X.; Kalbacher, H.; Gründer, S. Interaction of acid-sensing ion channel (ASIC) 1 with the tarantula toxin psalmotoxin 1 is state dependent. J. Gen. Physiol. 2006, 127, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Chambers, M.C.; MacLean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef]
- Korolkova, Y.; Makarieva, T.; Tabakmakher, K.; Shubina, L.; Kudryashova, E.; Andreev, Y.; Mosharova, I.; Lee, H.S.; Lee, Y.J.; Kozlov, S. Marine cyclic guanidine alkaloids monanchomycalin B and urupocidin a act as inhibitors of TRPV1, TRPV2 and TRPV3, but not TRPA1 receptors. Mar. Drugs 2017, 15, 87. [Google Scholar] [CrossRef] [Green Version]
- Osmakov, D.I.; Koshelev, S.G.; Lyukmanova, E.N.; Shulepko, M.A.; Andreev, Y.A.; Illes, P.; Kozlov, S.A. Multiple modulation of acid-sensing ion channel 1a by the alkaloid daurisoline. Biomolecules 2019, 9, 336. [Google Scholar] [CrossRef] [Green Version]
- Cremonez, C.M.; Maiti, M.; Peigneur, S.; Cassoli, J.S.; Dutra, A.A.A.; Waelkens, E.; Lescrinier, E.; Herdewijn, P.; de Lima, M.E.; Pimenta, A.M.C.; et al. Structural and functional elucidation of peptide TS11 shows evidence of a novel subfamily of scorpion venom toxins. Toxins 2016, 8, 288. [Google Scholar] [CrossRef] [Green Version]
- Wuthrich, K. NMR of Proteins and Nucleic Acids; Wiley: New York, NY, USA, 1986. [Google Scholar]
- Güntert, P.; Buchner, L. Combined automated NOE assignment and structure calculation with CYANA. J. Biomol. NMR 2015, 62, 453–471. [Google Scholar] [CrossRef]
- Delaglio, F.; Grzesiek, S.; Vuister, G.W.; Zhu, G.; Pfeifer, J.; Bax, A. NMRPipe: A multidimensional spectral processing system based on UNIX pipes. J. Biomol. NMR 1995, 6, 277–293. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | PSMs | Peptide | AA-from | AA-to | Missed Cleavages |
|---|---|---|---|---|---|
| trypsin | 11 | KSSECCGCSVCQCNLFGQNCQCK | 10 | 32 | 1 |
| trypsin | 8 | SSECCGCSVCQCNLFGQNCQCK | 11 | 32 | 0 |
| GluC | 12 | CCGCSVCQCNLFGQNCQCKSGGLIAC-NH2 | 14 | 39 | 0 |
| Distance and Angle Restraints | ||
|---|---|---|
| Total NOEs | 361 | |
| intraresidual | 94 | |
| interresidual | 267 | |
| sequential(|i − j| = 1) | 78 | |
| medium range(1 < |i − j| ≤ 4) | 48 | |
| long-range(|i − j| > 4) | 141 | |
| Hydrogen bond restraints(upper/lower) | 23/23 | |
| S-S bond restraints(upper/lower) | 15/15 | |
| J-couplings | ||
| 3JHNHα | 29 | |
| 3JHαHβ | 42 | |
| Angles | ||
| φ | 38 | |
| Χ1 | 18 | |
| Total restraints/per residue | 484/12 | |
| Statistics of the obtained set of structures | ||
| CYANA target function | 1.64 ± 0.25 | |
| Restraints violations | ||
| distance(>0.2 Å) | 0 | |
| angle(>5°) | 2 | |
| RMSD(Å) | SS 1 | RR 2 |
| backbone | 0.18 ± 0.03 | 0.57 ± 0.11 |
| all heavy atoms | 0.76 ± 0.07 | 1.40 ± 0.17 |
| Ramachandran analysis * | ||
| %residues in most favored regions | 69.7 | |
| %residues in additional allowed regions | 30.3 | |
| %residues in generously allowed regions | 0.0 | |
| %residues in disallowed regions | 0.0 | |
| Peptides | PDB Codes | N Res | N Align | % Ident | Scoring | RMSD | |
|---|---|---|---|---|---|---|---|
| Q | Z | ||||||
| SGTX1 | 1la4:A | 34 | 33 | 21 | 0.63 | 6.2 | 1.68 |
| robustoxin | 1qdp:A | 42 | 38 | 26 | 0.60 | 5.8 | 2.06 |
| gurmarin 1 | 5oll:A | 35 | 30 | 20 | 0.54 | 4.6 | 1.43 |
| PT1 | 2kgu:A | 35 | 31 | 35 | 0.53 | 6.3 | 1.72 |
| PcTx1 | 1lmm:A | 40 | 33 | 21 | 0.52 | 4.1 | 1.77 |
| ASIP(80-132:Q115Y, S124Y) | 1y7j:A | 40 | 34 | 29 | 0.43 | 3.6 | 2.52 |
| AGRP(87-132) | 1hyk:A | 46 | 33 | 36 | 0.43 | 4.6 | 1.92 |
| Magi3 | 6ax2:A | 46 | 31 | 32 | 0.34 | 3.6 | 2.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korolkova, Y.; Maleeva, E.; Mikov, A.; Lobas, A.; Solovyeva, E.; Gorshkov, M.; Andreev, Y.; Peigneur, S.; Tytgat, J.; Kornilov, F.; et al. New Insectotoxin from Tibellus Oblongus Spider Venom Presents Novel Adaptation of ICK Fold. Toxins 2021, 13, 29. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010029
Korolkova Y, Maleeva E, Mikov A, Lobas A, Solovyeva E, Gorshkov M, Andreev Y, Peigneur S, Tytgat J, Kornilov F, et al. New Insectotoxin from Tibellus Oblongus Spider Venom Presents Novel Adaptation of ICK Fold. Toxins. 2021; 13(1):29. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010029
Chicago/Turabian StyleKorolkova, Yuliya, Ekaterina Maleeva, Alexander Mikov, Anna Lobas, Elizaveta Solovyeva, Mikhail Gorshkov, Yaroslav Andreev, Steve Peigneur, Jan Tytgat, Fedor Kornilov, and et al. 2021. "New Insectotoxin from Tibellus Oblongus Spider Venom Presents Novel Adaptation of ICK Fold" Toxins 13, no. 1: 29. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010029