Quantitative Proteomic Profiling of Fungal Growth, Development, and Ochratoxin A Production in Aspergillus ochraceus on High- and Low-NaCl Cultures


Abstract
:1. Introduction
2. Results
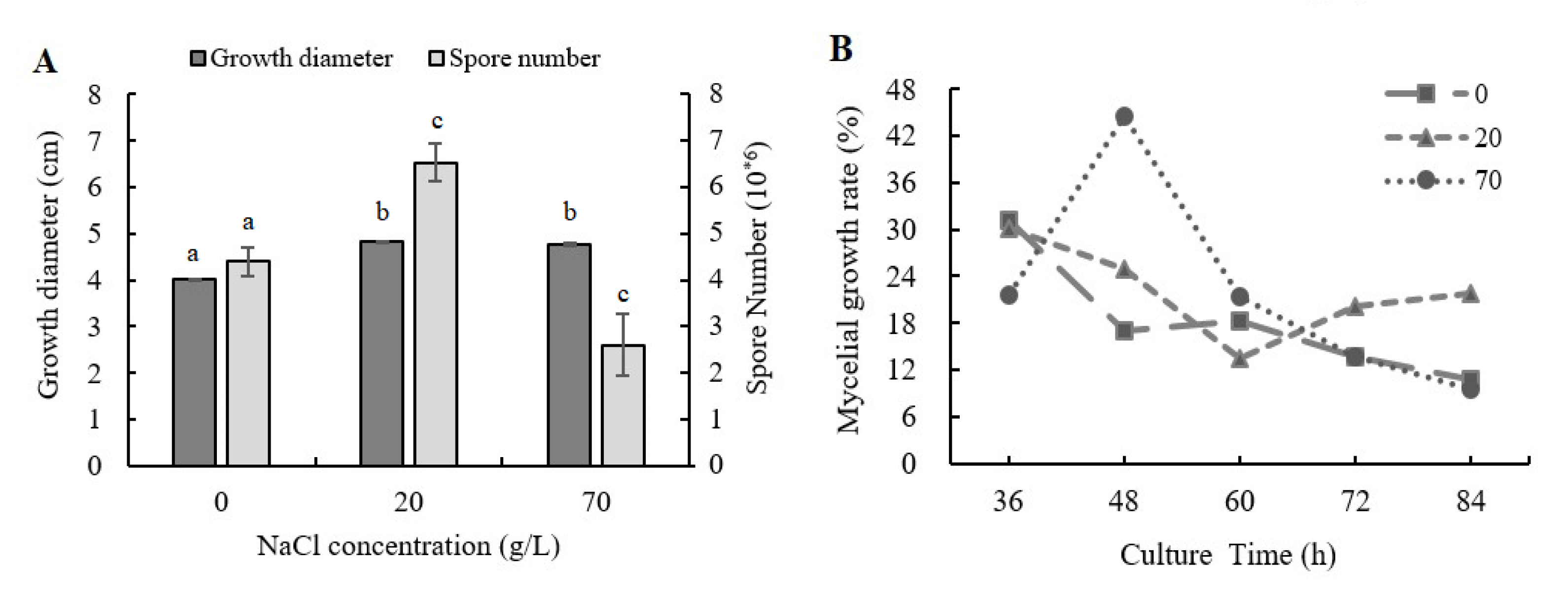
2.1. Influence of NaCl Addition on Mycelial Growth and Sporulation Ability of A. ochraceus
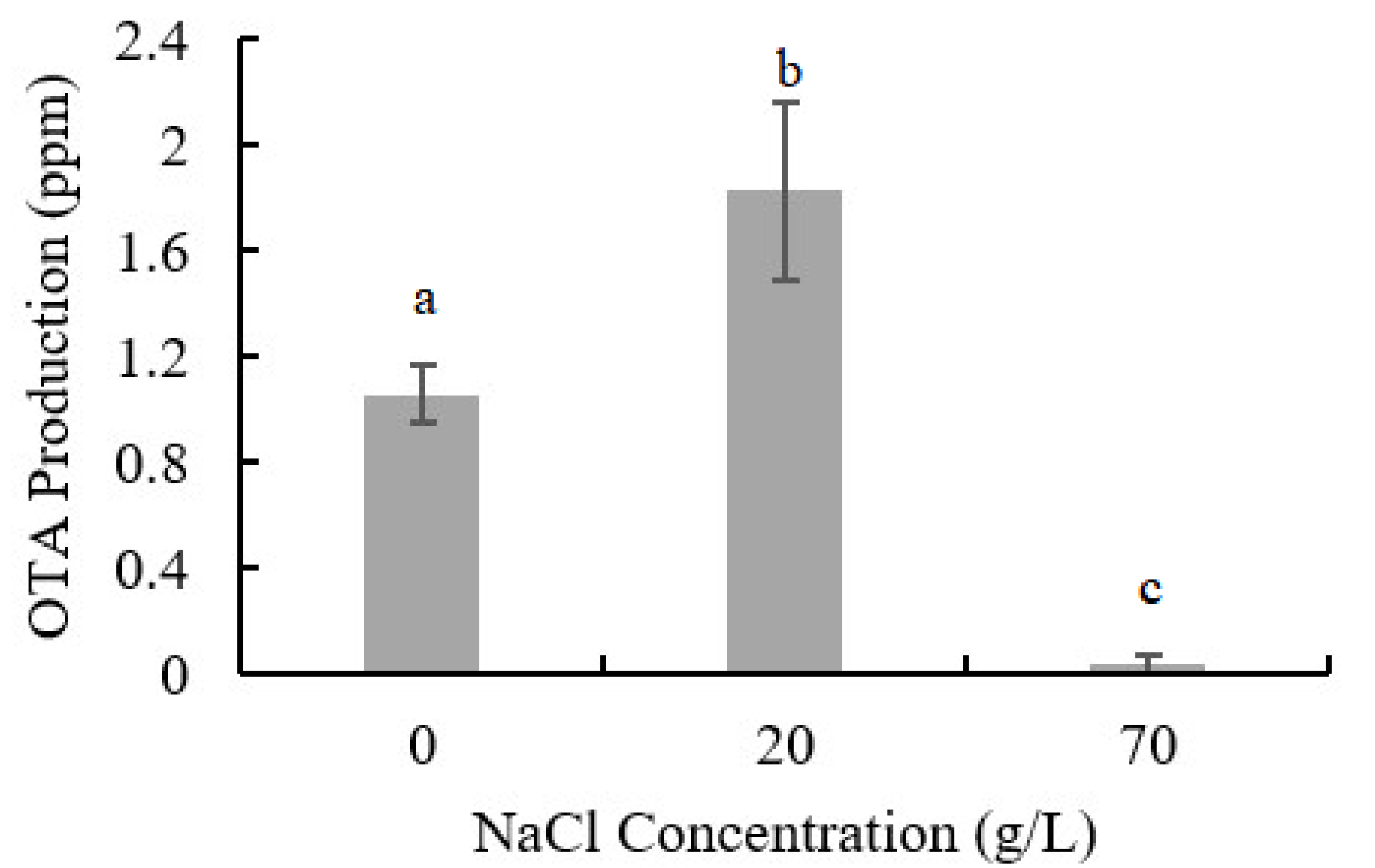
2.2. Influence of NaCl Addition on OTA Production of A. ochraceus
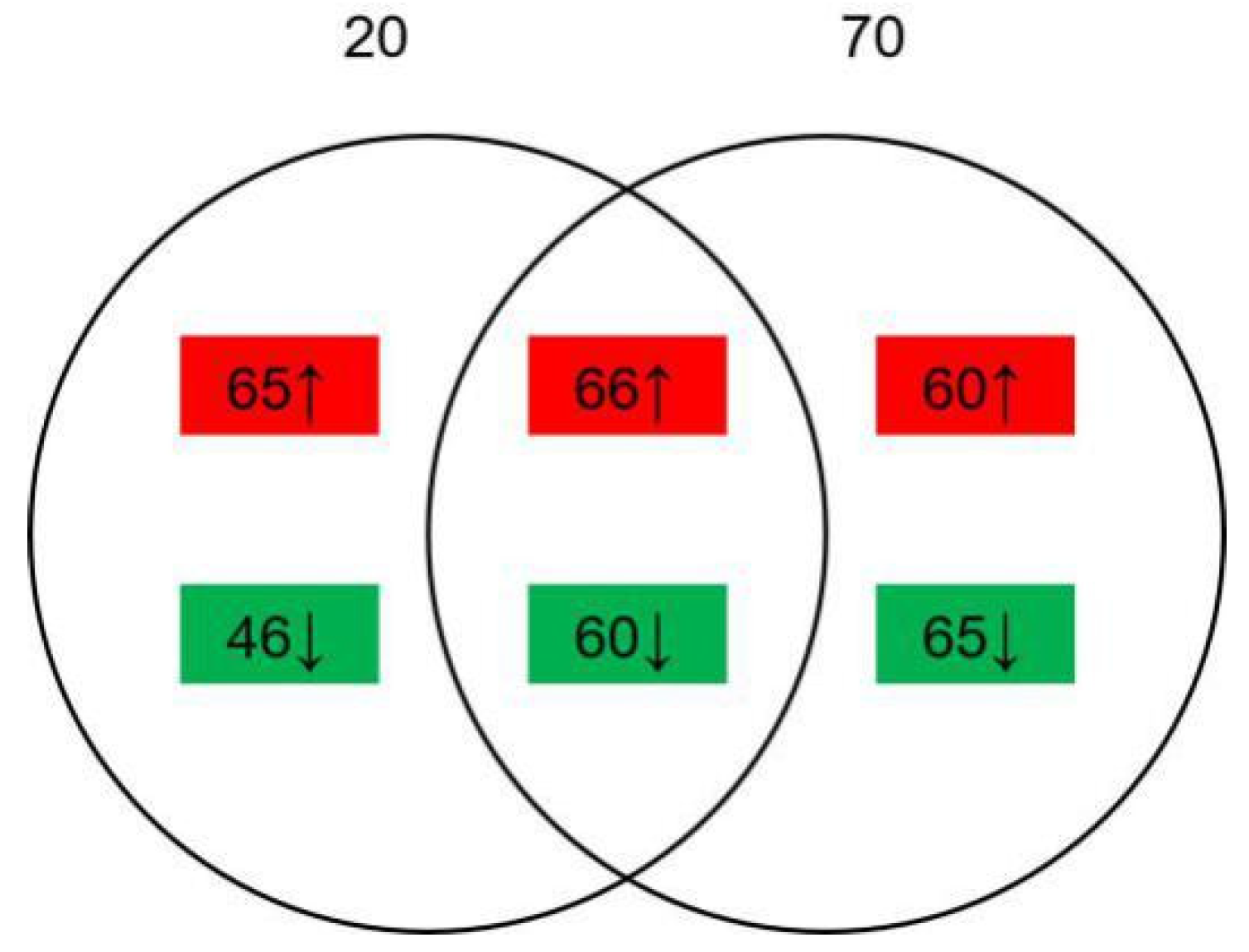
2.3. Alteration of Proteins with or without NaCl Addition
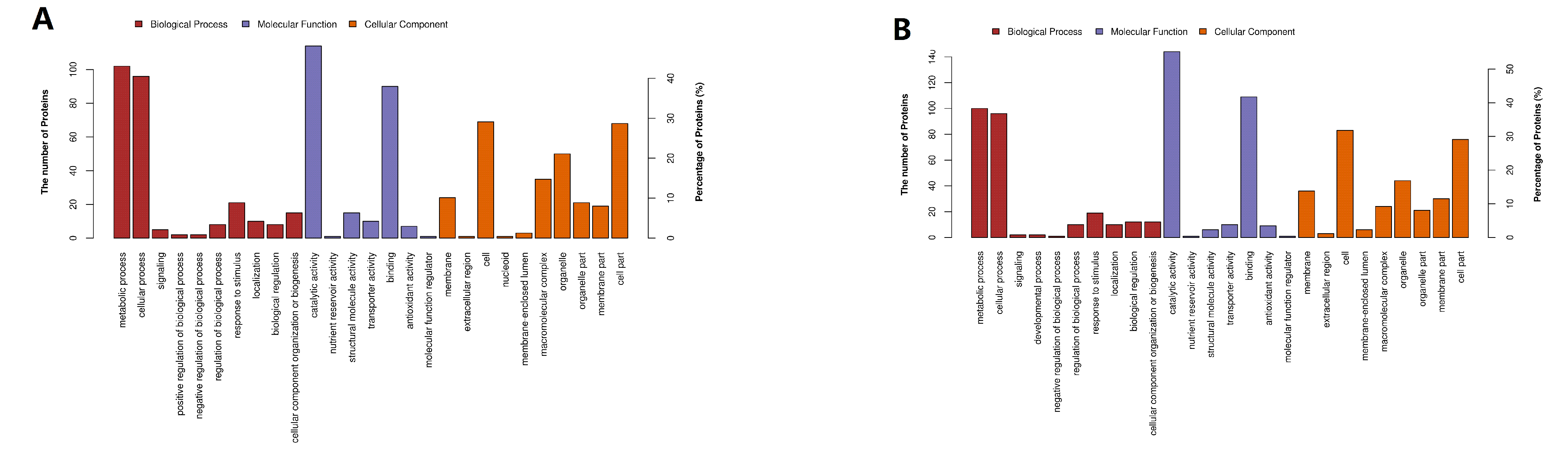
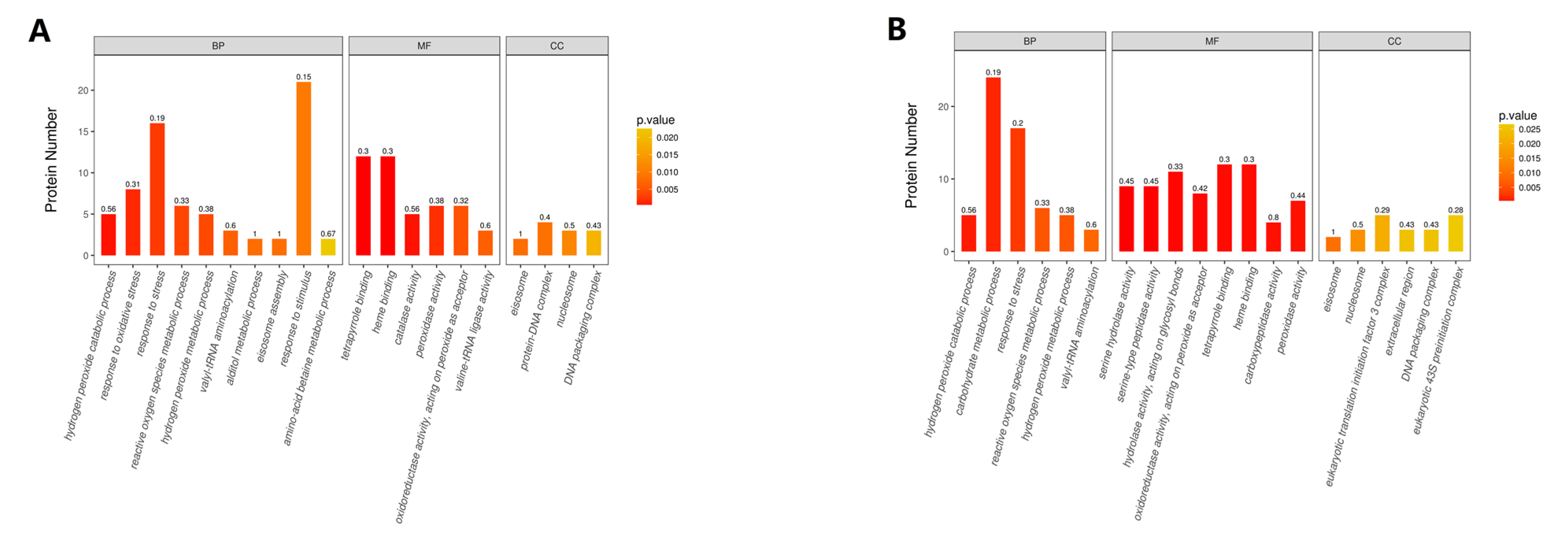
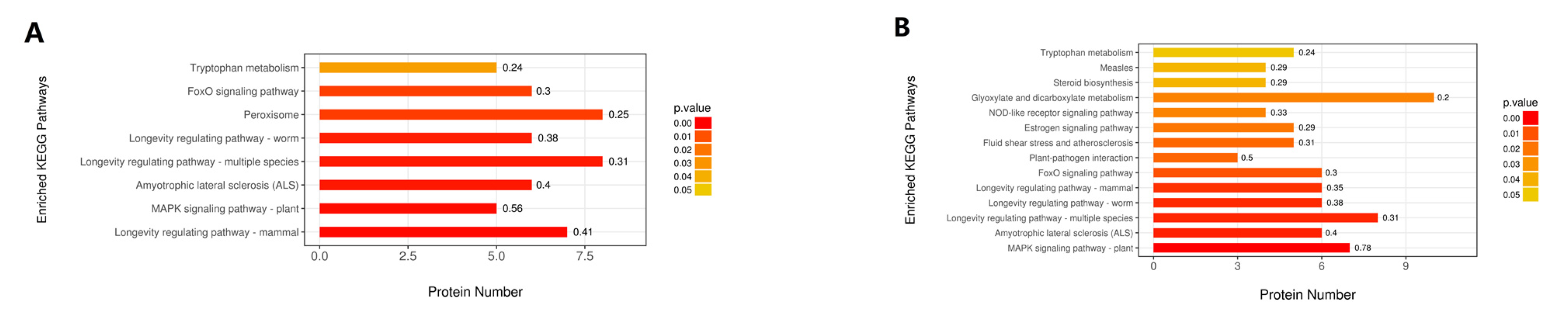
2.4. Hierarchical Clustering, Functional Classification, and Enrichment Analysis of DEPs
2.5. Classification of DEPs
3. Discussion
3.1. Fungal Growth Promoted by NaCl Addition
3.2. Energy Metabolism and Oxidative Stress Influenced by NaCl Addition
3.3. Protein Synthesis and Processing Repressed by NaCl Addition
3.4. Signal Transduction and Autophagy Activated by NaCl Addition
3.5. Fungal Secondary Metabolism Affected by NaCl Addition
4. Conclusions
5. Materials and Methods
5.1. Strains, Media, and Culture Conditions
5.2. Mycelial Growth and Conidia Production Assessments
5.3. OTA Detection
5.4. Proteomic Analysis
5.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Iacumin, L.; Milesi, S.; Pirani, S.; Comi, G.; Chiesa, L.M. Ochratoxigenic mold and Ochratoxin A in fermented sausages from different areas in Northern Italy: Occurrence, reduction or prevention with ozonated air. J. Food Saf. 2011, 31, 538–545. [Google Scholar] [CrossRef]
- Khaneghah, A.M.; Fakhri, Y.; Abdi, L.; Coppa, C.; Franco, L.T.; de Oliveira, C.A.F. The concentration and prevalence of ochratoxin A in coffee and coffee-based products: A global systematic review, meta-analysis and meta-regression. Fungal Biol. 2019, 123, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Liu, F.; Wang, Q.; Selvaraj, J.N.; Xing, F.; Zhao, Y.; Liu, Y. Ochratoxin A producing fungi, biosynthetic pathway and regulatory mechanisms. Toxins 2016, 8, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacumin, L.; Chiesa, L.; Boscolo, D.; Manzano, M.; Cantoni, C.; Orlic, S.; Comi, G. Moulds and ochratoxin A on surfaces of artisanal and industrial dry sausages. Food Microbiol. 2009, 26, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Peromingo, B.; Nunez, F.; Rodriguez, A.; Alia, A.; Andrade, M.J. Potential of yeasts isolated from dry-cured ham to control ochratoxin A production in meat models. Int. J. Food Microbiol. 2018, 268, 73–80. [Google Scholar] [CrossRef]
- Rodriguez, A.; Medina, A.; Cordoba, J.J.; Magan, N. The influence of salt (NaCl) on ochratoxin A biosynthetic genes, growth and ochratoxin A production by three strains of Penicillium nordicum on a dry-cured ham-based medium. Int. J. Food Microbiol. 2014, 178, 113–119. [Google Scholar] [CrossRef]
- Schmidt-Heydt, M.; Graf, E.; Stoll, D.; Geisen, R. The biosynthesis of ochratoxin A by Penicillium as one mechanism for adaptation to NaCl rich foods. Food Microbiol. 2012, 29, 233–241. [Google Scholar] [CrossRef]
- Álvarez, M.; Rodríguez, A.; Núñez, F.; Silva, A.; Andrade, M.J. In vitro antifungal effects of spices on ochratoxin A production and related gene expression in Penicillium nordicum on a dry-cured fermented sausage medium. Food Control 2020, 114, 107222. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Wu, F.; Liu, F.; Wang, Q.; Zhang, X.; Selvaraj, J.N.; Zhao, Y.; Xing, F.; Yin, W.B.; et al. A consensus Ochratoxin A biosynthetic pathway: Insights from the genome sequence of Aspergillus ochraceus and a comparative genomic analysis. Appl. Environ. Microbiol. 2018, 84, e01009-18. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.; Pitt, J.; Wu, F.; Gelderblom, W.; Wild, C.; Riley, R.; Baan, R. Improving public health through mycotoxin control. In International Agency for Research on Cancer; IARC WHO: Lyon, France, 2012. [Google Scholar]
- Ramos, A.J.; Labernia, N.; Marín, S.; Sanchis, V.; Magan, N. Effect of water activity and temperature on growth and ochratoxin production by three strains of Aspergillus ochraceus on a barley extract medium and on barley grains. Int. J. Food Microbiol. 1998, 44, 133–140. [Google Scholar] [CrossRef]
- Wang, Y.; Yan, H.; Neng, J.; Gao, J.; Yang, B.; Liu, Y. The Influence of NaCl and Glucose content on Growth and Ochratoxin A production by Aspergillus ochraceus, Aspergillus carbonarius and Penicillium nordicum. Toxins 2020, 12, 515. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Li, X.; Zhang, D.; Liu, Y. Comparison of protein differences between high- and low-quality goat and bovine parts based on iTRAQ technology. Food Chem. 2019, 289, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Canel, R.S.; Wagner, J.R.; Stenglein, S.A.; Ludemann, V. Indigenous filamentous fungi on the surface of Argentinean dry fermented sausages produced in Colonia Caroya (Cordoba). Int. J. Food Microbiol. 2013, 164, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.-L.; Yuan, J.-P.; Zhang, Y.; Xu, X.-M.; Wu, C.-F.; Peng, J.; Wang, J.-H. Fatty acid composition in ergosteryl esters and triglycerides from the fungus Ganoderma lucidum. J. Am. Oil Chem. Soc. 2013, 90, 1495–1502. [Google Scholar] [CrossRef]
- Seitz, L. Ergosterol as a measure of fungal growth. Phytopathology 1979, 69, 1202. [Google Scholar] [CrossRef] [Green Version]
- Gori, K.; Hebraud, M.; Chambon, C.; Mortensen, H.D.; Arneborg, N.; Jespersen, L. Proteomic changes in Debaryomyces hansenii upon exposure to NaCl stress. FEMS Yeast Res. 2007, 7, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Easlon, E.; Tsang, F.; Skinner, C.; Wang, C.; Lin, S.J. The malate-aspartate NADH shuttle components are novel metabolic longevity regulators required for calorie restriction-mediated life span extension in yeast. Genes Dev. 2008, 22, 931–944. [Google Scholar] [CrossRef] [Green Version]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxid. Med. Cell Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef]
- Frand, A.R.; Kaiser, C.A. Ero1p oxidizes protein disulfide isomerase in a pathway for disulfide bond formation in the endoplasmic reticulum. Mol. Cell 1999, 4, 469–477. [Google Scholar] [CrossRef]
- de la Cruz, J.; Gomez-Herreros, F.; Rodriguez-Galan, O.; Begley, V.; de la Cruz Munoz-Centeno, M.; Chavez, S. Feedback regulation of ribosome assembly. Curr. Genet. 2018, 64, 393–404. [Google Scholar] [CrossRef]
- Willi, J.; Kupfer, P.; Evequoz, D.; Fernandez, G.; Katz, A.; Leumann, C.; Polacek, N. Oxidative stress damages rRNA inside the ribosome and differentially affects the catalytic center. Nucleic Acids Res. 2018, 46, 1945–1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genest, O.; Wickner, S.; Doyle, S.M. Hsp90 and Hsp70 chaperones: Collaborators in protein remodeling. J. Biol. Chem. 2019, 294, 2109–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamriben, L.; Graham, J.B.; Adams, B.M.; Hebert, D.N. N-Glycan-based ER molecular chaperone and protein quality control system: The calnexin binding cycle. Traffic 2016, 17, 308–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearl, L.H. Review: The HSP90 molecular chaperone-an enigmatic ATPase. Biopolymers 2016, 105, 594–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarnuczak, A.F.; Eyers, C.E.; Schwartz, J.M.; Grant, C.M.; Hubbard, S.J. Quantitative proteomics and network analysis of SSA1 and SSB1 deletion mutants reveals robustness of chaperone HSP70 network in Saccharomyces cerevisiae. Proteomics 2015, 15, 3126–3139. [Google Scholar] [CrossRef] [Green Version]
- Liao, J.; Marinelli, F.; Lee, C.; Huang, Y.; Faraldo-Gomez, J.D.; Jiang, Y. Mechanism of extracellular ion exchange and binding-site occlusion in a sodium/calcium exchanger. Nat. Struct. Mol. Biol. 2016, 23, 590–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Lee, J.; Liem, D.; Ping, P. HSPA5 Gene encoding Hsp70 chaperone BiP in the endoplasmic reticulum. Gene 2017, 618, 14–23. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein ID | Protein Name | Log2 A 1 | Log2 B 1 |
|---|---|---|---|
| Nutrient uptake | |||
| A0A2I2GMT7 | Neutral protease 2 | 1.72 | 2.10 |
| A0A2I2GLV5 | Secreted lipase | N | 2.03 |
| Q0CPJ4 | Endoglucanase 3 | N | 1.74 |
| Ergosterol synthesis pathway | |||
| A0A2I2GRQ5 | Cyp51A (ERG11) | 1.71 | N |
| A0A2I2G1P7 | C-14 sterol reductase (ERG24) | N | 2.00 |
| A0A0U5G2H3 | Putative C-8 sterol isomerase (ERG2) | 1.81 | 1.65 |
| A0A2I2GQM9 | Cytochrome P450 (ERG5) | 1.64 | 2.19 |
| Cell cycle | |||
| A0A1L9RRV9 | Cell cycle arrest protein (BUB3) | 2.02 | N |
| Q5BGV2 | DNA helicase (MCM6) | 1.61 | N |
| A0A1E3BQY7 | Serine/threonine-protein phosphatase 2A regulatory subunit A (PP2A) | 1.53 | N |
| Glycolysis | |||
| A0A0F8XU69 | Glucose-6-phosphate isomerase | 0.61 | 0.66 |
| A0A2I1C3N2 | Fructose-bisphosphate aldolase, class II | 1.57 | N |
| A0A254U7 × 1 | Triosephosphate isomerase | 1.52 | N |
| A1CD39 | Glyceraldehyde-3-phosphate dehydrogenase | 0.62 | N |
| P20445 | Glyceraldehyde-3-phosphate dehydrogenase | N | 0.65 |
| A0A1R3RQC6 | 2,3-bisphosphoglycerate-independent phosphoglycerate mutase | 0.60 | N |
| Citric acid cycle | |||
| Q0CIX6 | Malate dehydrogenase | N | 0.39 |
| A0A1F8AGJ2 | Malate dehydrogenase | N | 0.60 |
| Q2ULH0 | Aconitate hydratase, mitochondrial | N | 0.61 |
| Respiratory chain | |||
| A0A0F8WWK6 | Putative NADH-ubiquinone oxidoreductase 39 kDa subunit | N | 0.56 |
| A0A2I2FTY0 | Putative NADH-ubiquinone oxidoreductase subunit B | N | 0.60 |
| A0A1M3TZM0 | Mitochondrial succinate dehydrogenase assembly factor 2 | N | 0.65 |
| A0A2I2GQA1 | Non-heme 11 kDa protein of cytochrome bc1 complex | N | 1.71 |
| A0A2G7FY34 | Cytochrome c oxidase assembly protein (COX19) | 1.85 | 3.67 |
| A0A2I2GGG7 | COX5A-domain-containing protein | 1.68 | N |
| A0A2I2GKR3 | Cytochrome c oxidase subunit 7A | 1.58 | N |
| A0A1M3T952 | Cytochrome c oxidase subunit 5b (COX5B) | 1.57 | N |
| A0A254TW86 | COX assembly mitochondrial protein | N | 1.75 |
| A0A2I2GS70 | Cytochrome c oxidase subunit | N | 1.72 |
| H6S076 | Cytochrome c oxidase subunit 1 | N | 0.51 |
| A0A0F8V271 | ATP synthase subunit alpha | 1.71 | N |
| A0A0L1J0I5 | ATP synthase subunit alpha | 0.62 | 0.50 |
| A0A2I2FVN7 | F-type H+-transporting ATPase subunit g | 0.65 | 0.64 |
| A0A2I2G857 | ATP synthase subunit beta | N | 0.58 |
| A0A1L9UVI1 | Plasma membrane ATPase | N | 0.46 |
| Non-phosphorylated redox reaction | |||
| A0A2I2G2E3 | Cytochrome b5-like heme binding domain-containing protein | 2.08 | 1.66 |
| A0A2I2GRW8 | Putative cytochrome b5 | 1.53 | 4.10 |
| A1CEK3 | Putative cytochrome b5 | N | 2.44 |
| A0A1L9U588 | Sulfite reductase (NADPH) hemoprotein beta-component | 2.02 | N |
| A0A2I2GR30 | Assimilatory sulfite reductase | 1.61 | N |
| A0A2I2G6M7 | FAD/NAD(P)-binding domain-containing protein | 1.80 | N |
| A0A0K8LSM9 | Mitochondrial 5-demethoxyubiquinone hydroxylase | 1.61 | N |
| A0A2I2G2W0 | Thioredoxin | 1.57 | 2.48 |
| A0A2I2G575 | Thioredoxin-domain-containing protein | N | 1.92 |
| Q0CW86 | Protein disulfide-isomerase (tigA) | 1.77 | 2.72 |
| G3Y6N5 | Protein disulfide isomerase | 1.52 | 1.66 |
| Antioxidant defense system | |||
| A0A2P2H6U1 | Peroxidase | 1.54 | 1.72 |
| A0A1L9S160 | Peroxidase | N | 1.67 |
| A0A2G7FRT8 | Superoxide dismutase | 2.18 | 3.80 |
| A0A2I2FSX8 | Cu, Zn superoxide dismutase-like protein | 1.56 | 2.47 |
| A0A0F8UM10 | Catalase | 3.32 | 5.24 |
| A0A1L9RM09 | Catalase | 2.18 | 3.69 |
| A0A0F0IKG0 | Catalase | 1.87 | 2.69 |
| A0A2I2GCH5 | Catalase | 1.79 | 1.93 |
| A0A2I2FWF9 | Catalase | 1.71 | 2.63 |
| A0A1L9WU32 | Glutathione S-transferase | N | 1.66 |
| Transcription, splicing and translation | |||
| A0A2I2GRA8 | DNA-directed RNA polymerase subunit beta | N | 0.62 |
| A0A2I1D678 | Centromere/microtubule binding protein (cbf5) | 0.60 | 0.54 |
| A0A2I2GK37 | Nucleolar protein 58 | 0.63 | 0.54 |
| A0A2I2GP60 | Nucleolar GTP-binding protein 2 | N | 0.64 |
| A0A2I2GCA0 | ATP-dependent RNA helicase (dbp3) | 0.59 | 0.52 |
| A0A2I2FYP3 | Putative 40S ribosomal protein S14 | 1.58 | N |
| A0A2I2GLK4 | 37S ribosomal protein S16 | 0.66 | N |
| A0A1L9WPN8 | Small subunit ribosomal protein S18e | 0.67 | N |
| A0A1L9PM46 | Small subunit ribosomal protein S18e | 0.67 | N |
| A0A2I2FXR7 | Putative 30S ribosomal subunit S4 | N | 0.62 |
| A0A2I1CC93 | Ribosomal protein L24 | 1.61 | N |
| A0A2I2GHR7 | 60S ribosomal protein L37 | 0.41 | 0.58 |
| A0A1L9RUB2 | Large subunit ribosomal protein L28e | 0.51 | N |
| Q0CTP9 | Large subunit ribosomal protein LP1 | 0.56 | N |
| A0A1E3BC43 | 60S ribosomal protein L27a | 0.56 | 0.56 |
| A0A2I2G2I4 | 60S ribosomal protein L44 | 0.57 | N |
| Q0CRD9 | 60S ribosomal protein L20 | 0.63 | N |
| A0A1L9WKW1 | Large subunit ribosomal protein L8e | 0.63 | N |
| A0A231MJW9 | 60S ribosomal protein L13 | 0.65 | N |
| A0A2I2G636 | Ribosomal protein L22 | 0.67 | N |
| A0A2I2FUS3 | Ribosomal protein L4 | N | 0.65 |
| A0A1S9D4D7 | Seryl-tRNA synthetase | 0.65 | 0.64 |
| Q5BD96 | Valyl-tRNA synthetase | 0.60 | N |
| A0A2G7G5N0 | Valyl-tRNA synthetase | 0.66 | 0.63 |
| A0A2I1C057 | Valyl-tRNA synthetase | N | 0.66 |
| A0A0F0IBN1 | Lysyl-tRNA synthetase, class II | N | 0.58 |
| A0A0F8UQN1 | Cofactor for methionyl-and glutamyl-tRNA synthetase | N | 0.56 |
| A0A2I2GNK9 | SnRNP assembly factor | 1.73 | 1.97 |
| A1CAI8 | Putative pre-mRNA splicing factor | 1.68 | N |
| A0A1R3R8F2 | Pre-mRNA-processing factor 6 | 1.59 | N |
| A0A146FRE5 | Pre-mRNA splicing factor | 0.64 | N |
| A0A0F0IP01 | Translation initiation factor (1A/IF-1) | 1.61 | 1.96 |
| A0A146FRB4 | Translation initiation factor (SUI1) | 0.60 | N |
| A0A017S0I0 | Eukaryotic translation initiation factor 3 subunit L | 0.66 | 0.60 |
| A0A2I2GDK3 | Eukaryotic translation initiation factor 3 subunit D | 0.66 | 0.61 |
| A0A2G7FVH8 | Eukaryotic translation initiation factor 3 subunit A | N | 0.58 |
| A0A1L9RAX6 | Eukaryotic translation initiation factor 3 subunit L | N | 0.61 |
| A0A2I2GJ17 | Eukaryotic translation initiation factor 3 subunit E | N | 0.64 |
| A0A1L9UG37 | Elongation factor 1-alpha | 0.64 | 0.58 |
| A0A2I2FT46 | Elongation factor 1-beta | N | 0.62 |
| Protein folding and remodeling | |||
| A1CEK9 | Putative ER Hsp70 chaperone (BiP) | 0.67 | 0.51 |
| A0A017SPH2 | Putative Hsp70 chaperone | N | 0.63 |
| A0A2I2FVP1 | Putative Hsp70 chaperone | 0.58 | 0.52 |
| A0A017SGX5 | Heat shock protein 70 | N | 0.64 |
| A0A2I2FR82 | HSP90-domain-containing protein | 0.66 | 0.49 |
| A0A1R3RIX4 | HSP90A (HtpG) | N | 0.55 |
| Calcium signaling pathway | |||
| A0A2J5HNT7 | Outer mitochondrial membrane protein porin (VDAC2) | 1.66 | 1.87 |
| A0A1L9PAT2 | Calmodulin (CALM) | N | 1.82 |
| Autophagy | |||
| A0A2I2GJ12 | Aspartic endopeptidase (Pep2) | N | 1.66 |
| A0A229X8Y1 | Saccharopepsin (PEP4) | N | 2.43 |
| Secondary metabolism | |||
| A0A2I2GGR1 | Farnesyl pyrophosphate synthetase 1 | N | 0.63 |
| Q5D0Q7 | Nonribosomal peptide synthetase 10 | 1.7 | N |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Guan, Y.; Lin, W.; Yan, H.; Neng, J.; Sun, P. Quantitative Proteomic Profiling of Fungal Growth, Development, and Ochratoxin A Production in Aspergillus ochraceus on High- and Low-NaCl Cultures. Toxins 2021, 13, 51. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010051
Wang Y, Guan Y, Lin W, Yan H, Neng J, Sun P. Quantitative Proteomic Profiling of Fungal Growth, Development, and Ochratoxin A Production in Aspergillus ochraceus on High- and Low-NaCl Cultures. Toxins. 2021; 13(1):51. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010051
Chicago/Turabian StyleWang, Yan, Yue Guan, Wei Lin, Hao Yan, Jing Neng, and Peilong Sun. 2021. "Quantitative Proteomic Profiling of Fungal Growth, Development, and Ochratoxin A Production in Aspergillus ochraceus on High- and Low-NaCl Cultures" Toxins 13, no. 1: 51. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13010051