Pan-American Lancehead Pit-Vipers: Coagulotoxic Venom Effects and Antivenom Neutralisation of Bothrops asper and B. atrox Geographical Variants

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Snake Venom Sample Preparation
4.2. Human Plasma
4.3. Venom-Induced Coagulation
4.4. Antivenom Neutralising Efficacy
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; de Silva, H.J. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruha, A.-M.; Kleinschmidt, K.C.; Greene, S.; Spyres, M.B.; Brent, J.; Wax, P.; Padilla-Jones, A.; Campleman, S. The epidemiology, clinical course, and management of snakebites in the North American snakebite registry. J. Med. Toxicol. 2017, 13, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Dolab, J.A.; de Roodt, A.R.; de Titto, E.H.; García, S.I.; Funes, R.; Salomón, O.D.; Chippaux, J.-P. Epidemiology of snakebite and use of antivenom in Argentina. Trans. R. Soc. Trop. Med. Hyg. 2014, 108, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, M.K.B.; da Fonseca, C.S.; Navoni, J.A.; Freire, E.M.X. Snakebite accidents in Rio Grande do Norte state, Brazil: Epidemiology, health management and influence of the environmental scenario. Trop. Med. Int. Health 2019, 24, 432–441. [Google Scholar] [CrossRef]
- Chippaux, J.-P. Epidemiology of envenomations by terrestrial venomous animals in Brazil based on case reporting: From obvious facts to contingencies. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Mota-da-Silva, A.; Colombini, M.; Moura-da-Silva, A.M.; Souza, R.M.; Monteiro, W.M.; Bernarde, P.S. Epidemiological and clinical aspects of snakebites in the upper Juruá River region, western Brazilian Amazonia. Acta Amaz. 2019. [Google Scholar] [CrossRef]
- Monteiro, W.M.; Contreras-Bernal, J.C.; Bisneto, P.F.; Sachett, J.; Mendonça da Silva, I.; Lacerda, M.; Guimarães da Costa, A.; Val, F.; Brasileiro, L.; Sartim, M.A.; et al. Bothrops atrox, the most important snake involved in human envenomings in the amazon: How venomics contributes to the knowledge of snake biology and clinical toxinology. Toxicon X 2020, 6, 100037. [Google Scholar] [CrossRef]
- Otero-Patiño, R. Epidemiological, clinical and therapeutic aspects of Bothrops asper bites. Toxicon 2009, 54, 998–1011. [Google Scholar] [CrossRef]
- Gutiérrez, J.M. Current challenges for confronting the public health problem of snakebite envenoming in Central America. J. Venom. Anim. Toxins Incl. Trop. Dis. 2014, 20, 7. [Google Scholar] [CrossRef] [Green Version]
- Chippaux, J.-P. Incidence and mortality due to snakebite in the Americas. PLoS Negl. Trop. Dis. 2017, 11, e0005662. [Google Scholar] [CrossRef]
- Sousa, L.F.; Zdenek, C.N.; Dobson, J.S.; Op den Brouw, B.; Coimbra, F.C.P.; Gillett, A.; Del-Rei, T.H.M.; Chalkidis, H.D.M.; Sant’Anna, S.; Teixeira-da-Rocha, M.M.; et al. Coagulotoxicity of Bothrops (lancehead pit-vipers) venoms from Brazil: Differential biochemistry and antivenom efficacy resulting from prey-driven venom variation. Toxins 2018, 10, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colman, R.W.; Clowes, A.W.; George, J.N.; Goldhaber, S.Z.; Marder, V.J. Overview of hemostasis. In Hemostasis and Thrombosis: Basic Principles and Clinical Practice; Colman, R.W., Clowes, A.W., George, J.N., Goldhaber, S.Z., Marder, V.J., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2006; pp. 3–20. [Google Scholar]
- Kini, R.M. The intriguing world of prothrombin activators from snake venom. Toxicon 2005, 45, 1133–1145. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M. Anticoagulant proteins from snake venoms: Structure, function and mechanism. Biochem. J. 2006, 397, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kini, R.M.; Rao, V.S.; Joseph, J.S. Procoagulant proteins from snake venoms. Pathophysiol. Haemost. Thromb. 2001, 31, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Markland, F.S. Snake venoms and the hemostatic system. Toxicon 1998, 36, 1749–1800. [Google Scholar] [CrossRef]
- Loría, G.D.; Rucavado, A.; Kamiguti, A.S.; Theakston, R.D.G.; Fox, J.W.; Alape, A.; Gutiérrez, J.M.A. Characterization of ‘basparin A,’ a prothrombin-activating metalloproteinase, from the venom of the snake Bothrops asper that inhibits platelet aggregation and induces defibrination and thrombosis. Arch. Biochem. Biophys. 2003, 418, 13–24. [Google Scholar] [CrossRef]
- Rucavado, A.; Escalante, T.; Gutiérrez, J.M.a. Effect of the metalloproteinase inhibitor batimastat in the systemic toxicity induced by Bothrops asper snake venom: Understanding the role of metalloproteinases in envenomation. Toxicon 2004, 43, 417–424. [Google Scholar] [CrossRef]
- Sousa, L.F.; Bernardoni, J.L.; Zdenek, C.N.; Dobson, J.; Coimbra, F.; Gillett, A.; Lopes-Ferreira, M.; Moura-da-Silva, A.M.; Fry, B.G. Differential coagulotoxicity of metalloprotease isoforms from Bothrops neuwiedi snake venom and consequent variations in antivenom efficacy. Toxicol. Lett. 2020, 333, 211–221. [Google Scholar] [CrossRef]
- Fuchs, J.; Faber, K.; Tuchscherer, D.T.; Tsakiris, D.A.; Weiler, S.; Hofer, K.E. Bite by a juvenile Bothrops venezuelensis (Venezuelan lancehead) resulting in severe envenomation: A case report. Toxicon 2020, 180, 39–42. [Google Scholar] [CrossRef]
- Resiere, D.; Mehdaoui, H.; Gutiérrez, J.M. Snakebite envenomation in the Caribbean: The role of medical and scientific cooperation. PLoS Negl. Trop. Dis. 2018, 12, e0006441. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Lomonte, B.; Sanz, L.; Calvete, J.J.; Pla, D. Immunological profile of antivenoms: Preclinical analysis of the efficacy of a polyspecific antivenom through antivenomics and neutralization assays. J. Proteom. 2014, 105, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Nadaud, A.; Perotti, F.; de Haro, L.; Boels, D. Snake envenomations in French Guiana: First clinical assessment of an antivenom imported from Mexico. Anaesth. Crit. Care Pain Med. 2019, 38, 193–194. [Google Scholar] [CrossRef] [PubMed]
- Heckmann, X.; Lambert, V.; Mion, G.; Ehrhardt, A.; Marty, C.; Perotti, F.; Carod, J.-F.; Jolivet, A.; Boels, D.; Lehida Andi, I.; et al. Failure of a Mexican antivenom on recovery from snakebite-related coagulopathy in French Guiana. Clin. Toxicol. 2020, 1–7. [Google Scholar] [CrossRef]
- De Silva Oliveira, S.; Campos Alves, E.; dos Santos Santos, A.; Freitas Nascimento, E.; Tavares Pereira, J.P.; da Mendonça Silva, I.; Sachett, J.; dos Santos Ibiapina, H.N.; Santos Sarraf, L.K.; Contreras Bernal, J.C.; et al. Bothrops snakebites in the Amazon: Recovery from hemostatic disorders after Brazilian antivenom therapy. Clin. Toxicol. 2020, 58, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Barlow, A.; Pook, C.E.; Harrison, R.A.; Wüster, W. Coevolution of diet and prey-specific venom activity supports the role of selection in snake venom evolution. Proc. R. Soc. Biol. Sci. Ser. B 2009, 276, 2443–2449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daltry, J.C.; Wüster, W.; Thorpe, R.S. Diet and snake venom evolution. Nature 1996, 379, 537–540. [Google Scholar] [CrossRef]
- Jackson, T.N.W.; Koludarov, I.; Ali, S.A.; Dobson, J.; Zdenek, C.N.; Dashevsky, D.; Op den Brouw, B.; Masci, P.P.; Nouwens, A.; Josh, P.; et al. Rapid radiations and the race to redundancy: An investigation of the evolution of Australian elapid snake venoms. Toxins 2016, 8, 309. [Google Scholar] [CrossRef]
- Sasa, M. Diet and snake venom evolution: Can local selection alone explain intraspecific venom variation? Toxicon 1999, 37, 253–260. [Google Scholar] [CrossRef]
- Williams, V.; White, J.; Schwaner, T.D.; Sparrow, A. Variation in venom proteins from isolated populations of tiger snakes (Notechis ater niger, N. scutatus) in South Australia. Toxicon 1988, 26, 1067–1075. [Google Scholar] [CrossRef]
- Calvete, J.J.; Sanz, L.; Pérez, A.; Borges, A.; Vargas, A.M.; Lomonte, B.; Angulo, Y.; Gutiérrez, J.M.; Chalkidis, H.M.; Mourão, R.H.V.; et al. Snake population venomics and antivenomics of Bothrops atrox: Paedomorphism along its transamazonian dispersal and implications of geographic venom variability on snakebite management. J. Proteom. 2011, 74, 510–527. [Google Scholar] [CrossRef]
- Alape-Girón, A.; Sanz, L.; Escolano, J.; Flores-Díaz, M.; Madrigal, M.; Sasa, M.; Calvete, J.J. Snake venomics of the lancehead pitviper Bothrops asper: Geographic, individual, and ontogenetic variations. J. Proteome Res. 2008, 7, 3556–3571. [Google Scholar] [CrossRef] [PubMed]
- Aragón, F.; Gubenšek, F. Bothrops asper venom from the Atlantic and Pacific zones of Costa Rica. Toxicon 1981, 19, 797–805. [Google Scholar] [CrossRef]
- Wüster, W.; Salomão, M.; Quijada-Mascareñas, J.A.; Thorpe, R.; BBBSP. Origins and evolution of the South American pitviper fauna: Evidence from mitochondrial DNA sequence analysis. In Biology of the Vipers; NHBS Academic & Professional Books: Eagle Mountain, UT, USA, 2002; pp. 111–128. [Google Scholar]
- Saldarriaga-Córdoba, M.; Parkinson, C.L.; Daza, J.M.; Wüster, W.; Sasa, M. Phylogeography of the Central American lancehead Bothrops asper (Serpentes: Viperidae). PLoS ONE 2017, 12, e0187969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora-Obando, D.; Salazar-Valenzuela, D.; Pla, D.; Lomonte, B.; Guerrero-Vargas, J.A.; Ayerbe, S.; Gibbs, H.L.; Calvete, J.J. Venom variation in Bothrops asper lineages from North-Western South America. J. Proteom. 2020, 229, 103945. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, C.C.; Argôlo, A.J.S.; Arzamendia, V.; Azevedo, J.A.; Barbo, F.E.; Bérnils, R.S.; Bolochio, B.E.; Borges-Martins, M.; Brasil-Godinho, M.; Braz, H.; et al. Atlas of Brazilian Snakes: Verified point-locality maps to mitigate the Wallacean shortfall in a megadiverse snake fauna. S. Am. J. Herpetol. 2019, 14, 1–274. [Google Scholar] [CrossRef]
- Campbell, J.A.; Lamar, W.W. The Venomous Reptiles of the Western Hemisphere; Comstock Publishing Associates: Ithaca, NY, USA, 2004; Volume 1. [Google Scholar]
- Gibbs, H.L.; Sovic, M.; Amazonas, D.; Chalkidis, H.; Salazar-Valenzuela, D.; Moura-Da-Silva, A.M. Recent lineage diversification in a venomous snake through dispersal across the Amazon River. Biol. J. Linn. Soc. 2018, 123, 651–665. [Google Scholar] [CrossRef] [Green Version]
- Wüster, W.; Salomão, M.G.; Duckett, G.J.; Thorpe, R.S. BBBSP Mitochondrial DNA phylogeny of the Bothrops atrox species complex (Squamata: Serpentes: Viperidae). Kaupia Darmst Beitr Natgesch 1999, 8, 135–144. [Google Scholar]
- Salazar Valenzuela, C.D. Diversification in the Neotropics: Insights from Demographic and Phylogenetic Patterns of Lancehead Pitvipers (Bothrops spp.). Ph.D. Thesis, The Ohio State University, Columbus, OH, USA, 2016. [Google Scholar]
- Resiere, D.; Villalta, M.; Arias, A.S.; Kallel, H.; Nèviére, R.; Vidal, N.; Mehdaoui, H.; Gutiérrez, J.M. Snakebite envenoming in French Guiana: Assessment of the preclinical efficacy against the venom of Bothrops atrox of two polyspecific antivenoms. Toxicon 2020, 173, 1–4. [Google Scholar] [CrossRef]
- Resiere, D.; Houcke, S.; Pujo, J.M.; Mayence, C.; Mathien, C.; NkontCho, F.; Blaise, N.; Demar, M.P.; Hommel, D.; Kallel, H. Clinical features and management of snakebite envenoming in French Guiana. Toxins 2020, 12, 662. [Google Scholar] [CrossRef]
- Segura, A.; Castillo, M.C.; Núñez, V.; Yarlequé, A.; Gonçalves, L.R.C.; Villalta, M.; Bonilla, C.; Herrera, M.; Vargas, M.; Fernández, M.; et al. Preclinical assessment of the neutralizing capacity of antivenoms produced in six Latin American countries against medically-relevant Bothrops snake venoms. Toxicon 2010, 56, 980–989. [Google Scholar] [CrossRef]
- Thomas, L.; Chausson, N.; Uzan, J.; Kaidomar, S.; Vignes, R.; Plumelle, Y.; Bucher, B.; Smadja, D. Thrombotic stroke following snake bites by the “Fer-de-Lance” Bothrops lanceolatus in Martinique despite antivenom treatment: A report of three recent cases. Toxicon 2006, 48, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Otero, R.; Gutiérrez, J.; Beatriz Mesa, M.a.; Duque, E.; Rodríguez, O.; Luis Arango, J.; Gómez, F.; Toro, A.; Cano, F.; María Rodríguez, L.; et al. Complications of Bothrops, Porthidium, and Bothriechis snakebites in Colombia. A clinical and epidemiological study of 39 cases attended in a university hospital. Toxicon 2002, 40, 1107–1114. [Google Scholar] [CrossRef]
- Isbister, G.K. Snakebite doesn’t cause disseminated intravascular coagulation: Coagulopathy and thrombotic microangiopathy in snake envenoming. Semin. Thromb. Hemost. 2010, 36, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Resiere, D.; Mégarbane, B.; Valentino, R.; Mehdaoui, H.; Thomas, L. Bothrops lanceolatus bites: Guidelines for severity assessment and emergent management. Toxins 2010, 2, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Currier, R.B.; Harrison, R.A.; Rowley, P.D.; Laing, G.D.; Wagstaff, S.C. Intra-specific variation in venom of the African Puff Adder (Bitis arietans): Differential expression and activity of snake venom metalloproteinases (SVMPs). Toxicon 2010, 55, 864–873. [Google Scholar] [CrossRef]
- Oliveira, I.S.d.; Cardoso, I.A.; Bordon, K.d.C.F.; Carone, S.E.I.; Boldrini-França, J.; Pucca, M.B.; Zoccal, K.F.; Faccioli, L.H.; Sampaio, S.V.; Rosa, J.C.; et al. Global proteomic and functional analysis of Crotalus durissus collilineatus individual venom variation and its impact on envenoming. J. Proteom. 2019, 191, 153–165. [Google Scholar] [CrossRef]
- Tasoulis, T.; Silva, A.; Veerati, P.C.; Baker, M.; Hodgson, W.C.; Dunstan, N.; Isbister, G.K. Intra-specific venom variation in the Australian coastal taipan Oxyuranus scutellatus. Toxins 2020, 12, 485. [Google Scholar] [CrossRef]
- Estevao-Costa, M.I.; Gontijo, S.S.; Correia, B.L.; Yarleque, A.; Vivas-Ruiz, D.; Rodrigues, E.; Chávez-Olortegui, C.; Oliveira, L.S.; Sanchez, E.F. Neutralization of toxicological activities of medically-relevant Bothrops snake venoms and relevant toxins by two polyvalent bothropic antivenoms produced in Peru and Brazil. Toxicon 2016, 122, 67–77. [Google Scholar] [CrossRef]
- Kohlhoff, M.; Borges, M.H.; Yarleque, A.; Cabezas, C.; Richardson, M.; Sanchez, E.F. Exploring the proteomes of the venoms of the Peruvian pit vipers Bothrops atrox, B. barnetti and B. pictus. J. Proteom. 2012, 75, 2181–2195. [Google Scholar] [CrossRef]
- Bogarín, G.; Romero, M.; Rojas, G.; Lutsch, C.; Casadamont, M.; Lang, J.; Otero, R.; Gutiérrez, J.M.a. Neutralization, by a monospecific Bothrops lanceolatus antivenom, of toxic activities induced by homologous and heterologous Bothírops snake venoms. Toxicon 1999, 37, 551–557. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Sanz, L.; Escolano, J.; Fernández, J.; Lomonte, B.; Angulo, Y.; Rucavado, A.; Warrell, D.A.; Calvete, J.J. Snake venomics of the Lesser Antillean pit vipers Bothrops caribbaeus and Bothrops lanceolatus: Correlation with toxicological activities and immunoreactivity of a heterologous antivenom. J. Proteome Res. 2008, 7, 4396–4408. [Google Scholar] [CrossRef] [PubMed]
- Niewiarowski, S.; Kirby, E.P.; Brudzynski, T.M.; Stocker, K. Thrombocytin, a serine protease from Bothrops atrox venom. 2. Interaction with platelets and plasma-clotting factors. Biochemistry 1979, 18, 3570–3577. [Google Scholar] [CrossRef] [PubMed]
- Rucavado, A.; Soto, M.; Escalante, T.; Loría, G.D.; Arni, R.; Gutiérrez, J.M. Thrombocytopenia and platelet hypoaggregation induced by Bothrops asper snake venom. Thromb. Haemost. 2005, 94, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Rucavado, A.; Soto, M.; Kamiguti, A.S.; Theakston, D.G.; Fox, J.W.; Escalante, T.; Gutiérrez, J.M. Characterization of aspercetin, a platelet aggregating component from the venom of the snake Bothrops asper which induces thrombocytopenia and potentiates metalloproteinase-induced hemorrhage. Thromb. Haemost. 2001, 85, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.; Ownby, C.L.; Odell, G.V. Isolation of a myotoxin from Bothrops asper venom: Partial characterization and action on skeletal muscle. Toxicon 1984, 22, 115–128. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Lomonte, B. Phospholipase A2 myotoxins from Bothrops snake venoms. Toxicon 1995, 33, 1405–1424. [Google Scholar] [CrossRef]
- Lister, C.; Arbuckle, K.; Jackson, T.N.W.; Debono, J.; Zdenek, C.N.; Dashevsky, D.; Dunstan, N.; Allen, L.; Hay, C.; Bush, B.; et al. Catch a tiger snake by its tail: Differential toxicity, co-factor dependence and antivenom efficacy in a procoagulant clade of Australian venomous snakes. Comp. Biochem. Physiol. C Toxicol. Pharm. 2017, 202, 39–54. [Google Scholar] [CrossRef] [Green Version]
- O’Leary, M.A.; Kornhauser, R.S.; Hodgson, W.C.; Isbister, G.K. An examination of the activity of expired and mistreated commercial Australian antivenoms. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 937–942. [Google Scholar] [CrossRef]
- Al-Abdulla, I.; Casewell, N.R.; Landon, J. Long-term physicochemical and immunological stability of a liquid formulated intact ovine immunoglobulin-based antivenom. Toxicon 2013, 64, 38–42. [Google Scholar] [CrossRef]
- Da Galizio, N.C.; Serino-Silva, C.; Stuginski, D.R.; Abreu, P.A.E.; Sant’Anna, S.S.; Grego, K.F.; Tashima, A.K.; Tanaka-Azevedo, A.M.; de Morais-Zani, K. Compositional and functional investigation of individual and pooled venoms from long-term captive and recently wild-caught Bothrops jararaca snakes. J. Proteom. 2018, 186, 56–70. [Google Scholar] [CrossRef]
- Castellone, D. How to deliver quality results in the coagulation laboratory: Commonly asked questions. Lab. Med. 2004, 35, 208–213. [Google Scholar] [CrossRef]
- Bourke, L.A.; Youngman, N.J.; Zdenek, C.N.; op den Brouw, B.; Violette, A.; Fourmy, R.; Fry, B.G. Trimeresurus albolabris snakebite treatment implications arising from ontogenetic venom comparisons of anticoagulant function, and antivenom efficacy. Toxicol. Lett. 2020, 327, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Debono, J.; Bos, M.H.A.; Frank, N.; Fry, B. Clinical implications of differential antivenom efficacy in neutralising coagulotoxicity produced by venoms from species within the arboreal viperid snake genus Trimeresurus. Toxicol. Lett. 2019, 316, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Zdenek, C.N.; Hay, C.; Arbuckle, K.; Jackson, T.N.W.; Bos, M.H.A.; op den Brouw, B.; Debono, J.; Allen, L.; Dunstan, N.; Morley, T.; et al. Coagulotoxic effects by brown snake (Pseudonaja) and taipan (Oxyuranus) venoms, and the efficacy of a new antivenom. Toxicol. In Vitro 2019, 58, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, G.; Jiménez, J.; Gutiérrez, J. Caprylic acid fractionation of hyperimmune horse plasma: Description of a simple procedure for antivenom production. Toxicon 1994, 32, 351–363. [Google Scholar] [CrossRef]




{kind=link}
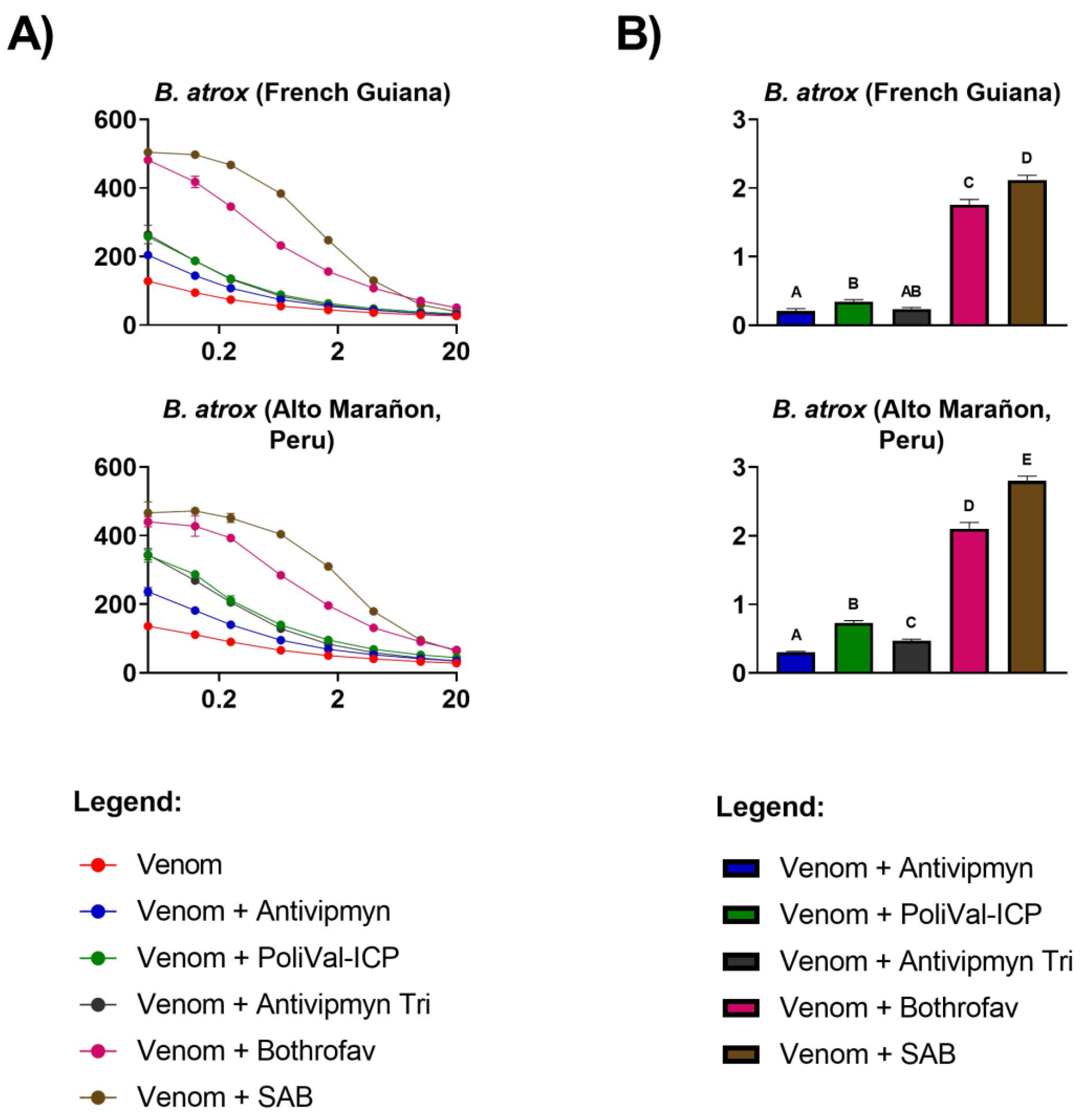{kind=link}
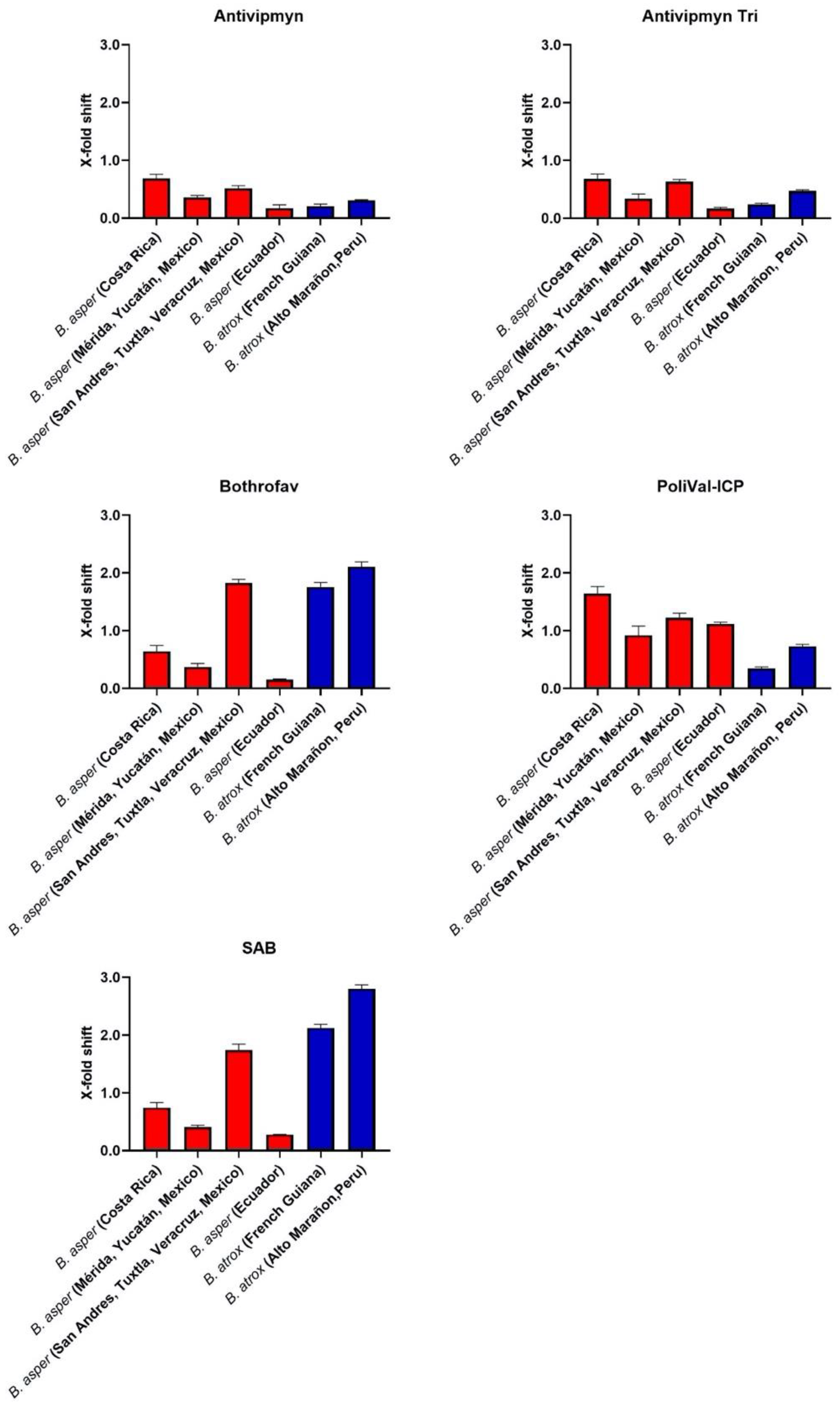{kind=link}
| Venom | 20 μg/mL Clotting Time | AUC Value |
|---|---|---|
| B. asper (Costa Rica) | 17.73 ± 2.17 | 481.70 ± 18.01 |
| B. asper (Ecuador) | 13.50 ± 0.85 | 384.00 ± 5.86 |
| B. asper (Mérida, Yucatán, Mexico) | 14.27 ± 1.12 | 377.33 ± 9.15 |
| B. asper (San Andres, Tuxtla, Veracruz, Mexico) | 47.20 ± 2.04 | 1327.33 ± 26.03 |
| B. atrox (French Guiana) | 26.80 ± 0.96 | 669.70 ± 15.85 |
| B. atrox (Alto Marañon, Peru) | 28.30 ± 0.66 | 741.73 ± 5.47 |
| B. asper (Costa Rica) | B. asper (Ecuador) | B. asper (Mérida, Yucatán, Mexico) | B. asper (San Andres, Tuxtla, Veracruz, Mexico) | B. atrox (French Guiana) | B. atrox (Alto Marañon, Peru) | |
|---|---|---|---|---|---|---|
| B. asper (Costa Rica) | X | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| B. asper (Ecuador) | X | X | 0.9936 | <0.0001 | <0.0001 | <0.0001 |
| B. asper (Mérida, Yucatán, Mexico) | X | X | X | <0.0001 | <0.0001 | <0.0001 |
| B. asper (San Andres, Tuxtla, Veracruz, Mexico) | X | X | X | X | <0.0001 | <0.0001 |
| B. atrox (French Guiana) | X | X | X | X | X | 0.0010 |
| B. atrox (Alto Marañon, Peru) | X | X | X | X | X | X |
| B. asper (Costa Rica) | B. asper (Ecuador) | B. asper (Mérida, Yucatán, Mexico) | B. asper (San Andres, Tuxtla, Veracruz, Mexico) | |
|---|---|---|---|---|
| Antivipmyn vs. PoliVal-ICP | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Antivipmyn vs. Antivipmyn-Tri | >0.9999 | 0.9996 | 0.9984 | 0.3096 |
| Antivipmyn vs. Bothrofav | 0.9710 | 0.9667 | 0.9993 | <0.0001 |
| Antivipmyn vs. SAB | 0.9437 | 0.0169 | 0.9525 | <0.0001 |
| PoliVal-ICP vs. Antivipmyn-Tri | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| PoliVal-ICP vs. Bothrofav | <0.0001 | <0.0001 | 0.0001 | <0.0001 |
| PoliVal-ICP vs. SAB | <0.0001 | <0.0001 | 0.0002 | <0.0001 |
| Antivipmyn-Tri vs. Bothrofav | 0.9848 | 0.9916 | 0.9851 | <0.0001 |
| Antivipmyn-Tri vs. SAB | 0.9146 | 0.0125 | 0.8598 | <0.0001 |
| Bothrofav vs. SAB | 0.6759 | 0.0066 | 0.9882 | 0.5859 |
| B. atrox (French Guiana) | B. atrox (Alto Marañon, Peru) | |
|---|---|---|
| Antivipmyn vs. PoliVal-ICP | 0.0408 | <0.0001 |
| Antivipmyn vs. Antivipmyn-Tri | 0.9318 | 0.0266 |
| Antivipmyn vs. Bothrofav | <0.0001 | <0.0001 |
| Antivipmyn vs. SAB | <0.0001 | <0.0001 |
| PoliVal-ICP vs. Antivipmyn-Tri | 0.1344 | 0.0013 |
| PoliVal-ICP vs. Bothrofav | <0.0001 | <0.0001 |
| PoliVal-ICP vs. SAB | <0.0001 | <0.0001 |
| Antivipmyn-Tri vs. Bothrofav | <0.0001 | <0.0001 |
| Antivipmyn-Tri vs. SAB | <0.0001 | <0.0001 |
| Bothrofav vs. SAB | <0.0001 | <0.0001 |
| Species | Antivenom | B. asper (Costa Rica) | B. asper (Mérida, Yucatán, Mexico) | B. asper (San Andres, Tuxtla, Veracruz, Mexico) | B. asper (Ecuador) |
|---|---|---|---|---|---|
| B. atrox (French Guiana) | PoliVal-ICP | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| SAB | <0.0001 | <0.0001 | 0.0002 | <0.0001 | |
| Bothrofav | <0.0001 | <0.0001 | 0.8651 | <0.0001 | |
| B. atrox (Alto Marañon, Peru) | PoliVal-ICP | <0.0001 | 0.1637 | 0.0003 | 0.0023 |
| SAB | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| Bothrofav | <0.0001 | <0.0001 | 0.0065 | <0.0001 |
| Central America and Mexico | ||||
|---|---|---|---|---|
| B. asper (Costa Rica) | B. asper (Mérida, Yucatán, Mexico) | B. asper (San Andres, Tuxtla, Veracruz, Mexico) | ||
| South America | B. asper (Ecuador) | <0.0001 | 0.0044 | <0.0001 |
| B. atrox (French Guiana) | <0.0001 | 0.0213 | <0.0001 | |
| B. atrox (Alto Marañon, Peru) | <0.0001 | 0.7904 | 0.0019 | |
| Species | Locality | Abbreviation | Additional Information |
|---|---|---|---|
| Bothrops asper | Costa Rica | N.A. | Pooled from adults collected in the Pacific region (n = 40) |
| Bothrops asper | Ecuador | N.A. | Pooled from captive-born adult males (n = 2) |
| Bothrops asper | Mérida, Yucatán, Mexico | Yucatán, Mexico | Adult (individual) |
| Bothrops asper | San Andres, Tuxtla, Veracruz, Mexico | S.A.T, Mexico | Adult (individual) |
| Bothrops atrox | French Guiana | N.A. | Pooled from captive-born and wild adults (male + females, n = 66) |
| Bothrops atrox | Alto Marañon, Peru | A.M., Peru | Pooled from wild adults caught in the Amazon rainforest and kept in captivity (n values not supplied) |
| Antivenom | Lot # and Expiry | Immunising Mixture * | Preparation | Neutralising Potency (Specified by Manufacturer) |
|---|---|---|---|---|
| Antivipmyn®, Instituto Bioclon, Mexico | Lot: B-6F-16; Exp: October 2010 | Bothrops asper (Mexico), Crotalus simus simus (Mexico) | Polyspecific equine F(ab’)2 preparation | 1 vial neutralises >780 LD50 of Bothrops sp. and >790 of Crotalus sp. |
| Antivipmyn-Tri®, Instituto Bioclon, Mexico | Lot: B-4F-13; Exp: 28 June 2009 | Bothrops asper (Colombia), Crotalus simus simus (Mexico), and Lachesis muta (Not stated) | Polyspecific equine F(ab’)2 preparation | Unknown |
| Soro Antibotrópico (SAB)®, Instituto Butantan, Brazil | Lot: 1305077; Exp: May 2016 | Bothrops jararaca (Brazil) (50%), Bothrops jararacussu (Brazil) (12.5%), Bothrops neuwiedi (Brazil) (12.5%), Bothrops alternatus (Brazil) (12.5%), Bothrops moojeni (Brazil) (12.5%) | Polyspecific equine IgG F(ab′)2 preparation | 1 mL neutralises 5 mg venom of Botrops jararaca (Brazil) |
| Central American polyspecific antivenom (PoliVal-ICP)®, Instituto Clodomiro Picado, Universidad de Costa Rica | Lot: 5720416; Exp: April 2021 | Bothrops asper (Costa Rica), Crotalus simus (Costa Rica), and Lachesis stenophrys (Costa Rica) [69] | Polyspecific equine whole IgG—purified by caprylic acid fractionation [69] | 1ml neutralises 3 mg venom of Bothrops asper (Costa Rica) |
| Bothrofav®, MicroPharm, United Kingdom | Lot P4AP61V; Exp: October 2020 | Bothrops lanceolatus | Monospecific equine F(ab′)2 preparation | Unknown |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bourke, L.A.; Zdenek, C.N.; Neri-Castro, E.; Bénard-Valle, M.; Alagón, A.; Gutiérrez, J.M.; Sanchez, E.F.; Aldridge, M.; Fry, B.G. Pan-American Lancehead Pit-Vipers: Coagulotoxic Venom Effects and Antivenom Neutralisation of Bothrops asper and B. atrox Geographical Variants. Toxins 2021, 13, 78. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020078
Bourke LA, Zdenek CN, Neri-Castro E, Bénard-Valle M, Alagón A, Gutiérrez JM, Sanchez EF, Aldridge M, Fry BG. Pan-American Lancehead Pit-Vipers: Coagulotoxic Venom Effects and Antivenom Neutralisation of Bothrops asper and B. atrox Geographical Variants. Toxins. 2021; 13(2):78. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020078
Chicago/Turabian StyleBourke, Lachlan A., Christina N. Zdenek, Edgar Neri-Castro, Melisa Bénard-Valle, Alejandro Alagón, José María Gutiérrez, Eladio F. Sanchez, Matt Aldridge, and Bryan G. Fry. 2021. "Pan-American Lancehead Pit-Vipers: Coagulotoxic Venom Effects and Antivenom Neutralisation of Bothrops asper and B. atrox Geographical Variants" Toxins 13, no. 2: 78. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020078