First Evidence of the Toxin Domoic Acid in Antarctic Diatom Species

 , , ,
, , ,
Abstract
:1. Introduction
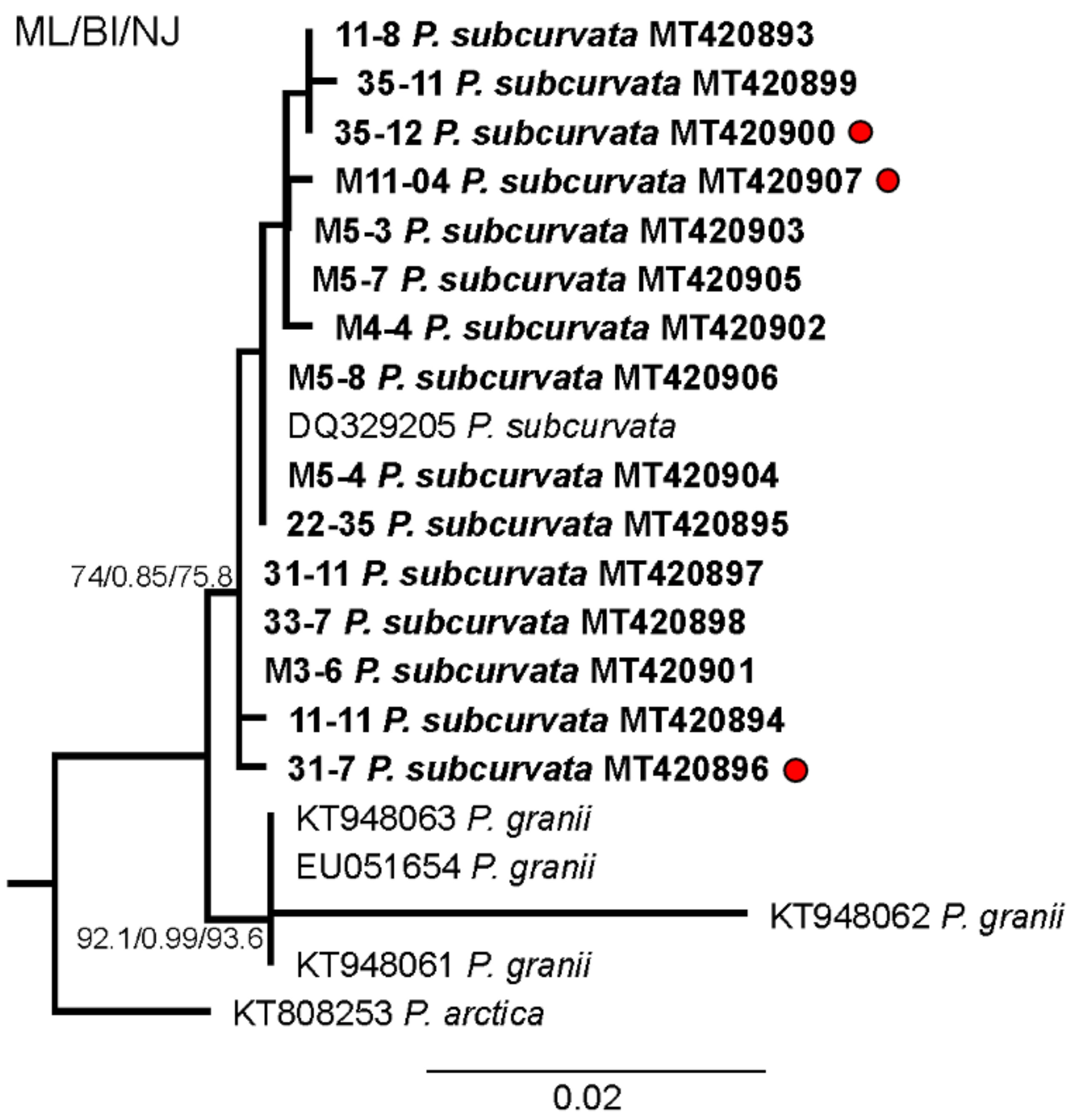
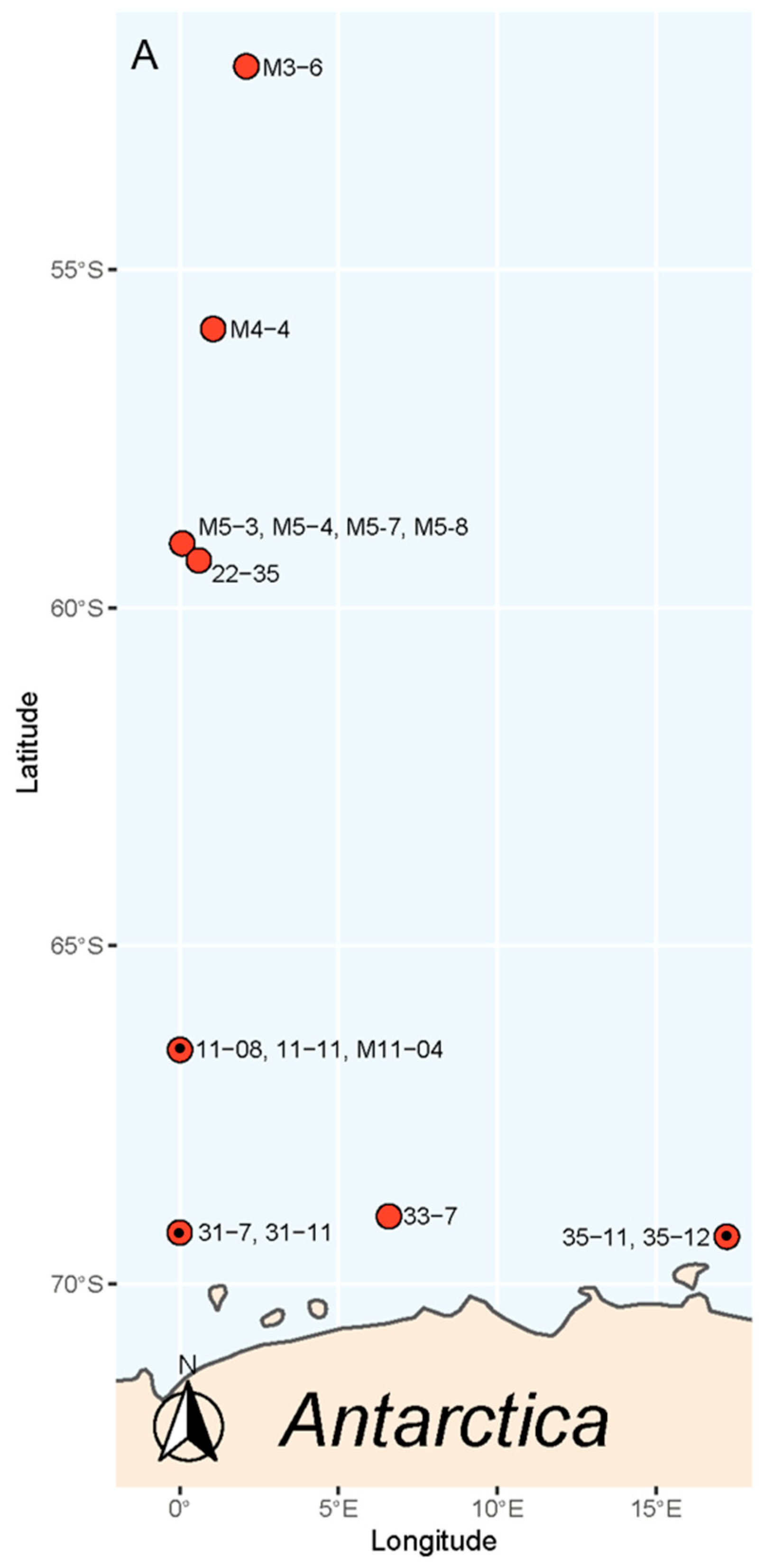
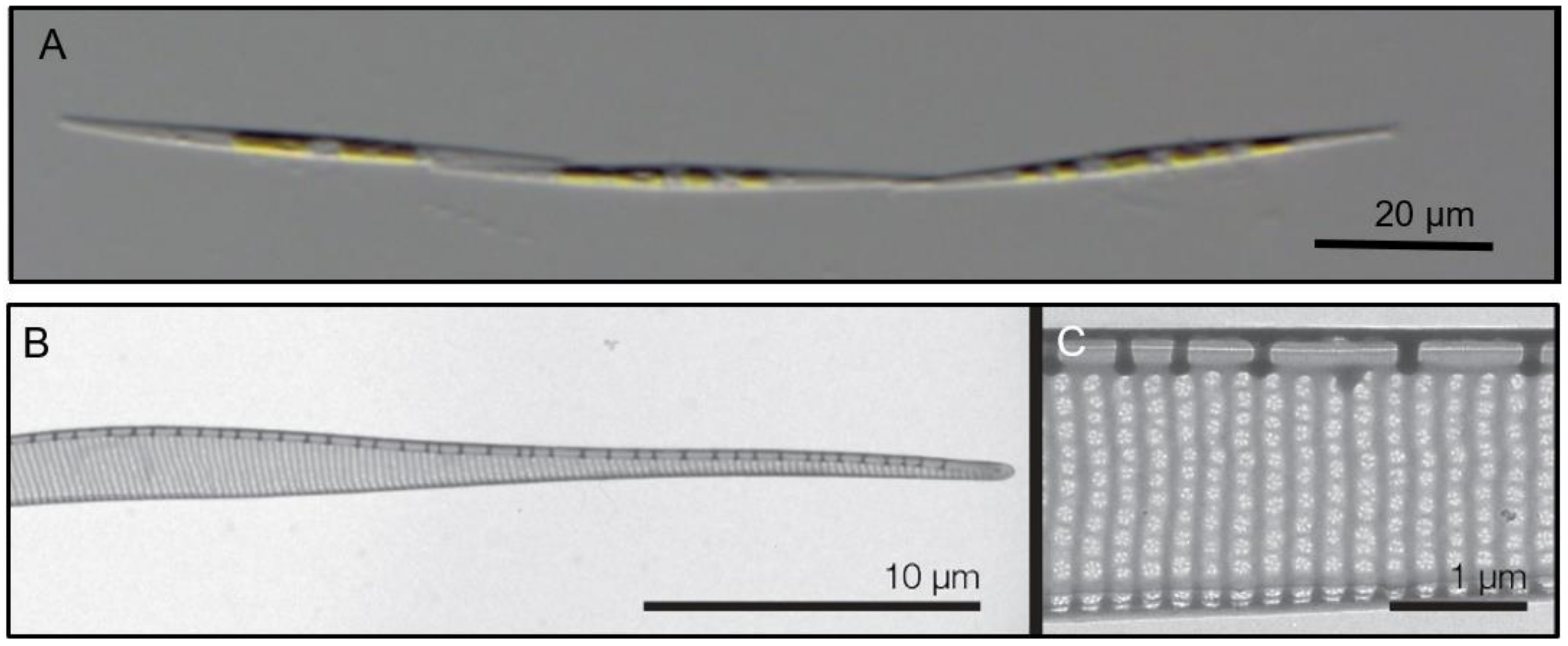
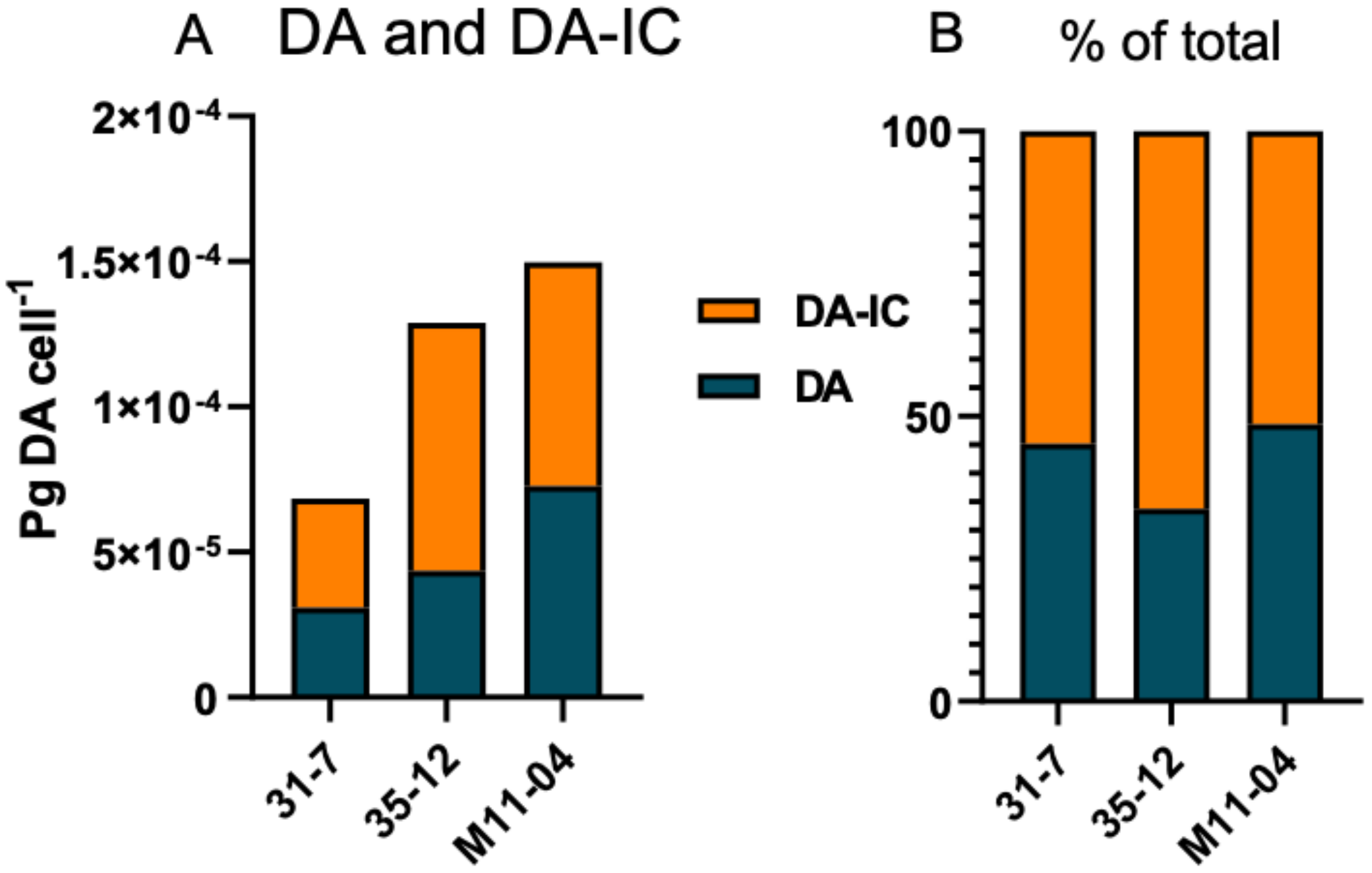
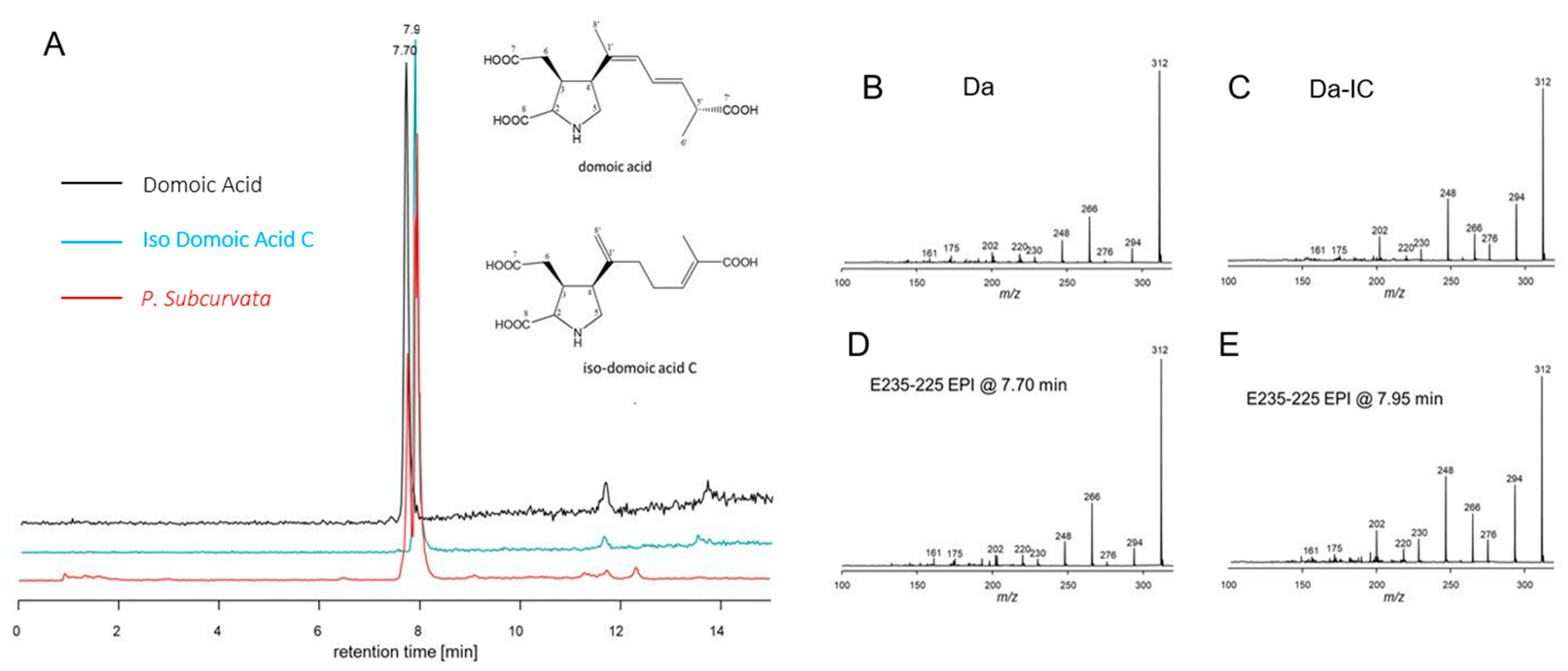
2. Results
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bates, S.S.; Hubbard, K.A.; Lundholm, N.; Montresor, M.; Leaw, C.P. Pseudo-nitzschia, Nitzschia, and domoic acid: New research since 2011. Harmful Algae 2018, 79, 3–43. [Google Scholar] [CrossRef]
- Almandoz, G.O.; Ferreyra, G.A.; Schloss, I.R.; Dogliotti, A.I.; Rupolo, V.; Paparazzo, F.E.; Esteves, J.L.; Ferrario, M.E. Distribution and ecology of Pseudo-nitzschia species (Bacillariophyceae) in surface waters of the Weddell Sea (Antarctica). Polar Biol. 2008, 31, 429–442. [Google Scholar] [CrossRef]
- Malviya, S.; Malviya, S.; Scalco, E.; Audic, S.; Vincent, F.; Veluchamy, A.; Poulain, J.; Wincker, P.; Iudicone, D.; de Vargas, C.; et al. Insights into global diatom distribution and diversity in the world’s ocean. Proc. Natl. Acad. Sci. USA 2016, 11, E1516–E1525. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, K.A.; Quakenbush, L.; Frame, E.; Huntington, K.B.; Sheffield, G.; Stimmelmayr, R.; Bryan, A.; Kendrick, P.; Ziel, H.; Goldstein, T.; et al. Prevalence of algal toxins in Alaskan marine mammals foraging in a changing arctic and subarctic environment. Harmful Algae 2016, 55, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, K.A.; Quakenbush, L.; Frame, E.; Huntington, K.B.; Sheffield, G.; Stimmelmayr, R.; Bryan, A.; Kendrick, P.; Ziel, H.; Goldstein, T.; et al. Epidemiology of Domoic Acid Poisoning in Brown Pelicans (Pelecanus occidentalis) and Brandt’ s Cormorants (Phalacrocorax penicillatus) in California. J. Zoo Wildl. Med. 1993, 24, 54–62. [Google Scholar]
- Goldstein, T.; Zabka, T.S.; DeLong, R.L.; Wheeler, E.A.; Ylitalo, G.; Bargu, S.; Silver, M.; Leighfield, T.; Van Dolah, F.; Langlois, G.; et al. The role of domoic acid in abortion and premature parturition of California sea lions (Zalophus caufornianus) on San Miguel island, California. J. Wildl. Dis. 2009, 45, 91–108. [Google Scholar] [CrossRef] [Green Version]
- Gobler, C.J. Climate Change and Harmful Algal Blooms: Insights and perspective. Harmful Algae 2020, 91, 101731. [Google Scholar] [CrossRef]
- Lim, H.C.; Tan, S.N.; Teng, S.T.; Lundholm, N.; Orive, E.; David, H.; Quijano-Scheggia, S.; Leong, S.C.Y.; Wolf, M.; Bates, S.S.; et al. Phylogeny and species delineation in the marine diatom Pseudo-nitzschia (Bacillariophyta) using cox1, LSU, and ITS2 rRNA genes: A perspective in character evolution. J. Phycol. 2018, 54, 234–248. [Google Scholar] [CrossRef]
- Dong, H.C.; Lundholm, N.; Teng, S.T.; Li, A.; Wang, C.; Hu, Y.; Li, Y. Occurrence of Pseudo-nitzschia species and associated domoic acid production along the Guangdong coast, South China Sea. Harmful Algae 2020, 98, 101899. [Google Scholar] [CrossRef]
- Holland, P.T.; Selwood, A.I.; Mountfort, D.O.; Wilkins, A.L.; McNabb, P.; Rhodes, L.L.; Doucette, G.J.; Mikulski, C.M.; King, K.L. Isodomoic acid C, an unusual amnesic shellfish poisoning toxin from Pseudo-nitzschia australis. Chem. Res. Toxicol. 2005, 18, 814–816. [Google Scholar] [CrossRef]
- Sawant, P.M.; Holland, P.T.; Mountfort, D.O.; Kerr, D.S. In vivo seizure induction and pharmacological preconditioning by domoic acid and isodomoic acids A., B. and C. Neuropharmacology 2008, 55, 1412–1418. [Google Scholar] [CrossRef]
- Silver, M.W.; Bargu, S.; Coale, S.L.; Benitez-Nelson, C.R.; Garcia, A.C.; Roberts, K.J.; Sekula-Wood, E.; Bruland, K.W.; Coale, K.H. Toxic diatoms and domoic acid in natural and iron enriched waters of the oceanic Pacific. Proc. Natl. Acad. Sci USA 2010, 107, 20762–20767. [Google Scholar] [CrossRef] [Green Version]
- Ferrario, M.E.; Licea, S. Species of the genus Pseudo-nitzschia (Bacillariophyta) in Antarctic waters: Morphology and distribution. Nov. Hedwig. 2006, 130, 1–15. [Google Scholar]
- Kang, S.H.; Fryxell, G.A. Phytoplankton in the Weddell Sea, Antarctica: Composition, abundance and distribution in water-column assemblages of the marginal ice-edge zone during austral autumn. Mar. Biol. Int. J. Life Ocean. Coast. Waters 1993, 116, 335–348. [Google Scholar] [CrossRef]
- Bargu, S.; Powell, C.; Coale, S.; Busman, M.; Doucette, G.; Silver, M. Domoic acid detection in krill: A potential vector in marine food webs. Mar. Ecol. Prog. Ser. 2002, 237, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Hasle, G. Nitzschia and Fragilariopsis species studied in the light and electron microscopes. I. Some marine species of the groups Nitzschiella and Lanceolatae. Det Nor. Vidensk. Akad. Oslo Mat. 1964, 16, 28. [Google Scholar]
- Fryxell, G.A.; Garza, S.A.; Roelke, D.L. Auxospore formation in an Antarctic clone of Nitzschia subcurvate, Hasle. Diatom Res. 1991, 6, 235–245. [Google Scholar] [CrossRef]
- Trick, C.G.; Bill, B.D.; Cochlan, W.P.; Wells, M.L.; Trainer, V.L.; Pickell, L.D. Iron enrichment stimulates toxic diatom production in high-nitrate, low-chlorophyll areas. Proc. Natl. Acad. Sci USA 2010, 107, 5887–5892. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, L.; White, D.; Syhre, M.; Atkinson, M. Pseudo-nitzschia species isolated from New Zealand coastal waters: Domoic acid production in vitro and links with shellfish toxicity. In Harmful Toxic Algal Bloom; Yasumoto, T., Oshima, Y., Fukuyo, Y., Eds.; The Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 1996; pp. 155–158. [Google Scholar]
- Lundholm, N.; Bates, S.S.; Baugh, K.A.; Bill, B.D.; Connell, L.B.; Léger, C.; Trainer, V.L. Cryptic and pseudo-cryptic diversity in diatoms-with descriptions of Pseudo-nitzschia hasleana sp. nov. and P. fryxelliana sp. nov. 1. J. Phycol. 2012, 48, 436–454. [Google Scholar] [CrossRef]
- Trainer, V.L.; Wells, M.L.; Cochlan, W.P.; Trick, C.G.; Bill, B.D.; Baugh, K.A.; Beall, B.F.; Herndon, J.; Lundholmf, N. An ecological study of a massive bloom of toxigenic Pseudo-nitzschia cuspidata off the Washington State coast. Limnol. Oceanogr. 2009, 54, 1461–1474. [Google Scholar] [CrossRef]
- Smith, J. Population dynamics and toxicity of various species of Dinophysis and Nitzschia from the southern Gulf of St. Lawrence. Can. Tech. Rep. Fish. Aquat. Sci. 1991, 1799, 25. [Google Scholar]
- Baugh, K.A.; Bush, J.M.; Bill, B.D.; Lefebvre, K.A.; Trainer, V.L. Estimates of specific toxicity in several Pseudo-nitzschia species from the Washington coast, based on culture and field studies. Afr. J. Mar. Sci. 2006, 28, 403–407. [Google Scholar] [CrossRef]
- Moschandreou, K.K.; Papaefthimiou, D.; Katikou, P.; Kalopesa, E.; Panou, A.; Nikolaidis, G. Morphology, phylogeny and toxin analysis of Pseudo-nitzschia pseudodelicatissima (Bacillariophyceae) isolated from the Thermaikos Gulf, Greece. Phycologia 2010, 49, 260–273. [Google Scholar] [CrossRef]
- Ryabushko, L.I.; Besiktepe, S.; Ediger, D.; Yilmaz, D.; Zenginer, A.; Ryabushko, V.I.; Lee, R.I. Toxic diatom of Pseudo-nitzschia calliantha Lundholm, Moestrup et Hasle from the Black Sea: Morphology, taxonomy, ecology. Mar. Ecol. J. ECOSI Gidrofiz. 2008, 7, 51–60. [Google Scholar]
- Thessen, A.; Bowers, H.; Stoeckera, D. Intra- and interspecies differences in growth and toxicity of Pseudo-nitzschia while using different nitrogen sources. Harmful Algae 2009, 8, 792–810. [Google Scholar] [CrossRef]
- Cerino, F.; Orsini, L.; Sarno, D.; Dell’Aversano, C.; Tartaglione, L.; Zingone, A. The alternation of different morphotypes in the seasonal cycle of the toxic diatom Pseudo-nitzschia galaxiae. Harmful Algae 2005, 4, 33–48. [Google Scholar] [CrossRef]
- Lundholm, N.; Krock, B.; John, U.; Skov, J.; Cheng, J.; Pančić, M.; Wohlrab, S.; Rigby, K.; Nielsen, T.G.; Selander, E.; et al. Induction of domoic acid production in diatoms—Types of grazers and diatoms are important. Harmful Algae 2018, 79, 64–73. [Google Scholar] [CrossRef]
- Geuer, J.K.; Krock, B.; Leefmann, T.; Koch, B.P. Quantification, extractability and stability of dissolved domoic acid within marine dissolved organic matter. Mar. Chem. 2019, 215, 103669. [Google Scholar] [CrossRef]
- Rhodes, L.L.; Holland, P.T.; Adamson, J.; McNabb, P.; Selwood, A.I. Production of a new isomer of domoic acid by New Zealand isolates of the diatom Pseudo-nitzschia australis. In Molluscan Shellfish Safety; Xunta Galicia IOC UNESCO: Paris, France, 2003; Volume 2003, pp. 43–58. [Google Scholar]
- Kotaki, Y.; Furio, E.F.; Satake, M.; Lundholm, N.; Katayama, T.; Koike, K.; Fulgueras, V.P.; Bajarias, F.A.; Takata, Y.; Kobayashi, K.; et al. Production of isodomoic acids A and B as major toxin components of a pennate diatom Nitzschia navis-varingica. Toxicon 2005, 46, 946–953. [Google Scholar] [CrossRef]
- Meda, M.; Kodama, T.; Tanaka, T.; Yoshizumi, H.; Takemoto, T.; Nomoto, K.; Fujita, T. Structures of isodomoic acids A, B and C novel insecticidal amino acids from the red alga Chondria armata. Chem. Pharm. Bull. 1986, 34, 4892–4895. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Qu, P.; Fu, F.; Tennenbaum, N.; Tatters, A.O.; Hutchins, D.A. Understanding the blob bloom: Warming increases toxicity and abundance of the harmful bloom diatom Pseudo-nitzschia in California coastal waters. Harmful Algae 2017, 67, 36–43. [Google Scholar] [CrossRef]
- D’Agostino, V.C.; Degrati, M.; Sastre, V.; Santinelli, N.; Krock, B.; Krohn, T.; Dans, S.L.; Hoffmeyer, M.S. Domoic acid in a marine pelagic food web: Exposure of southern right whales Eubalaena australis to domoic acid on the Península Valdés calving ground, Argentina. Harmful Algae 2017, 68, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Riekkola, L.; Zerbini, A.N.; Andrews, O.; Andrews-Goff, V.; Baker, C.S.; Chandler, D.; Childerhouse, S.; Clapham, P.; Dodémont, R.; Donnelly, D.; et al. Application of a multi-disciplinary approach to reveal population structure and Southern Ocean feeding grounds of humpback whales. Ecol. Indic. 2018, 89, 455–465. [Google Scholar] [CrossRef]
- Krock, B.; Tillmann, U.; John, U.; Cembella, A. LC-MS-MS aboard ship: Tandem mass spectrometry in the search for phycotoxins and novel toxigenic plankton from the North Sea. Anal. Bioanal. Chem. 2008, 392, 797–803. [Google Scholar] [CrossRef] [Green Version]
- Lundholm, N.; Daugbjerg, N.; Moestrup, Ø. Phylogeny of the Bacillariaceae with emphasis on the genus Pseudo-nitzschia (Bacillariophyceae) based on partial LSU rDNA. Eur. J. Phycol. 2002, 37, 115–134. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | pg. DA cell−1 | pg. DA-IC cell−1 | Limit of Detection (pg cell−1) | Cells in DA pellet | Cell Volume (µm3) |
|---|---|---|---|---|---|
| 31-7 | 3.09 × 10−5 | 3.76 × 10−5 | 1.38 × 10−5 | 32.5 × 106 | 235.8 |
| 35-12 | 4.36 × 10−5 | 8.54 × 10−5 | 2.75 × 10−5 | 16.4 × 102 | 162.5 |
| M11-04 | 7.28 × 10−5 | 7.69 × 10−5 | 1.74 × 10−5 | 28.8 × 106 | 171.7 |
| Species | Total pg DA cell−1 | Location | Reference |
|---|---|---|---|
| P. turgidula | Max: 3.3 × 10−2 Min: 0.52 × 10−5 | Tauranga Harbor, New Zealand Ocean Station PAPA (NE Pacific) | [18,19] |
| P. cuspidata | Max: 3.1 × 10−2 Min: 1.9 × 10−2 | Washington State coastal waters | [20,21] |
| P. delicatissima | Min: 0.2 × 10−3 Max: 0.5 × 10−2 | Prince Edward Island, Canada | [22,23] |
| P. pseudodelicatissima | 0.78 × 10−2 | Thermaikos Gulf, Greece | [24] |
| P. calliantha | Max: 4.3 × 10−1 Min: 0.57 × 10−2 | Black Sea Chesapeake Bay, Maryland, USA | [25,26] |
| P. galaxiae | 0.36 × 10−3 | Gulf of Naples, Italy | [27] |
| P. subcurvata | Min. 6.85 × 10−5 max: 1.5 × 10−4 | Southern Ocean | Current study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olesen, A.J.; Leithoff, A.; Altenburger, A.; Krock, B.; Beszteri, B.; Eggers, S.L.; Lundholm, N. First Evidence of the Toxin Domoic Acid in Antarctic Diatom Species. Toxins 2021, 13, 93. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020093
Olesen AJ, Leithoff A, Altenburger A, Krock B, Beszteri B, Eggers SL, Lundholm N. First Evidence of the Toxin Domoic Acid in Antarctic Diatom Species. Toxins. 2021; 13(2):93. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020093
Chicago/Turabian StyleOlesen, Anna J., Anneliese Leithoff, Andreas Altenburger, Bernd Krock, Bánk Beszteri, Sarah Lena Eggers, and Nina Lundholm. 2021. "First Evidence of the Toxin Domoic Acid in Antarctic Diatom Species" Toxins 13, no. 2: 93. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020093