Structural Basis of the Pore-Forming Toxin/Membrane Interaction

 ,
,
Abstract
:1. Introduction
2. Structural Classification and Characterization of the Pore-Forming Toxins
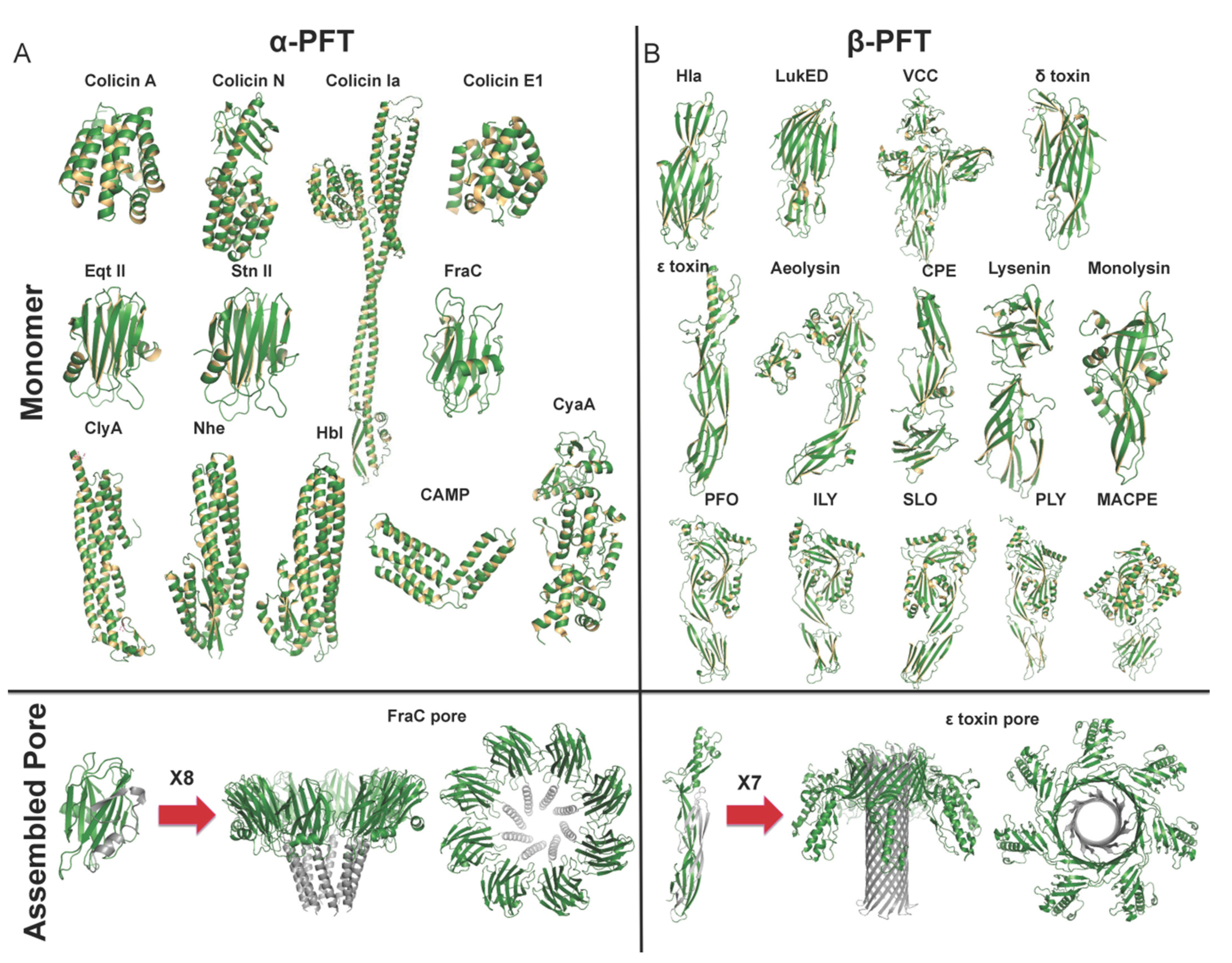
2.1. α-PFTs Family
2.1.1. The Colicin Subfamily
2.1.2. The Cytolysin a Subfamily
2.1.3. The Actinoporin Subfamily
2.1.4. Other α-PFT Families
2.2. β-PFTs Family
2.2.1. The Hemolysin Subfamily
2.2.2. The Aerolysin Subfamily
2.2.3. The Cholesterol-Dependent Cytolysin Subfamily
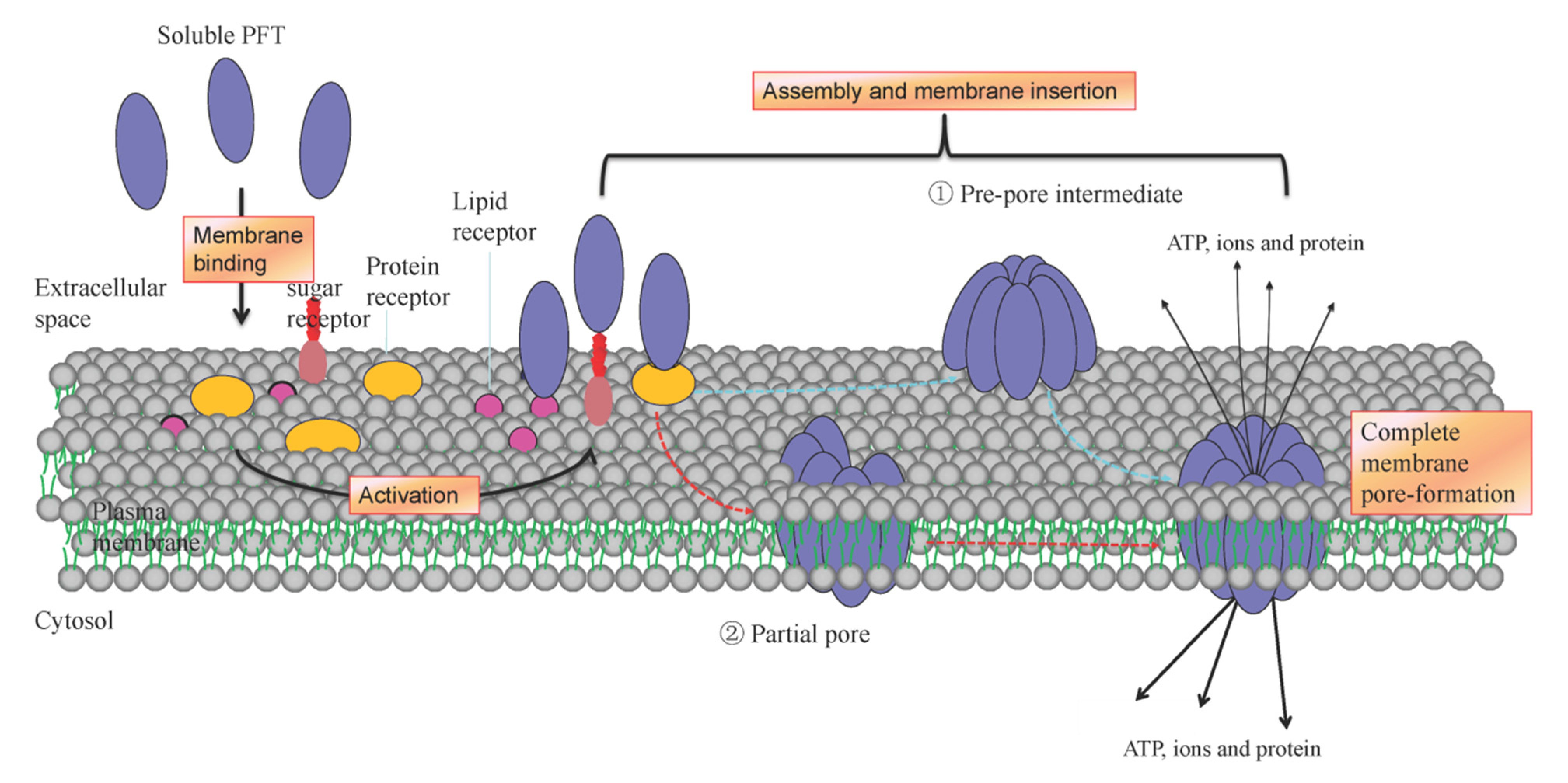
3. Diversification of Interaction of PFTs with Membrane Components
4. Anti-Infection Therapeutic Strategies and Application Targeting PFTs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gonzalez, M.R.; Bischofberger, M.; Pernot, L.; van der Goot, F.G.; Freche, B. Bacterial pore-forming toxins: The (w)hole story? Cell. Mol. Life Sci. 2008, 65, 493–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischofberger, M.; Iacovache, I.; Van der Goot, F.G. Pathogenic pore-forming proteins: Function and host response. Cell Host Microbe 2012, 12, 266–275. [Google Scholar] [CrossRef] [Green Version]
- Dal Peraro, M.; van der Goot, F.G. Pore-forming toxins: Ancient, but never really out of fashion. Nat. Rev. Microbiol. 2016, 14, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Lara-Tejero, M.; Waxham, M.N.; Li, W.; Hu, B.; Galan, J.E.; Liu, J. Visualization of the type III secretion mediated Salmonella-host cell interface using cryo-electron tomography. eLife 2018, 7. [Google Scholar] [CrossRef]
- Fowler, C.C.; Stack, G.; Jiao, X.; Lara-Tejero, M.; Galan, J.E. Alternate subunit assembly diversifies the function of a bacterial toxin. Nat. Commun. 2019, 10, 3684. [Google Scholar] [CrossRef] [Green Version]
- Parker, M.W.; Postma, J.P.; Pattus, F.; Tucker, A.D.; Tsernoglou, D. Refined structure of the pore-forming domain of colicin A at 2.4 A resolution. J. Mol. Biol. 1992, 224, 639–657. [Google Scholar] [CrossRef]
- Vetter, I.R.; Parker, M.W.; Tucker, A.D.; Lakey, J.H.; Pattus, F.; Tsernoglou, D. Crystal structure of a colicin N fragment suggests a model for toxicity. Structure 1998, 6, 863–874. [Google Scholar] [CrossRef] [Green Version]
- Wiener, M.; Freymann, D.; Ghosh, P.; Stroud, R.M. Crystal structure of colicin Ia. Nature 1997, 385, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Elkins, P.; Bunker, A.; Cramer, W.A.; Stauffacher, C.V. A mechanism for toxin insertion into membranes is suggested by the crystal structure of the channel-forming domain of colicin E1. Structure 1997, 5, 443–458. [Google Scholar] [CrossRef] [Green Version]
- Hilsenbeck, J.L.; Park, H.; Chen, G.; Youn, B.; Postle, K.; Kang, C. Crystal structure of the cytotoxic bacterial protein colicin B at 2.5 A resolution. Mol. Microbiol. 2004, 51, 711–720. [Google Scholar] [CrossRef]
- Athanasiadis, A.; Anderluh, G.; Macek, P.; Turk, D. Crystal structure of the soluble form of equinatoxin II, a pore-forming toxin from the sea anemone Actinia equina. Structure 2001, 9, 341–346. [Google Scholar] [CrossRef] [Green Version]
- Mancheno, J.M.; Martin-Benito, J.; Martinez-Ripoll, M.; Gavilanes, J.G.; Hermoso, J.A. Crystal and electron microscopy structures of sticholysin II actinoporin reveal insights into the mechanism of membrane pore formation. Structure 2003, 11, 1319–1328. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Caaveiro, J.M.; Morante, K.; Gonzalez-Manas, J.M.; Tsumoto, K. Structural basis for self-assembly of a cytolytic pore lined by protein and lipid. Nat. Commun. 2015, 6, 6337. [Google Scholar] [CrossRef] [Green Version]
- Wallace, A.J.; Stillman, T.J.; Atkins, A.; Jamieson, S.J.; Bullough, P.A.; Green, J.; Artymiuk, P.J.E. coli hemolysin E (HlyE, ClyA, SheA): X-ray crystal structure of the toxin and observation of membrane pores by electron microscopy. Cell 2000, 100, 265–276. [Google Scholar] [CrossRef]
- Mueller, M.; Grauschopf, U.; Maier, T.; Glockshuber, R.; Ban, N. The structure of a cytolytic alpha-helical toxin pore reveals its assembly mechanism. Nature 2009, 459, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; de Souza Santos, M.; Li, Y.; Tomchick, D.R.; Orth, K. High-resolution cryo-EM structures of the E. coli hemolysin ClyA oligomers. PLoS ONE 2019, 14, e0213423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganash, M.; Phung, D.; Sedelnikova, S.E.; Lindback, T.; Granum, P.E.; Artymiuk, P.J. Structure of the NheA component of the Nhe toxin from Bacillus cereus: Implications for function. PLoS ONE 2013, 8, e74748. [Google Scholar] [CrossRef]
- Madegowda, M.; Eswaramoorthy, S.; Burley, S.K.; Swaminathan, S. X-ray crystal structure of the B component of Hemolysin BL from Bacillus cereus. Proteins 2008, 71, 534–540. [Google Scholar] [CrossRef] [Green Version]
- Brauning, B.; Bertosin, E.; Praetorius, F.; Ihling, C.; Schatt, A.; Adler, A.; Richter, K.; Sinz, A.; Dietz, H.; Groll, M. Structure and mechanism of the two-component alpha-helical pore-forming toxin YaxAB. Nat. Commun. 2018, 9, 1806. [Google Scholar] [CrossRef]
- Schubert, E.; Vetter, I.R.; Prumbaum, D.; Penczek, P.A.; Raunser, S. Membrane insertion of alpha-xenorhabdolysin in near-atomic detail. eLife 2018, 7. [Google Scholar] [CrossRef]
- Jin, T.; Brefo-Mensah, E.; Fan, W.; Zeng, W.; Li, Y.; Zhang, Y.; Palmer, M. Crystal structure of the Streptococcus agalactiae CAMP factor provides insights into its membrane-permeabilizing activity. J. Biol. Chem. 2018, 293, 11867–11877. [Google Scholar] [CrossRef] [Green Version]
- Zeng, W.; Ma, H.; Fan, W.; Yang, Y.; Zhang, C.; Arnaud Kombe Kombe, J.; Fan, X.; Zhang, Y.; Dong, Z.; Shen, Z.; et al. Structure determination of CAMP factor of Mobiluncus curtisii and insights into structural dynamics. Int. J. Biol. Macromol. 2020, 150, 1027–1036. [Google Scholar] [CrossRef]
- Guo, Q.; Shen, Y.; Lee, Y.S.; Gibbs, C.S.; Mrksich, M.; Tang, W.J. Structural basis for the interaction of Bordetella pertussis adenylyl cyclase toxin with calmodulin. EMBO J. 2005, 24, 3190–3201. [Google Scholar] [CrossRef]
- O’Brien, D.P.; Durand, D.; Voegele, A.; Hourdel, V.; Davi, M.; Chamot-Rooke, J.; Vachette, P.; Brier, S.; Ladant, D.; Chenal, A. Calmodulin fishing with a structurally disordered bait triggers CyaA catalysis. PLoS Biol. 2017, 15, e2004486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Kozhaya, L.; Torres, V.J.; Unutmaz, D.; Lu, M. Structure-based discovery of a small-molecule inhibitor of methicillin-resistant Staphylococcus aureus virulence. J. Biol. Chem. 2020, 295, 5944–5959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawara, T.; Yamashita, D.; Kato, K.; Peng, Z.; Ueda, J.; Kaneko, J.; Kamio, Y.; Tanaka, Y.; Yao, M. Structural basis for pore-forming mechanism of staphylococcal alpha-hemolysin. Toxicon 2015, 108, 226–231. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Mikhailova, E.; Cheley, S.; Gu, L.Q.; Montoya, M.; Nagaoka, Y.; Gouaux, E.; Bayley, H. Molecular bases of cyclodextrin adapter interactions with engineered protein nanopores. Proc. Natl. Acad. Sci. USA 2010, 107, 8165–8170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, L.; Hobaugh, M.R.; Shustak, C.; Cheley, S.; Bayley, H.; Gouaux, J.E. Structure of staphylococcal alpha-hemolysin, a heptameric transmembrane pore. Science 1996, 274, 1859–1866. [Google Scholar] [CrossRef]
- Yamashita, D.; Sugawara, T.; Takeshita, M.; Kaneko, J.; Kamio, Y.; Tanaka, I.; Tanaka, Y.; Yao, M. Molecular basis of transmembrane beta-barrel formation of staphylococcal pore-forming toxins. Nat. Commun. 2014, 5, 4897. [Google Scholar] [CrossRef] [Green Version]
- Nocadello, S.; Minasov, G.; Shuvalova, L.; Dubrovska, I.; Sabini, E.; Bagnoli, F.; Grandi, G.; Anderson, W.F. Crystal structures of the components of the Staphylococcus aureus leukotoxin ED. Acta Crystallogr. Sect. D Struct. Biol. 2016, 72, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Savva, C.G.; Fernandes da Costa, S.P.; Bokori-Brown, M.; Naylor, C.E.; Cole, A.R.; Moss, D.S.; Titball, R.W.; Basak, A.K. Molecular architecture and functional analysis of NetB, a pore-forming toxin from Clostridium perfringens. J. Biol. Chem. 2013, 288, 3512–3522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huyet, J.; Naylor, C.E.; Savva, C.G.; Gibert, M.; Popoff, M.R.; Basak, A.K. Structural Insights into Clostridium perfringens Delta Toxin Pore Formation. PLoS ONE 2013, 8, e66673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, R.; Gouaux, E. Crystal structure of the Vibrio cholerae cytolysin (VCC) pro-toxin and its assembly into a heptameric transmembrane pore. J. Mol. Biol. 2005, 350, 997–1016. [Google Scholar] [CrossRef]
- Kaus, K.; Lary, J.W.; Cole, J.L.; Olson, R. Glycan specificity of the Vibrio vulnificus hemolysin lectin outlines evolutionary history of membrane targeting by a toxin family. J. Mol. Biol. 2014, 426, 2800–2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Justin, N.; Walker, N.; Bullifent, H.L.; Songer, G.; Bueschel, D.M.; Jost, H.; Naylor, C.; Miller, J.; Moss, D.S.; Titball, R.W.; et al. The first strain of Clostridium perfringens isolated from an avian source has an alpha-toxin with divergent structural and kinetic properties. Biochemistry 2002, 41, 6253–6262. [Google Scholar] [CrossRef] [PubMed]
- Savva, C.G.; Clark, A.R.; Naylor, C.E.; Popoff, M.R.; Moss, D.S.; Basak, A.K.; Titball, R.W.; Bokori-Brown, M. The pore structure of Clostridium perfringens epsilon toxin. Nat. Commun. 2019, 10, 2641. [Google Scholar] [CrossRef]
- Bokori-Brown, M.; Kokkinidou, M.C.; Savva, C.G.; Fernandes da Costa, S.; Naylor, C.E.; Cole, A.R.; Moss, D.S.; Basak, A.K.; Titball, R.W. Clostridium perfringens epsilon toxin H149A mutant as a platform for receptor binding studies. Protein Sci. A Publ. Protein Soc. 2013, 22, 650–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, A.R.; Gibert, M.; Popoff, M.; Moss, D.S.; Titball, R.W.; Basak, A.K. Clostridium perfringens epsilon-toxin shows structural similarity to the pore-forming toxin aerolysin. Nat. Struct. Mol. Biol. 2004, 11, 797–798. [Google Scholar] [CrossRef]
- Iacovache, I.; De Carlo, S.; Cirauqui, N.; Dal Peraro, M.; van der Goot, F.G.; Zuber, B. Cryo-EM structure of aerolysin variants reveals a novel protein fold and the pore-formation process. Nat. Commun. 2016, 7, 12062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degiacomi, M.T.; Iacovache, I.; Pernot, L.; Chami, M.; Kudryashev, M.; Stahlberg, H.; van der Goot, F.G.; Dal Peraro, M. Molecular assembly of the aerolysin pore reveals a swirling membrane-insertion mechanism. Nat. Chem. Biol. 2013, 9, 623–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, M.W.; Buckley, J.T.; Postma, J.P.; Tucker, A.D.; Leonard, K.; Pattus, F.; Tsernoglou, D. Structure of the Aeromonas toxin proaerolysin in its water-soluble and membrane-channel states. Nature 1994, 367, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Yelland, T.S.; Naylor, C.E.; Bagoban, T.; Savva, C.G.; Moss, D.S.; McClane, B.A.; Blasig, I.E.; Popoff, M.; Basak, A.K. Structure of a C. perfringens enterotoxin mutant in complex with a modified Claudin-2 extracellular loop 2. J. Mol. Biol. 2014, 426, 3134–3147. [Google Scholar] [CrossRef] [PubMed]
- Podobnik, M.; Savory, P.; Rojko, N.; Kisovec, M.; Wood, N.; Hambley, R.; Pugh, J.; Wallace, E.J.; McNeill, L.; Bruce, M.; et al. Crystal structure of an invertebrate cytolysin pore reveals unique properties and mechanism of assembly. Nat. Commun. 2016, 7, 11598. [Google Scholar] [CrossRef] [PubMed]
- Bokori-Brown, M.; Martin, T.G.; Naylor, C.E.; Basak, A.K.; Titball, R.W.; Savva, C.G. Cryo-EM structure of lysenin pore elucidates membrane insertion by an aerolysin family protein. Nat. Commun. 2016, 7, 11293. [Google Scholar] [CrossRef] [Green Version]
- De Colibus, L.; Sonnen, A.F.; Morris, K.J.; Siebert, C.A.; Abrusci, P.; Plitzko, J.; Hodnik, V.; Leippe, M.; Volpi, E.; Anderluh, G.; et al. Structures of lysenin reveal a shared evolutionary origin for pore-forming proteins and its mode of sphingomyelin recognition. Structure 2012, 20, 1498–1507. [Google Scholar] [CrossRef] [Green Version]
- Angulo, I.; Acebron, I.; de las Rivas, B.; Munoz, R.; Rodriguez-Crespo, I.; Menendez, M.; Garcia, P.; Tateno, H.; Goldstein, I.J.; Perez-Agote, B.; et al. High-resolution structural insights on the sugar-recognition and fusion tag properties of a versatile beta-trefoil lectin domain from the mushroom Laetiporus sulphureus. Glycobiology 2011, 21, 1349–1361. [Google Scholar] [CrossRef] [Green Version]
- Leone, P.; Bebeacua, C.; Opota, O.; Kellenberger, C.; Klaholz, B.; Orlov, I.; Cambillau, C.; Lemaitre, B.; Roussel, A. X-ray and Cryo-electron Microscopy Structures of Monalysin Pore-forming Toxin Reveal Multimerization of the Pro-form. J. Biol. Chem. 2015, 290, 13191–13201. [Google Scholar] [CrossRef] [Green Version]
- Rossjohn, J.; Feil, S.C.; McKinstry, W.J.; Tweten, R.K.; Parker, M.W. Structure of a cholesterol-binding, thiol-activated cytolysin and a model of its membrane form. Cell 1997, 89, 685–692. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Huang, B.; Du, H.; Zhang, X.C.; Xu, J.; Li, X.; Rao, Z. Crystal structure of cytotoxin protein suilysin from Streptococcus suis. Protein Cell 2010, 1, 96–105. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, S.L.; Gorman, M.A.; Feil, S.C.; Mulhern, T.D.; Kuiper, M.J.; Ratner, A.J.; Tweten, R.K.; Morton, C.J.; Parker, M.W. Structural Basis for Receptor Recognition by the Human CD59-Responsive Cholesterol-Dependent Cytolysins. Structure 2016, 24, 1488–1498. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.; Brooks, N.J.; Smith, R.A.; Lea, S.M.; Bubeck, D. Structural basis for recognition of the pore-forming toxin intermedilysin by human complement receptor CD59. Cell Rep. 2013, 3, 1369–1377. [Google Scholar] [CrossRef] [Green Version]
- Polekhina, G.; Giddings, K.S.; Tweten, R.K.; Parker, M.W. Insights into the action of the superfamily of cholesterol-dependent cytolysins from studies of intermedilysin. Proc. Natl. Acad. Sci. USA 2005, 102, 600–605. [Google Scholar] [CrossRef] [Green Version]
- Koster, S.; van Pee, K.; Hudel, M.; Leustik, M.; Rhinow, D.; Kuhlbrandt, W.; Chakraborty, T.; Yildiz, O. Crystal structure of listeriolysin O reveals molecular details of oligomerization and pore formation. Nat. Commun. 2014, 5, 3690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feil, S.C.; Lawrence, S.; Mulhern, T.D.; Holien, J.K.; Hotze, E.M.; Farrand, S.; Tweten, R.K.; Parker, M.W. Structure of the lectin regulatory domain of the cholesterol-dependent cytolysin lectinolysin reveals the basis for its lewis antigen specificity. Structure 2012, 20, 248–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourdeau, R.W.; Malito, E.; Chenal, A.; Bishop, B.L.; Musch, M.W.; Villereal, M.L.; Chang, E.B.; Mosser, E.M.; Rest, R.F.; Tang, W.J. Cellular functions and X-ray structure of anthrolysin O, a cholesterol-dependent cytolysin secreted by Bacillus anthracis. J. Biol. Chem. 2009, 284, 14645–14656. [Google Scholar] [CrossRef] [Green Version]
- Feil, S.C.; Ascher, D.B.; Kuiper, M.J.; Tweten, R.K.; Parker, M.W. Structural studies of Streptococcus pyogenes streptolysin O provide insights into the early steps of membrane penetration. J. Mol. Biol. 2014, 426, 785–792. [Google Scholar] [CrossRef] [Green Version]
- van Pee, K.; Neuhaus, A.; D’Imprima, E.; Mills, D.J.; Kuhlbrandt, W.; Yildiz, O. CryoEM structures of membrane pore and prepore complex reveal cytolytic mechanism of Pneumolysin. eLife 2017, 6. [Google Scholar] [CrossRef]
- Marshall, J.E.; Faraj, B.H.; Gingras, A.R.; Lonnen, R.; Sheikh, M.A.; El-Mezgueldi, M.; Moody, P.C.; Andrew, P.W.; Wallis, R. The Crystal Structure of Pneumolysin at 2.0 A Resolution Reveals the Molecular Packing of the Pre-pore Complex. Sci. Rep. 2015, 5, 13293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, S.L.; Feil, S.C.; Morton, C.J.; Farrand, A.J.; Mulhern, T.D.; Gorman, M.A.; Wade, K.R.; Tweten, R.K.; Parker, M.W. Crystal structure of Streptococcus pneumoniae pneumolysin provides key insights into early steps of pore formation. Sci. Rep. 2015, 5, 14352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosado, C.J.; Buckle, A.M.; Law, R.H.; Butcher, R.E.; Kan, W.T.; Bird, C.H.; Ung, K.; Browne, K.A.; Baran, K.; Bashtannyk-Puhalovich, T.A.; et al. A common fold mediates vertebrate defense and bacterial attack. Science 2007, 317, 1548–1551. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Abdubek, P.; Astakhova, T.; Axelrod, H.L.; Bakolitsa, C.; Cai, X.; Carlton, D.; Chen, C.; Chiu, H.J.; Clayton, T.; et al. Structure of a membrane-attack complex/perforin (MACPF) family protein from the human gut symbiont Bacteroides thetaiotaomicron. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 1297–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cascales, E.; Buchanan, S.K.; Duche, D.; Kleanthous, C.; Lloubes, R.; Postle, K.; Riley, M.; Slatin, S.; Cavard, D. Colicin biology. Microbiol. Mol. Biol. Rev. 2007, 71, 158–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakey, J.H.; Slatin, S.L. Pore-forming colicins and their relatives. Curr. Top. Microbiol. Immunol. 2001, 257, 131–161. [Google Scholar] [CrossRef]
- Pulagam, L.P.; Steinhoff, H.J. Acidic pH-induced membrane insertion of colicin A into E. coli natural lipids probed by site-directed spin labeling. J. Mol. Biol. 2013, 425, 1782–1794. [Google Scholar] [CrossRef] [PubMed]
- Ridley, H.; Johnson, C.L.; Lakey, J.H. Interfacial interactions of pore-forming colicins. Adv. Exp. Med. Biol. 2010, 677, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Dolejsova, T.; Sokol, A.; Bosak, J.; Smajs, D.; Konopasek, I.; Mikusova, G.; Fiser, R. Colicin U from Shigella boydii Forms Voltage-Dependent Pores. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef]
- Oh, K.J.; Senzel, L.; Collier, R.J.; Finkelstein, A. Translocation of the catalytic domain of diphtheria toxin across planar phospholipid bilayers by its own T domain. Proc. Natl. Acad. Sci. USA 1999, 96, 8467–8470. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Wang, B.C.; Yu, Z.; Sun, M. Structural insights into Bacillus thuringiensis Cry, Cyt and parasporin toxins. Toxins 2014, 6, 2732–2770. [Google Scholar] [CrossRef] [Green Version]
- Barta, M.L.; Dickenson, N.E.; Patil, M.; Keightley, A.; Wyckoff, G.J.; Picking, W.D.; Picking, W.L.; Geisbrecht, B.V. The structures of coiled-coil domains from type III secretion system translocators reveal homology to pore-forming toxins. J. Mol. Biol. 2012, 417, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Westphal, D.; Dewson, G.; Czabotar, P.E.; Kluck, R.M. Molecular biology of Bax and Bak activation and action. Biochim. Biophys. Acta 2011, 1813, 521–531. [Google Scholar] [CrossRef] [Green Version]
- Hunt, S.; Green, J.; Artymiuk, P.J. Hemolysin E (HlyE, ClyA, SheA) and related toxins. Adv. Exp. Med. Biol. 2010, 677, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Brauning, B.; Groll, M. Structural and Mechanistic Features of ClyA-Like alpha-Pore-Forming Toxins. Toxins 2018, 10, 343. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.S.; Churchill-Angus, A.M.; Davies, S.P.; Sedelnikova, S.E.; Tzokov, S.B.; Rafferty, J.B.; Bullough, P.A.; Bisson, C.; Baker, P.J. (Identification and structural analysis of the tripartite alpha-pore forming toxin of Aeromonas hydrophila). Nat. Commun. 2019, 10, 2900. [Google Scholar] [CrossRef]
- Fahie, M.; Romano, F.B.; Chisholm, C.; Heuck, A.P.; Zbinden, M.; Chen, M. A non-classical assembly pathway of Escherichia coli pore-forming toxin cytolysin A. J. Biol. Chem. 2013, 288, 31042–31051. [Google Scholar] [CrossRef] [Green Version]
- Kristan, K.C.; Viero, G.; Dalla Serra, M.; Macek, P.; Anderluh, G. Molecular mechanism of pore formation by actinoporins. Toxicon 2009, 54, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Yanagihara, I.; Nakahira, K.; Yamane, T.; Kaieda, S.; Mayanagi, K.; Hamada, D.; Fukui, T.; Ohnishi, K.; Kajiyama, S.; Shimizu, T.; et al. Structure and functional characterization of Vibrio parahaemolyticus thermostable direct hemolysin. J. Biol. Chem. 2010, 285, 16267–16274. [Google Scholar] [CrossRef] [Green Version]
- Kundu, N.; Tichkule, S.; Pandit, S.B.; Chattopadhyay, K. Disulphide bond restrains the C-terminal region of thermostable direct hemolysin during folding to promote oligomerization. Biochem. J. 2017, 474, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.A.; Rojko, N.; Cronin, B.; Anderluh, G.; Wallace, M.I. Photobleaching reveals heterogeneous stoichiometry for equinatoxin II oligomers. ChemBioChem 2014, 15, 2139–2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechaly, A.E.; Bellomio, A.; Gil-Carton, D.; Morante, K.; Valle, M.; Gonzalez-Manas, J.M.; Guerin, D.M. Structural insights into the oligomerization and architecture of eukaryotic membrane pore-forming toxins. Structure 2011, 19, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Mondal, A.K.; Sreekumar, A.; Kundu, N.; Kathuria, R.; Verma, P.; Gandhi, S.; Chattopadhyay, K. Structural Basis and Functional Implications of the Membrane Pore-Formation Mechanisms of Bacterial Pore-Forming Toxins. Adv. Exp. Med. Biol. 2018, 1112, 281–291. [Google Scholar] [CrossRef]
- Linhartova, I.; Bumba, L.; Masin, J.; Basler, M.; Osicka, R.; Kamanova, J.; Prochazkova, K.; Adkins, I.; Hejnova-Holubova, J.; Sadilkova, L.; et al. RTX proteins: A highly diverse family secreted by a common mechanism. FEMS Microbiol. Rev. 2010, 34, 1076–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudryashova, E.; Heisler, D.; Zywiec, A.; Kudryashov, D.S. Thermodynamic properties of the effector domains of MARTX toxins suggest their unfolding for translocation across the host membrane. Mol. Microbiol. 2014, 92, 1056–1071. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, D.P.; Hernandez, B.; Durand, D.; Hourdel, V.; Sotomayor-Perez, A.C.; Vachette, P.; Ghomi, M.; Chamot-Rooke, J.; Ladant, D.; Brier, S.; et al. Structural models of intrinsically disordered and calcium-bound folded states of a protein adapted for secretion. Sci. Rep. 2015, 5, 14223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, D.P.; Perez, A.C.S.; Karst, J.; Cannella, S.E.; Enguene, V.Y.N.; Hessel, A.; Raoux-Barbot, D.; Voegele, A.; Subrini, O.; Davi, M.; et al. Calcium-dependent disorder-to-order transitions are central to the secretion and folding of the CyaA toxin of Bordetella pertussis, the causative agent of whooping cough. Toxicon 2018, 149, 37–44. [Google Scholar] [CrossRef]
- Baumann, U.; Wu, S.; Flaherty, K.M.; McKay, D.B. Three-dimensional structure of the alkaline protease of Pseudomonas aeruginosa: A two-domain protein with a calcium binding parallel beta roll motif. Embo J. 1993, 12, 3357–3364. [Google Scholar] [CrossRef]
- Ostolaza, H.; Gonzalez-Bullon, D.; Uribe, K.B.; Martin, C.; Amuategi, J.; Fernandez-Martinez, X. Membrane Permeabilization by Pore-Forming RTX Toxins: What Kind of Lesions Do These Toxins Form? Toxins 2019, 11, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DuMont, A.L.; Yoong, P.; Liu, X.; Day, C.J.; Chumbler, N.M.; James, D.B.; Alonzo, F., 3rd; Bode, N.J.; Lacy, D.B.; Jennings, M.P.; et al. Identification of a crucial residue required for Staphylococcus aureus LukAB cytotoxicity and receptor recognition. Infect. Immun. 2014, 82, 1268–1276. [Google Scholar] [CrossRef] [Green Version]
- De, S.; Olson, R. Crystal structure of the Vibrio cholerae cytolysin heptamer reveals common features among disparate pore-forming toxins. Proc. Natl. Acad. Sci. USA 2011, 108, 7385–7390. [Google Scholar] [CrossRef] [Green Version]
- Unno, H.; Goda, S.; Hatakeyama, T. Hemolytic lectin CEL-III heptamerizes via a large structural transition from alpha-helices to a beta-barrel during the transmembrane pore formation process. J. Biol. Chem. 2014, 289, 12805–12812. [Google Scholar] [CrossRef] [Green Version]
- Tweten, R.K.; Hotze, E.M.; Wade, K.R. The Unique Molecular Choreography of Giant Pore Formation by the Cholesterol-Dependent Cytolysins of Gram-Positive Bacteria. Annu. Rev. Microbiol. 2015, 69, 323–340. [Google Scholar] [CrossRef]
- Reboul, C.F.; Whisstock, J.C.; Dunstone, M.A. Giant MACPF/CDC pore forming toxins: A class of their own. Biochim. Biophys. Acta 2016, 1858, 475–486. [Google Scholar] [CrossRef]
- Johnson, B.B.; Heuck, A.P. Perfringolysin O structure and mechanism of pore formation as a paradigm for cholesterol-dependent cytolysins. Sub Cell. Biochem. 2014, 80, 63–81. [Google Scholar] [CrossRef] [Green Version]
- Shannon, J.G.; Ross, C.L.; Koehler, T.M.; Rest, R.F. Characterization of anthrolysin O, the Bacillus anthracis cholesterol-dependent cytolysin. Infect. Immun. 2003, 71, 3183–3189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, J.C.; Johnstone, B.A.; Lawrence, S.L.; Morton, C.J.; Christie, M.P.; Parker, M.W.; Tweten, R.K. A Key Motif in the Cholesterol-Dependent Cytolysins Reveals a Large Family of Related Proteins. mBio 2020, 11. [Google Scholar] [CrossRef]
- Wade, K.R.; Lawrence, S.L.; Farrand, A.J.; Hotze, E.M.; Kuiper, M.J.; Gorman, M.A.; Christie, M.P.; Panjikar, S.; Morton, C.J.; Parker, M.W.; et al. The Structural Basis for a Transition State That Regulates Pore Formation in a Bacterial Toxin. mBio 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Pee, K.; Mulvihill, E.; Muller, D.J.; Yildiz, O. Unraveling the Pore-Forming Steps of Pneumolysin from Streptococcus pneumoniae. Nano Lett. 2016, 16, 7915–7924. [Google Scholar] [CrossRef] [PubMed]
- Leung, C.; Dudkina, N.V.; Lukoyanova, N.; Hodel, A.W.; Farabella, I.; Pandurangan, A.P.; Jahan, N.; Pires Damaso, M.; Osmanovic, D.; Reboul, C.F.; et al. Stepwise visualization of membrane pore formation by suilysin, a bacterial cholesterol-dependent cytolysin. eLife 2014, 3, e04247. [Google Scholar] [CrossRef]
- Sato, T.K.; Tweten, R.K.; Johnson, A.E. Disulfide-bond scanning reveals assembly state and beta-strand tilt angle of the PFO beta-barrel. Nat. Chem. Biol. 2013, 9, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Czajkowsky, D.M.; Hotze, E.M.; Shao, Z.; Tweten, R.K. Vertical collapse of a cytolysin prepore moves its transmembrane beta-hairpins to the membrane. Embo J. 2004, 23, 3206–3215. [Google Scholar] [CrossRef]
- Levan, S.; De, S.; Olson, R. Vibrio cholerae cytolysin recognizes the heptasaccharide core of complex N-glycans with nanomolar affinity. J. Mol. Biol. 2013, 425, 944–957. [Google Scholar] [CrossRef] [Green Version]
- Rai, A.K.; Paul, K.; Chattopadhyay, K. Functional mapping of the lectin activity site on the beta-prism domain of vibrio cholerae cytolysin: Implications for the membrane pore-formation mechanism of the toxin. J. Biol. Chem. 2013, 288, 1665–1673. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Takahashi, J.; Seno, T.; Tani, Y.; Honda, T. Analysis of receptor for Vibrio cholerae El tor hemolysin with a monoclonal antibody that recognizes glycophorin B of human erythrocyte membrane. Infect. Immun. 1999, 67, 5332–5337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diep, D.B.; Nelson, K.L.; Raja, S.M.; Pleshak, E.N.; Buckley, J.T. Glycosylphosphatidylinositol anchors of membrane glycoproteins are binding determinants for the channel-forming toxin aerolysin. J. Biol. Chem. 1998, 273, 2355–2360. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Ohishi, K.; Inoue, N.; Kang, J.Y.; Shime, H.; Horiguchi, Y.; van der Goot, F.G.; Sugimoto, N.; Kinoshita, T. Requirement of N-glycan on GPI-anchored proteins for efficient binding of aerolysin but not Clostridium septicum alpha-toxin. EMBO J. 2002, 21, 5047–5056. [Google Scholar] [CrossRef]
- Ivie, S.E.; McClain, M.S. Identification of amino acids important for binding of Clostridium perfringens epsilon toxin to host cells and to HAVCR1. Biochemistry 2012, 51, 7588–7595. [Google Scholar] [CrossRef] [Green Version]
- Giddings, K.S.; Zhao, J.; Sims, P.J.; Tweten, R.K. Human CD59 is a receptor for the cholesterol-dependent cytolysin intermedilysin. Nat. Struct. Mol. Biol. 2004, 11, 1173–1178. [Google Scholar] [CrossRef]
- Li, Y.; Zeng, W.; Li, Y.; Fan, W.; Ma, H.; Fan, X.; Jiang, J.; Brefo-Mensah, E.; Zhang, Y.; Yang, M.; et al. Structure determination of the CAMP factor of Streptococcus agalactiae with the aid of an MBP tag and insights into membrane-surface attachment. Acta Crystallogr. Sect. D Struct. Biol. 2019, 75, 772–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, C.L.; Ridley, H.; Marchetti, R.; Silipo, A.; Griffin, D.C.; Crawford, L.; Bonev, B.; Molinaro, A.; Lakey, J.H. The antibacterial toxin colicin N binds to the inner core of lipopolysaccharide and close to its translocator protein. Mol. Microbiol. 2014, 92, 440–452. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Zheng, H.; Derebe, M.G.; Callenberg, K.M.; Partch, C.L.; Rollins, D.; Propheter, D.C.; Rizo, J.; Grabe, M.; Jiang, Q.X.; et al. Antibacterial membrane attack by a pore-forming intestinal C-type lectin. Nature 2014, 505, 103–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacovache, I.; Paumard, P.; Scheib, H.; Lesieur, C.; Sakai, N.; Matile, S.; Parker, M.W.; van der Goot, F.G. A rivet model for channel formation by aerolysin-like pore-forming toxins. EMBO J. 2006, 25, 457–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakai, Y.; Sakurai, Y.; Yamaji, A.; Asou, H.; Umeda, M.; Uyemura, K.; Itoh, K. Lysenin-sphingomyelin binding at the surface of oligodendrocyte lineage cells increases during differentiation in vitro. J. Neurosci. Res. 2000, 62, 521–529. [Google Scholar] [CrossRef]
- Zitzer, A.; Westover, E.J.; Covey, D.F.; Palmer, M. Differential interaction of the two cholesterol-dependent, membrane-damaging toxins, streptolysin O and Vibrio cholerae cytolysin, with enantiomeric cholesterol. FEBS Lett. 2003, 553, 229–231. [Google Scholar] [CrossRef] [Green Version]
- Rai, A.K.; Chattopadhyay, K. Revisiting the membrane interaction mechanism of a membrane-damaging beta-barrel pore-forming toxin Vibrio cholerae cytolysin. Mol. Microbiol. 2015, 97, 1051–1062. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q.; London, E. Altering hydrophobic sequence lengths shows that hydrophobic mismatch controls affinity for ordered lipid domains (rafts) in the multitransmembrane strand protein perfringolysin O. J. Biol. Chem. 2013, 288, 1340–1352. [Google Scholar] [CrossRef] [Green Version]
- Kozorog, M.; Sani, M.A.; Lenarcic Zivkovic, M.; Ilc, G.; Hodnik, V.; Separovic, F.; Plavec, J.; Anderluh, G. (19)F NMR studies provide insights into lipid membrane interactions of listeriolysin O, a pore forming toxin from Listeria monocytogenes. Sci. Rep. 2018, 8, 6894. [Google Scholar] [CrossRef]
- Farrand, A.J.; Hotze, E.M.; Sato, T.K.; Wade, K.R.; Wimley, W.C.; Johnson, A.E.; Tweten, R.K. The Cholesterol-dependent Cytolysin Membrane-binding Interface Discriminates Lipid Environments of Cholesterol to Support beta-Barrel Pore Insertion. J. Biol. Chem. 2015, 290, 17733–17744. [Google Scholar] [CrossRef] [Green Version]
- Dowd, K.J.; Farrand, A.J.; Tweten, R.K. The cholesterol-dependent cytolysin signature motif: A critical element in the allosteric pathway that couples membrane binding to pore assembly. PLoS Pathog. 2012, 8, e1002787. [Google Scholar] [CrossRef]
- Farrand, A.J.; LaChapelle, S.; Hotze, E.M.; Johnson, A.E.; Tweten, R.K. Only two amino acids are essential for cytolytic toxin recognition of cholesterol at the membrane surface. Proc. Natl. Acad. Sci. USA 2010, 107, 4341–4346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilke, G.A.; Bubeck Wardenburg, J. Role of a disintegrin and metalloprotease 10 in Staphylococcus aureus alpha-hemolysin-mediated cellular injury. Proc. Natl. Acad. Sci. USA 2010, 107, 13473–13478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaan, A.N.; Henry, T.; van Rooijen, W.J.M.; Perret, M.; Badiou, C.; Aerts, P.C.; Kemmink, J.; de Haas, C.J.C.; van Kessel, K.P.M.; Vandenesch, F.; et al. The staphylococcal toxin Panton-Valentine Leukocidin targets human C5a receptors. Cell Host Microbe 2013, 13, 584–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruggisser, J.; Tarek, B.; Wyder, M.; Muller, P.; von Ballmoos, C.; Witz, G.; Enzmann, G.; Deutsch, U.; Engelhardt, B.; Posthaus, H. CD31 (PECAM-1) Serves as the Endothelial Cell-Specific Receptor of Clostridium perfringens beta-Toxin. Cell Host Microbe 2020, 28, 69–78 e66. [Google Scholar] [CrossRef]
- Zilnyte, M.; Venclovas, C.; Zvirbliene, A.; Pleckaityte, M. The cytolytic activity of vaginolysin strictly depends on cholesterol and is potentiated by human CD59. Toxins 2015, 7, 110–128. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Qiu, J.; Zhang, Y.; Lu, C.; Dai, X.; Wang, J.; Li, H.; Wang, X.; Tan, W.; Luo, M.; et al. Oroxylin A inhibits hemolysis via hindering the self-assembly of alpha-hemolysin heptameric transmembrane pore. PLoS Comput. Biol. 2013, 9, e1002869. [Google Scholar] [CrossRef] [PubMed]
- Vivekananda, J.; Salgado, C.; Millenbaugh, N.J. DNA aptamers as a novel approach to neutralize Staphylococcus aureus alpha-toxin. Biochem. Biophys. Res. Commun. 2014, 444, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Shen, X.; Wei, Y.; Si, X.; Deng, X.; Wang, J. Quercetin reduces Streptococcus suis virulence by inhibiting suilysin activity and inflammation. Int. Immunopharmacol. 2019, 69, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Guo, Z. Glycosylphosphatidylinositols are potential targets for the development of novel inhibitors for aerolysin-type of pore-forming bacterial toxins. Med. Res. Rev. 2010, 30, 258–269. [Google Scholar] [CrossRef]
- Ortines, R.V.; Liu, H.; Cheng, L.I.; Cohen, T.S.; Lawlor, H.; Gami, A.; Wang, Y.; Dillen, C.A.; Archer, N.K.; Miller, R.J.; et al. Neutralizing Alpha-Toxin Accelerates Healing of Staphylococcus aureus-Infected Wounds in Nondiabetic and Diabetic Mice. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Alonzo, F., 3rd; Kozhaya, L.; Rawlings, S.A.; Reyes-Robles, T.; DuMont, A.L.; Myszka, D.G.; Landau, N.R.; Unutmaz, D.; Torres, V. CCR5 is a receptor for Staphylococcus aureus leukotoxin ED. Nature 2013, 493, 51–55. [Google Scholar]
- Schwiering, M.; Husmann, M.; Hellmann, N. P2X-Receptor Antagonists Inhibit the Interaction of S. aureus Hemolysin A with Membranes. Toxins 2017, 9, 332. [Google Scholar] [CrossRef] [Green Version]
- Fernandes da Costa, S.P.; Savva, C.G.; Bokori-Brown, M.; Naylor, C.E.; Moss, D.S.; Basak, A.K.; Titball, R.W. Identification of a key residue for oligomerisation and pore-formation of Clostridium perfringens NetB. Toxins 2014, 6, 1049–1061. [Google Scholar] [CrossRef] [Green Version]
- Bokori-Brown, M.; Hall, C.A.; Vance, C.; Fernandes da Costa, S.P.; Savva, C.G.; Naylor, C.E.; Cole, A.R.; Basak, A.K.; Moss, D.S.; Titball, R.W. Clostridium perfringens epsilon toxin mutant Y30A-Y196A as a recombinant vaccine candidate against enterotoxemia. Vaccine 2014, 32, 2682–2687. [Google Scholar] [CrossRef] [Green Version]
- Lohith, G.K.; Kingston, J.J.; Singh, A.K.; Murali, H.S.; Batra, H.V. Evaluation of recombinant leukocidin domain of VvhA exotoxin of Vibrio vulnificus as an effective toxoid in mouse model. Immunol. Lett. 2015, 167, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Cockeran, R.; Steel, H.C.; Theron, A.J.; Mitchell, T.J.; Feldman, C.; Anderson, R. Characterization of the interactions of the pneumolysoid, Delta6 PLY, with human neutrophils in vitro. Vaccine 2011, 29, 8780–8782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douce, G.; Ross, K.; Cowan, G.; Ma, J.; Mitchell, T.J. Novel mucosal vaccines generated by genetic conjugation of heterologous proteins to pneumolysin (PLY) from Streptococcus pneumoniae. Vaccine 2010, 28, 3231–3237. [Google Scholar] [CrossRef] [PubMed]
- Mann, B.; Thornton, J.; Heath, R.; Wade, K.R.; Tweten, R.K.; Gao, G.; El Kasmi, K.; Jordan, J.B.; Mitrea, D.M.; Kriwacki, R.; et al. Broadly protective protein-based pneumococcal vaccine composed of pneumolysin toxoid-CbpA peptide recombinant fusion protein. J. Infect. Dis. 2014, 209, 1116–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.M.; Zhang, L. Nanotoxoid Vaccines. Nano Today 2014, 9, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Koo, J.; Escajadillo, T.; Zhang, L.; Nizet, V.; Lawrence, S.M. Erythrocyte-Coated Nanoparticles Block Cytotoxic Effects of Group B Streptococcus beta-Hemolysin/Cytolysin. Front. Pediatr. 2019, 7, 410. [Google Scholar] [CrossRef] [Green Version]
- Narva, K.E.; Wang, N.X.; Herman, R. Safety considerations derived from Cry34Ab1/Cry35Ab1 structure and function. J. Invertebr. Pathol. 2017, 142, 27–33. [Google Scholar] [CrossRef]
- Dhillon, S. Correction to: Moxetumomab Pasudotox: First Global Approval. Drugs 2019, 79, 105. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| PFT | Family | Receptor | PDB | Organisms | References |
|---|---|---|---|---|---|
| α-PFT | |||||
| Colicin A | Colicins | IM/OM | 1COL | E. coli | [6] |
| Colicin N | Colicins | IM/LPS-OM | 1A87 | E. coli | [7] |
| Colicin Ia | Colicins | IM/OM | 1CII | E. coli | [8] |
| Colicin E1 | Colicins | IM/OM | 2I88 | E. coli | [9] |
| Colicin B | Colicins | IM/OM | 1RH1 | E. coli | [10] |
| EquinatoxinII (EqtII) | Actinoporins | Sphingomyelin | 1IAZ | A. equina | [11] |
| SticholysinII (StnII) | Actinoporins | Sphingomyelin | 1GWY | S. helianthus | [12] |
| Fragaceatoxin C (FraC) | Actinoporins | Sphingomyelin | 4TSL, 4TSY | A. fragacea | [13] |
| Hemolysin E (HlyE) | CytolysinA | Cholesterol | 1QOY, 2WCD, 6MRT | E. coli | [14,15,16] |
| Non-hemolytic tripartite enterotoxin (Nhe) | Cytolysin A | Cholesterol | 4K1P | B. cereus | [17] |
| HaemolysinBL (Hbl) | Cytolysin A | Cholesterol | 2NRJ | B. cereus | [18] |
| YaxAB | Cytolysin A | 6EK4, 6EK7, 6EK8, 6EL1 | Y. enterolitica | [19] | |
| XaxAB | Cytolysin A | 6GY6 | X. nematophila | [20] | |
| CAMP | CAMP | GPI-anchored proteins | 5H6I 6JLC | S. agalactiae M. curtisii | [21,22] |
| α-hemolysin (HlyA) | RTX | β2 integrin | E. coli | ||
| Adenylate cyclase-hemolysin toxin (CyaA) | RTX | - | 2COL SASDCK9 SASDCL9 | B. pertussis | [23,24] |
| MARTX | RTX | - | - | A. hydrophila | |
| β-PFT | |||||
| α-haemolysin (Hla) | Hemolysin | PC/ADAM10/ disintegrin | 3M2L, 3M4D, 7AHL, 4YHD, 6U49, 6U4P, 6U3T | S. aureus | [25,26,27,28] |
| γ-hemolysin (Hlg) | Hemolysin | PC | 4P1Y | S. aureus | [29] |
| Leukocidin (HlgACB, LukED) | Hemolysin | CCR5, CXCR1, CXCR2, CCR2,C5aR, DARC | 3ROH, 4Q7G | S. aureus | [30] |
| Necrotic enteritis toxin B (NetB) | Hemolysin | Cholesterol | 4H56 | C. perfringens | [31] |
| δ toxin | Hemolysin | Monosialicganglioside 2 (GM2) | 2YGT | C. perfringens | [32] |
| Vibrio cholerae cytolysin (VCC) | Hemolysin | Glycoconjugates | 1XEZ | V. choleraes | [33] |
| Vibrio vulnificus hemolysin (VVH) | Hemolysin | gangliosides, N-acetyl-D-galactosamine, N-acetyl-D-lactosamine | 4OWJ, 4OWK, 4OWL | V. vulnificus | [34] |
| α-toxin | Aerolysin | GPI-anchored proteins | 1KHO | C. perfringens | [35] |
| ε-toxin (Etx) | Aerolysin | HAVCR1, MAL | 1UYJ, 6RB9, 3ZJX | C. perfringens | [36,37,38] |
| Aerolysin | Aerolysin | GPI-anchored proteins (CD52) | 1PRE, 3C0M, 3C0N, 3C0O, 5JZT | Aeromonas spp. | [39,40,41] |
| Hydralysin (Hln) | Aerolysin | - | - | Cnidaria spp. | |
| Enterotoxin (CPE) | Aerolysin | Claudin | 3ZIW | C. perfringens | [42] |
| Lysenin | Aerolysin | Sphingomyelin | 3ZXD, 3ZXG, 3ZX7, 5EC5, 5GAQ | E. fetida | [43,44,45] |
| Hemolytic lectin (LSL) | Aerolysin | Carbohydrates | 2Y9F | L. sulphureus | [46] |
| Monalysin | Aerolysin | 4MKO, 4MJT | P. entomophila | [47] | |
| Perfringolysin O (PFO) | CDCs | Cholesterol | 1PFO | C. perfringens | [48] |
| Suilysin (SLY) | CDCs | Cholesterol | 3HVN | S. suis | [49] |
| Intermedilysin (ILY) | CDCs | Cholesterol, CD59 | 1S3R, 4BIK, 5IMW, 5IMT | S. intermedius | [50,51,52] |
| Listeriolysin (LLO) | CDCs | Cholesterol | 4CDB | L. monocytoge-nes | [53] |
| Lectinolysin (LLY) | CDCs | Cholesterol, CD59 | 3LEI | S. mitis | [54] |
| Anthrolysin O (ALO) | CDCs | Cholesterol | 3CQF | B. anthracis | [55] |
| Streptolysin O (SLO) | CDCs | Cholesterol | 4HSC | S. pyogenes | [56] |
| Pneumolysin (PLY) | CDCs | Cholesterol | 4ZGH, 5AOF, 5LY6, 5CR6, 5AOE | S. pneumoniae | [57,58,59] |
| Vaginolysin (VLY) | CDCs | Cholesterol | 5IMY, 5IMT, 5IMW | G. vaginalis | [50] |
| Plu-MACPE | MACPF | - | 2QP2 | P. luminescens | [60] |
| Bth-MACPE | MACPF | - | 3KK7 | B. thetaiotaomicron | [61] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Li, Y.; Mengist, H.M.; Shi, C.; Zhang, C.; Wang, B.; Li, T.; Huang, Y.; Xu, Y.; Jin, T. Structural Basis of the Pore-Forming Toxin/Membrane Interaction. Toxins 2021, 13, 128. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020128
Li Y, Li Y, Mengist HM, Shi C, Zhang C, Wang B, Li T, Huang Y, Xu Y, Jin T. Structural Basis of the Pore-Forming Toxin/Membrane Interaction. Toxins. 2021; 13(2):128. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020128
Chicago/Turabian StyleLi, Yajuan, Yuelong Li, Hylemariam Mihiretie Mengist, Cuixiao Shi, Caiying Zhang, Bo Wang, Tingting Li, Ying Huang, Yuanhong Xu, and Tengchuan Jin. 2021. "Structural Basis of the Pore-Forming Toxin/Membrane Interaction" Toxins 13, no. 2: 128. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13020128