Assessing the Single and Combined Toxicity of Chlorantraniliprole and Bacillus thuringiensis (GO33A) against Four Selected Strains of Plutella xylostella (Lepidoptera: Plutellidae), and a Gene Expression Analysis


 ,
,
Abstract
:1. Introduction
2. Results
2.1. The Resistance Level of Greenhouse Strains of P. xylostella to Three Insecticides
2.2. Joint Action of Combinations
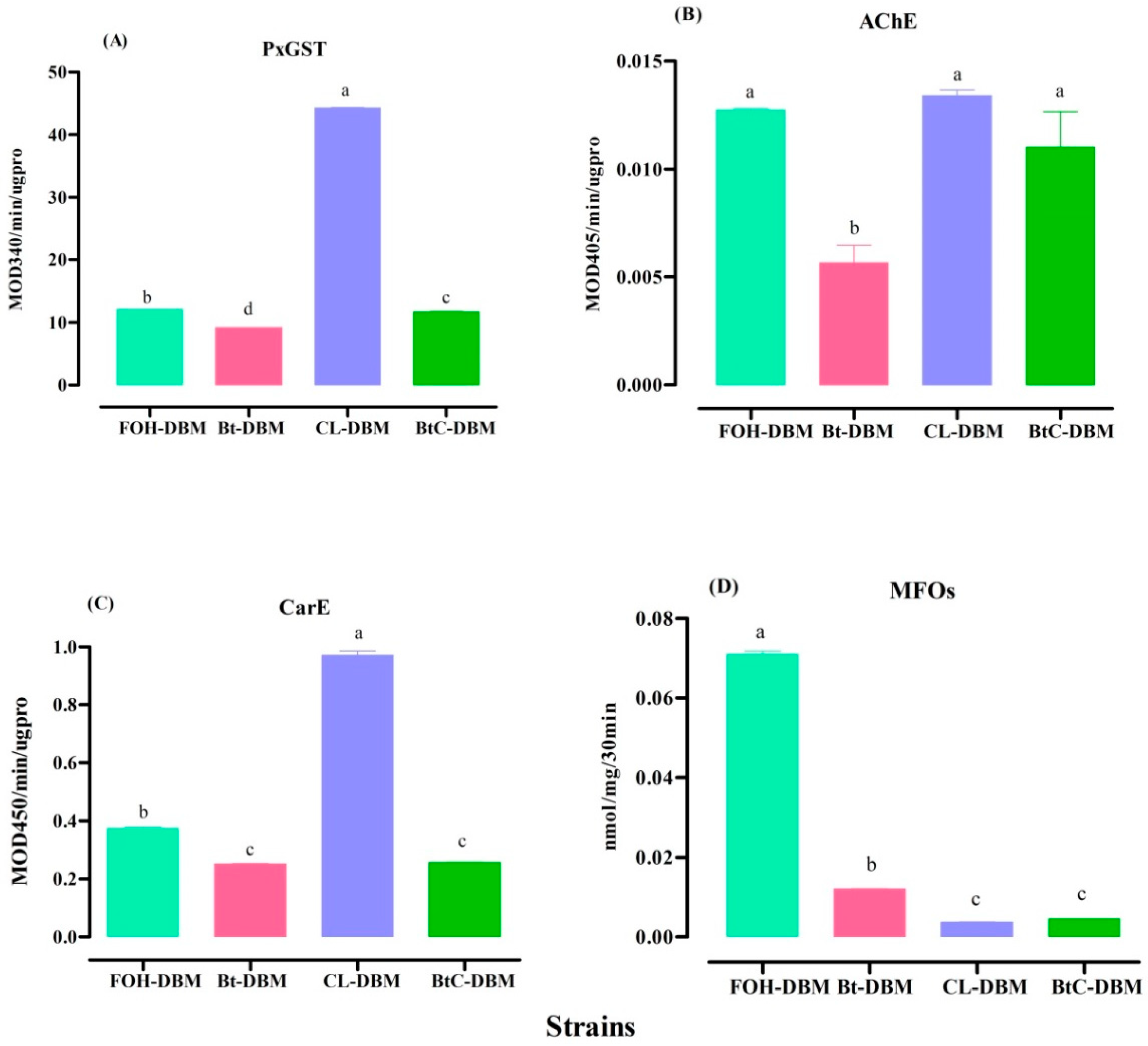
2.3. Detoxifying Enzyme Activities
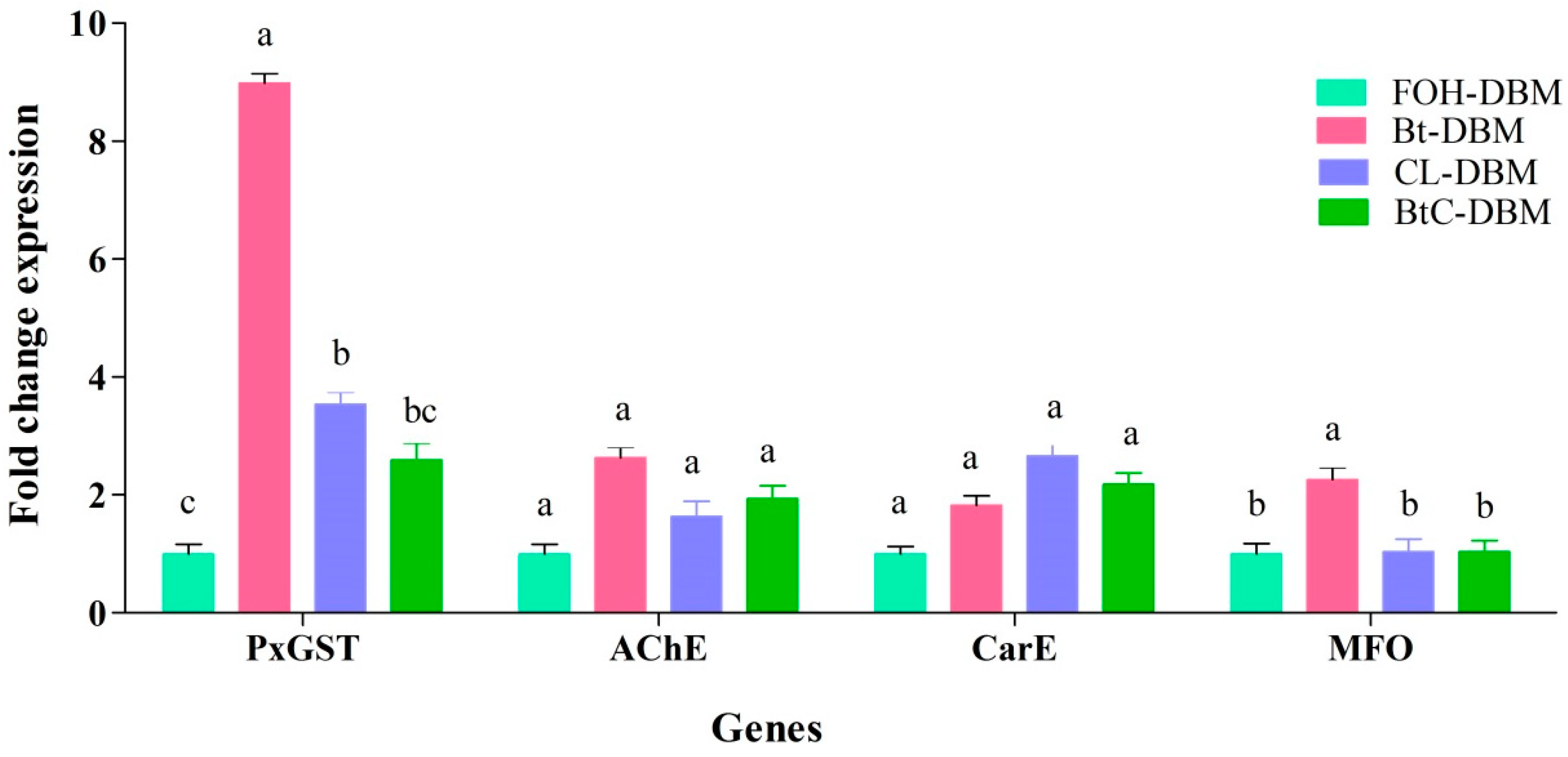
2.4. Expression Profiles of the Detoxification Genes
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Selection of DBM Strains
5.2. Insecticides and Chemical Reagents
5.3. Leaf Bioassay
5.4. The Joint Action of Insecticides
5.5. Enzyme Activity Assay
5.6. Validation of Gene Expression
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Furlong, M.J.; Wright, D.J.; Dosdall, L.M. Diamondback Moth Ecology and Management: Problems, Progress, and Prospects. Annu. Rev. Èntomol. 2013, 58, 517–541. [Google Scholar] [CrossRef]
- Jaleel, W.; Saeed, S.; Naqqash, M.N.; Sial, M.U.; Ali, M.; Zaka, S.M.; Sarwar, Z.M.; Ishtiaq, M.; Qayyum, M.A.; Aine, Q.U.; et al. Effects of temperature on baseline susceptibility and stability of insecticide resistance against Plutella xylostella (Lepidoptera: Plutellidae) in the absence of selection pressure. Saudi J. Biol. Sci. 2020, 27, 1–5. [Google Scholar] [CrossRef]
- Jaleel, W.; Saeed, S.; Saeed, Q.; Naqqash, M.N.; Sial, M.U.; Aine, Q.U.; Yanyuan, L.; Rui, Z.; He, Y.; Lu, L. Effects of three different cultivars of cruciferous plants on the age-stage, two-sex life table traits of Plutella xylostella (L.)(Lepidoptera: Plu-tellidae). Entomol. Res. 2019, 49, 151–157. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, X.; Shen, J.; Mao, K.; You, H.; Li, J. Susceptibility of field populations of the diamondback moth, Plutella xylostella, to a selection of insecticides in Central China. Pestic. Biochem. Physiol. 2016, 132, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Zalucki, M.P.; Shabbir, A.; Silva, R.; Adamson, D.; Liu, S.S.; Furlong, M.J. Estimating the Economic Cost of One of the World’s Major Insect Pests, Plutella xylostella (Lepidoptera: Plutellidae): Just How Long Is a Piece of String? J. Econ. Èntomol. 2012, 105, 1115–1129. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-Q.; Shi, Z.-H.; Zalucki, M.P.; Liu, S.-S. Conservation biological control and IPM practices in Brassica vegetable crops in China. Biol. Control. 2014, 68, 37–46. [Google Scholar] [CrossRef]
- Xia, Y.; Lu, Y.; Shen, J.; Gao, X.; Qiu, H.; Li, J. Resistance monitoring for eight insecticides in Plutella xylostella in central China. Crop. Prot. 2014, 63, 131–137. [Google Scholar] [CrossRef]
- Tabashnik, B.; Huang, F.; Ghimire, M.N.; Leonard, B.R.; Siegfried, B.D.; Rangasamy, M.; Yang, Y.; Wu, Y.; Gahan, L.J.; Heckel, D.G.; et al. Efficacy of genetically modified Bt toxins against insects with different genetic mechanisms of resistance. Nat. Biotechnol. 2011, 29, 1128–1131. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wu, Y. High Levels of Resistance to Chlorantraniliprole Evolved in Field Populations of Plutella xylostella. J. Econ. Èntomol. 2012, 105, 1019–1023. [Google Scholar] [CrossRef]
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insect resistance to neonicotinoid insecticides. Pestic. Biochem. Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Baxter, S.W.; Chen, M.; Dawson, A.; Zhao, J.-Z.; Vogel, H.; Shelton, A.M.; Heckel, D.G.; Jiggins, C.D. Mis-Spliced Transcripts of Nicotinic Acetylcholine Receptor α6 Are Associated with Field Evolved Spinosad Resistance in Plutella xylostella (L.). PLoS Genet. 2010, 6, e1000802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonoda, S. Molecular analysis of pyrethroid resistance conferred by target insensitivity and increased metabolic detoxification in Plutella xylostella. Pest Manag. Sci. 2010, 66, 572–575. [Google Scholar] [CrossRef] [PubMed]
- Bass, C.; Field, L.M. Gene amplification and insecticide resistance. Pest Manag. Sci. 2011, 67, 886–890. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular Mechanisms of Metabolic Resistance to Synthetic and Natural Xenobiotics. Annu. Rev. Èntomol. 2007, 52, 231–253. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Qu, C.; Wang, Z.; Yang, G.; Luo, C. Cross-resistance, biochemical mechanism and fitness costs of laboratory-selected resistance to pyridalyl in diamondback moth, Plutella xylostella. Pestic. Biochem. Physiol. 2020, 163, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Karunaratne, S.; Weerakoon, K. Involvement of metabolic and insensitive acetylcholinesterase mechanisms in insecticide re-sistance of rice insect pests and predatory populations from Batalagoda, Sri Lanka. J. Natn. Sci. 2007, 35, 103–108. [Google Scholar]
- Briefs, I. Global status of commercialized biotech/GM crops in 2017: Biotech crop adoption surges as economic benefits accumulate in 22 years. ISAAA Brief 2017, 53, 25–26. [Google Scholar]
- James, C. Global Review of Commercialized Transgenic Crops: 2002 Feature: Bt Maize; ISAAA Ithaca: New York, NY, USA, 2003; Volume 29. [Google Scholar]
- Shabbir, M.Z.; Quan, Y.; Wang, Z.; Bravo, A.; Soberón, M.; He, K. Characterization of the Cry1Ah resistance in Asian corn Borer and its cross-resistance to other Bacillus thuringiensis toxins. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- He, K.; Wang, Z.; Zhou, D.; Wen, L.; Song, Y.; Yao, Z. Evaluation of Transgenic Bt Corn for Resistance to the Asian Corn Borer (Lepidoptera: Pyralidae). J. Econ. Èntomol. 2003, 96, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Wang, Z.; Bravo, A.; Soberón, M.; He, K. Genetic Basis of Cry1F-Resistance in a Laboratory Selected Asian Corn Borer Strain and Its Cross-Resistance to Other Bacillus thuringiensis Toxins. PLoS ONE 2016, 11, e0161189. [Google Scholar] [CrossRef]
- Zhang, T.; He, M.; Gatehouse, A.M.R.; Wang, Z.; Edwards, M.G.; Li, Q.; He, K. Inheritance Patterns, Dominance and Cross-Resistance of Cry1Ab- and Cry1Ac-Selected Ostrinia furnacalis (Guenée). Toxins 2014, 6, 2694–2707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrière, Y.; Fabrick, J.A.; Tabashnik, B.E. Can Pyramids and Seed Mixtures Delay Resistance to Bt Crops? Trends Biotechnol. 2016, 34, 291–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, C. Global status of commercialized biotech/GM crops. ISAAA Briefs 2006, 35, 96. [Google Scholar]
- Kortenkamp, A. Ten Years of Mixing Cocktails: A Review of Combination Effects of Endocrine-Disrupting Chemicals. Environ. Health Perspect. 2007, 115, 98–105. [Google Scholar] [CrossRef] [Green Version]
- Cang, T.; Dai, D.; Yang, G.; Yu, Y.; Lv, L.; Cai, L.; Wang, Q. Combined toxicity of imidacloprid and three insecticides to the earthworm, Eisenia fetida (Annelida, Oligochaeta). Environ. Sci. Pollut. Res. 2017, 24, 8722–8730. [Google Scholar] [CrossRef] [PubMed]
- Legrand, E.; Boulangé-Lecomte, C.; Restoux, G.; Trémolet, G.; Duflot, A.; Forget-Leray, J. Individual and mixture acute toxicity of model pesticides chlordecone and pyriproxyfen in the estuarine copepod Eurytemora affinis. Environ. Sci. Pollut. Res. 2016, 24, 5976–5984. [Google Scholar] [CrossRef]
- Zhao, F.; Li, Y.; Huang, L.; Gu, Y.; Zhang, H.; Zeng, D.; Tan, H. Individual and combined toxicity of atrazine, butachlor, halosulfuron-methyl and mesotrione on the microalga Selenastrum capricornutum. Ecotoxicol. Environ. Saf. 2018, 148, 969–975. [Google Scholar] [CrossRef]
- Stemele, M. Comparative effects of a selective insecticide, Bacillus thuringiensis var. kurstaki and the broad-spectrum insecticide cypermethrin on diamondback moth and its parasitoid Cotesia vestalis (Hymenoptera; Braconidae). Crop. Prot. 2017, 101, 35–42. [Google Scholar] [CrossRef]
- Wang, Y.; Quan, Y.; Yang, J.; Shu, C.; Wang, Z.; Zhang, J.; Gatehouse, A.M.R.; Tabashnik, B.E.; He, K. Evolution of Asian Corn Borer Resistance to Bt Toxins Used Singly or in Pairs. Toxins 2019, 11, 461. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Liang, P.; Zhou, X.; Gao, X. Novel mutations and mutation combinations of ryanodine receptor in a chlorantraniliprole resistant population of Plutella xylostella (L.). Sci. Rep. 2015, 4, 6924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparks, T.C.; Nauen, R. IRAC: Mode of action classification and insecticide resistance management. Pestic. Biochem. Physiol. 2015, 121, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Maa, W.C.J.; Liao, S.C. Culture-dependent variation in esterase isozymes and malathion susceptibility of diamondback moth, Plutella xylostella L. Zool. Stud. Taipei 2000, 39, 375–386. [Google Scholar]
- Gong, Y.-J.; Wang, Z.-H.; Shi, B.-C.; Kang, Z.-J.; Zhu, L.; Jin, G.-H.; Weig, S.-J. Correlation between Pesticide Resistance and Enzyme Activity in the Diamondback Moth, Plutella xylostella. J. Insect Sci. 2013, 13, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Wu, S.; Yang, T.; Zhu, C.; Gao, C. Monitoring Field Populations ofPlutella xylostella(Lepidoptera: Plutellidae) for Resistance to Eight Insecticides in China. Fla. Èntomol. 2015, 98, 65–73. [Google Scholar] [CrossRef]
- Khaliq, A.; Attique, M.; Sayyed, A. Evidence for resistance to pyrethroids and organophosphates in Plutella xylostella (Lepidoptera: Plutellidae) from Pakistan. Bull. Èntomol. Res. 2007, 97, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.T.; Attia, F.L. Effects of Mixtures of Bacillus thuringiensis and Pesticides on Plutella xylostella and the Parasite Thyraeella collaris. J. Econ. Èntomol. 1977, 70, 146–148. [Google Scholar] [CrossRef]
- De Liguoro, M.; Riga, A.; Fariselli, P. Synergistic toxicity of some sulfonamide mixtures on Daphnia magna. Ecotoxicol. Environ. Saf. 2018, 164, 84–91. [Google Scholar] [CrossRef]
- Rizvi, S.A.H.; Ling, S.; Tian, F.; Xie, F.; Zeng, X. Toxicity and enzyme inhibition activities of the essential oil and dominant constituents derived from Artemisia absinthium L. against adult Asian citrus psyllid Diaphorina citri Kuwayama (Hemiptera: Psyllidae). Ind. Crop. Prod. 2018, 121, 468–475. [Google Scholar] [CrossRef]
- Feyereisen, R. Insect CYP Genes and P450 Enzymes. Insect Mol. Biol. Biochem. 2012, 236–316. [Google Scholar] [CrossRef]
- Oakeshott, J.G.; Claudianos, C.; Campbell, P.M.; Newcomb, R.D.; Russell, R.J. Biochemical Genetics and Genomics of Insect Esterases. Ref. Modul. Life Sci. 2019, 5. [Google Scholar] [CrossRef]
- Hu, Z.-D.; Feng, X.; Lin, Q.-S.; Chen, H.-Y.; Li, Z.-Y.; Yin, F.; Liang, P.; Gao, X.-W. Biochemical Mechanism of Chlorantraniliprole Resistance in the Diamondback Moth, Plutella xylostella Linnaeus. J. Integr. Agric. 2014, 13, 2452–2459. [Google Scholar] [CrossRef] [Green Version]
- Nehare, S.; Moharil, M.; Ghodki, B.; Lande, G.; Bisane, K.; Thakare, A.; Barkhade, U. Biochemical analysis and synergistic suppression of indoxacarb resistance in Plutella xylostella L. J. Asia-Pac. Èntomol. 2010, 13, 91–95. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Y.-L. Identification and characterisation of multiple glutathioneS-transferase genes from the diamondback moth, Plutella xylostella. Pest Manag. Sci. 2014, 71, 592–600. [Google Scholar] [CrossRef]
- Wang, N.; Qiu, X.; Ren, X.; Niu, F.; Wang, K. Resistance selection and biochemical characterization of spinosad resistance in Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Pestic. Biochem. Physiol. 2009, 95, 90–94. [Google Scholar] [CrossRef]
- Yalçın, M.; Mermer, S.; Kozacı, L.D.; Turgut, C. Insecticide resistance in two populations of Tuta absoluta (Meyrick, 1917) (Lepidoptera: Gelechiidae) from Turkey. Turk. J. Èntomol. 2015, 39, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Qian, L.; Cao, G.; Song, J.; Yin, Q.; Han, Z. Biochemical mechanisms conferring cross-resistance between tebufenozide and abamectin in Plutella xylostella. Pestic. Biochem. Physiol. 2008, 91, 175–179. [Google Scholar] [CrossRef]
- Yu, X.-Q.; Kanost, M.R. Manduca sexta lipopolysaccharide-specific immulectin-2 protects larvae from bacterial infection. Dev. Comp. Immunol. 2003, 27, 189–196. [Google Scholar] [CrossRef]
- Pavlidi, N.; Vontas, J.; Van Leeuwen, T. The role of glutathione S-transferases (GSTs) in insecticide resistance in crop pests and disease vectors. Curr. Opin. Insect Sci. 2018, 27, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.-N.; Wang, Y.-Q.; Wang, Z.-Y.; Ying-Hui, L.; Ling, Y.-H.; Zhen-Ying, W. Transcriptome differences between Cry1Ab resistant and susceptible strains of Asian corn borer. BMC Genom. 2015, 16, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Shabbir, M.Z.; Zhang, T.; Prabu, S.; Wang, Y.; Wang, Z.; Bravo, A.; Soberón, M.; He, K. Identification of Cry1Ah-binding proteins through pull down and gene expression analysis in Cry1Ah-resistant and susceptible strains of Ostrinia furnacalis. Pestic. Biochem. Physiol. 2020, 163, 200–208. [Google Scholar] [CrossRef]
- Ma, T.; Yan, H.; Shi, X.; Liu, B.; Ma, Z.; Zhang, X. Comprehensive evaluation of effective constituents in total alkaloids from Sophora alopecuroides L. and their joint action against aphids by laboratory toxicity and field efficacy. Ind. Crop. Prod. 2018, 111, 149–157. [Google Scholar] [CrossRef]
- Han, Y.; Yu, W.; Zhang, W.; Yang, Y.; Walsh, T.; Oakeshott, J.G.; Wu, Y. Variation in P450-mediated fenvalerate resistance levels is not correlated with CYP337B3 genotype in Chinese populations of Helicoverpa armigera. Pestic. Biochem. Physiol. 2015, 121, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Moores, G.D.; Denholm, I.; Devonshire, A.L. Association between Biochemical Markers and Insecticide Resistance in the Cotton Aphid, Aphis gossypiiGlover. Pestic. Biochem. Physiol. 1998, 62, 164–171. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- LeOra, S. Poloplus, a User’s Guide to Probit or Logit Analysis; LeOra Software: Berkeley, CA, USA, 2003. [Google Scholar]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Èntomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Insecticide | Strains | Slope ± SE | LC50 (95% FL) (mg/L) | df | χ2 | RR a |
|---|---|---|---|---|---|---|
| Chlorantraniliprole | FOH-DBM c | 1.075 ± 0.277 | 7.76 (1.277–16.26) | 5 | 0.744 | - |
| Bt-DBM d | 1.057 ± 0.209 | 188.08 (112.04–423.76) | 5 | 1.642 | 24.2 | |
| CL-DBM e | 0.902 ± 0.205 | 271.87 (143.82–927.94) | 5 | 1.069 | 31.2 | |
| BtC-DBM f | 0.791 ± 0.215 | 211.52 (104.06–901.522) | 5 | 0.731 | 27.3 | |
| Bt | FOH-DBM | 1.119 ± 0.175 | 0.87 (0.521–1.308) | 5 | 0.686 | - |
| Bt-DBM | 0.863 ± 0.166 | 16.29 (9.36–42.19) | 5 | 0.627 | 18.7 | |
| CL-DBM | 1.726 ± 0.117 | 1.58 (1.13–2.18) | 5 | 4.577 | 1.8 | |
| BtC-DBM | 1.592 ± 0.195 | 1.13 (0.7–1.69) | 5 | 5.097 | 1.3 | |
| Bt + Chl b | FOH-DBM | 1.061 ± 0.253 | 5.71 (1.388–11.05) | 5 | 2.251 | - |
| Bt-DBM | 1.111 ± 0.219 | 36.99 (21.51–62.92) | 5 | 0.694 | 6.5 | |
| CL-DBM | 1.022 ± 0.262 | 27.01 (16.39–42.98) | 5 | 1.403 | 4.7 | |
| BtC-DBM | 1.134 ± 0.187 | 30.25 (18.96–47.21) | 5 | 0.811 | 5.3 |
| Strains | Treatment a | Actual Mortality (%) b | Theoretical Mortality (%) c | Poison Ratio d |
|---|---|---|---|---|
| FOH-DBM | Bt + Chl | 50.00 ± 5.77 | 51.67 | 0.96 ± 0.11 b |
| Bt + (Bt + Chl) | 73.33 ± 3.33 | 53.33 | 1.35 ± 0.06 a | |
| Chl + (Bt + Chl) | 60.00 ± 3.33 | 48.33 | 1.24 ± 0.02 a, b | |
| Bt-DBM | Bt + Chl | 56.67 ± 3.33 | 56.67 | 1.00 ± 0.05 b |
| Bt + (Bt + Chl) | 76.67 ± 3.33 | 60.00 | 1.27 ± 0.10 a | |
| Chl + (Bt + Chl) | 66.67 ± 3.33 | 53.33 | 1.24 ± 0.06 a, b | |
| CL-DBM | Bt + Cl | 53.33 ± 3.33 | 53.33 | 1.00 ± 0.06 b |
| Bt + (Bt + Chl) | 73.33 ± 3.33 | 56.67 | 1.29 ± 0.10 a | |
| Chl + (Bt + Chl) | 63.33 ± 3.33 | 50.00 | 1.27 ± 0.06 a, b | |
| BtC-DBM | Bt + Chl | 56. 67 ± 3.33 | 56.67 | 1.00 ± 0.06 b |
| Bt + (Bt + Chl) | 73.33 ± 3.33 | 58. 33 | 1.25 ± 0.05 a | |
| Chl + (Bt + Chl) | 66.67 ± 3.33 | 55.00 | 1.21 ± 0.06 a, b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shabbir, M.Z.; He, L.; Shu, C.; Yin, F.; Zhang, J.; Li, Z.-Y. Assessing the Single and Combined Toxicity of Chlorantraniliprole and Bacillus thuringiensis (GO33A) against Four Selected Strains of Plutella xylostella (Lepidoptera: Plutellidae), and a Gene Expression Analysis. Toxins 2021, 13, 227. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13030227
Shabbir MZ, He L, Shu C, Yin F, Zhang J, Li Z-Y. Assessing the Single and Combined Toxicity of Chlorantraniliprole and Bacillus thuringiensis (GO33A) against Four Selected Strains of Plutella xylostella (Lepidoptera: Plutellidae), and a Gene Expression Analysis. Toxins. 2021; 13(3):227. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13030227
Chicago/Turabian StyleShabbir, Muhammad Zeeshan, Ling He, Changlong Shu, Fei Yin, Jie Zhang, and Zhen-Yu Li. 2021. "Assessing the Single and Combined Toxicity of Chlorantraniliprole and Bacillus thuringiensis (GO33A) against Four Selected Strains of Plutella xylostella (Lepidoptera: Plutellidae), and a Gene Expression Analysis" Toxins 13, no. 3: 227. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13030227