Effects of a Low Dose of T-2 Toxin on the Percentage of T and B Lymphocytes and Cytokine Secretion in the Porcine Ileal Wall

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
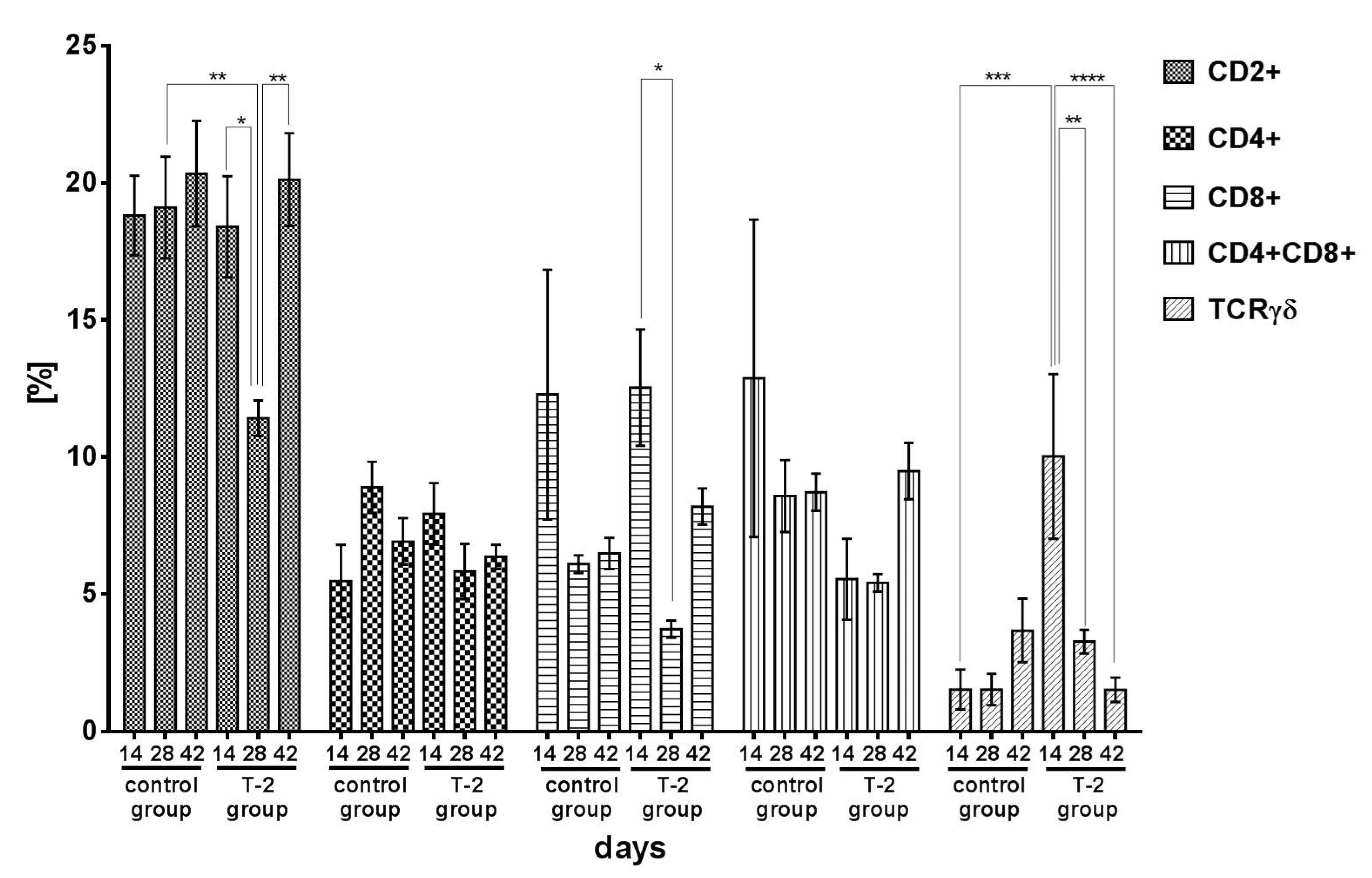
2.1. The Effect of T-2 Toxin on the Percentage of Lymphocyte Subpopulations
2.1.1. T Lymphocyte Populations
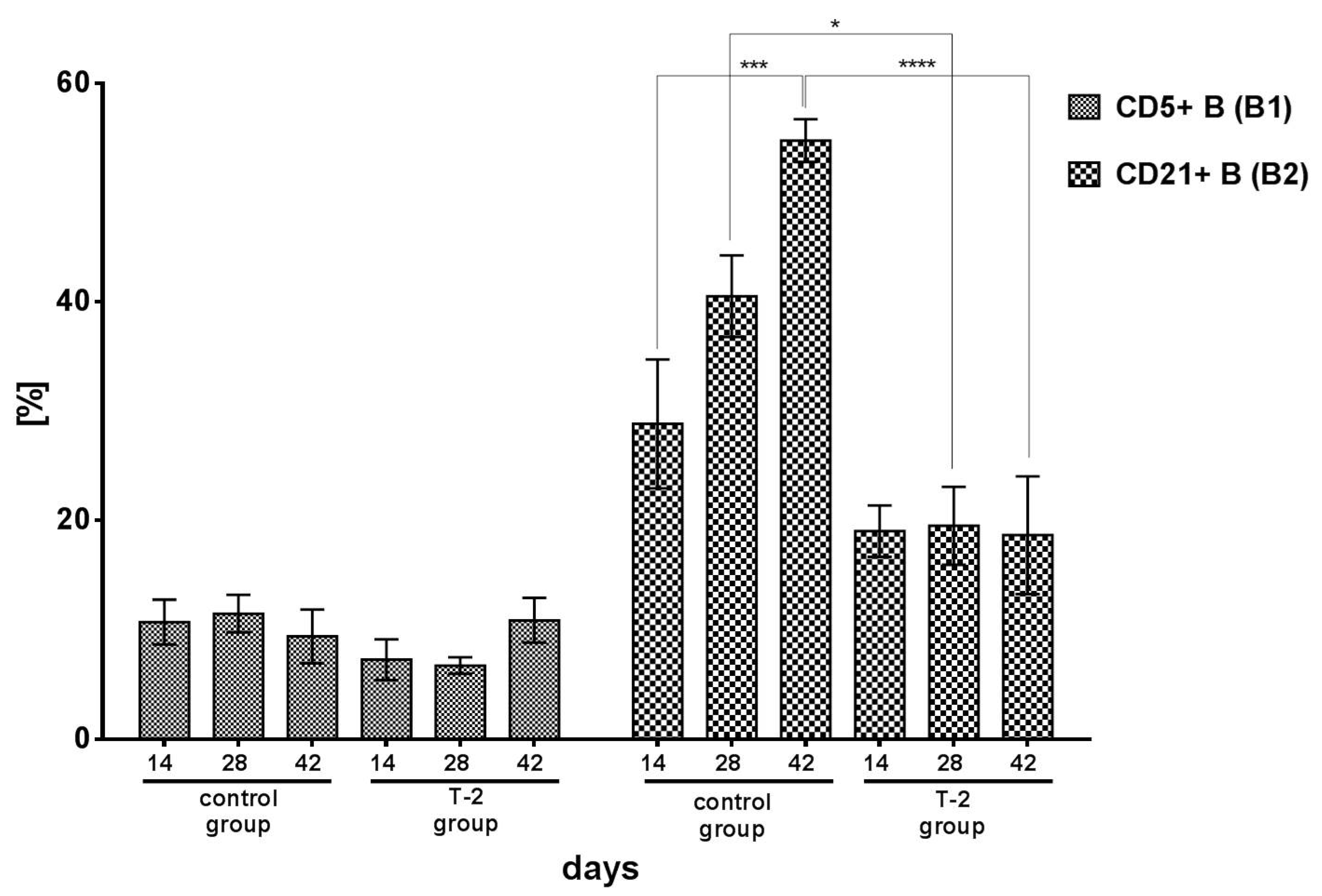
2.1.2. B Lymphocyte Populations
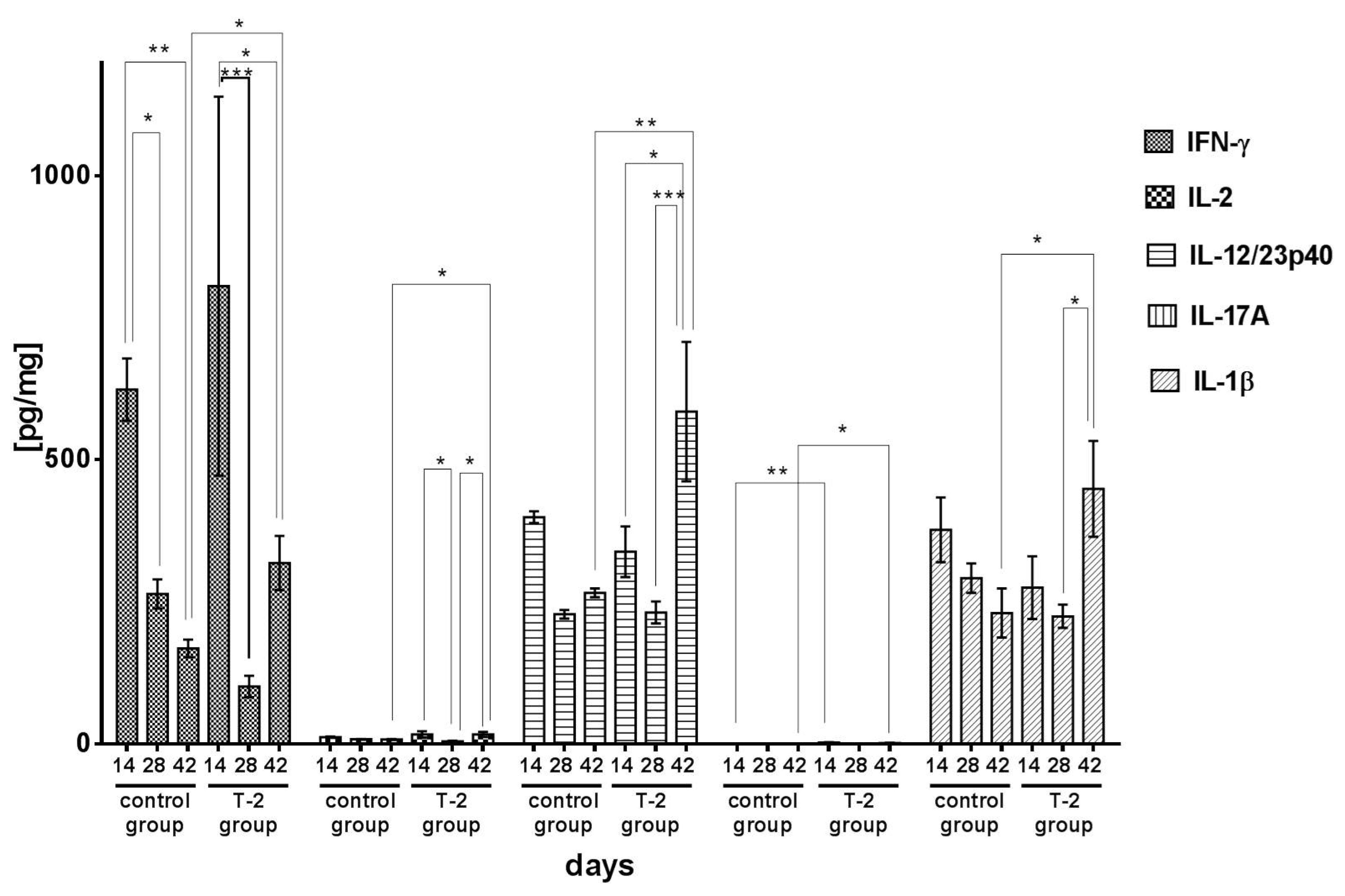
2.2. The Effect of T-2 Toxin on Cytokine Secretion
Proinflammatory Cytokines
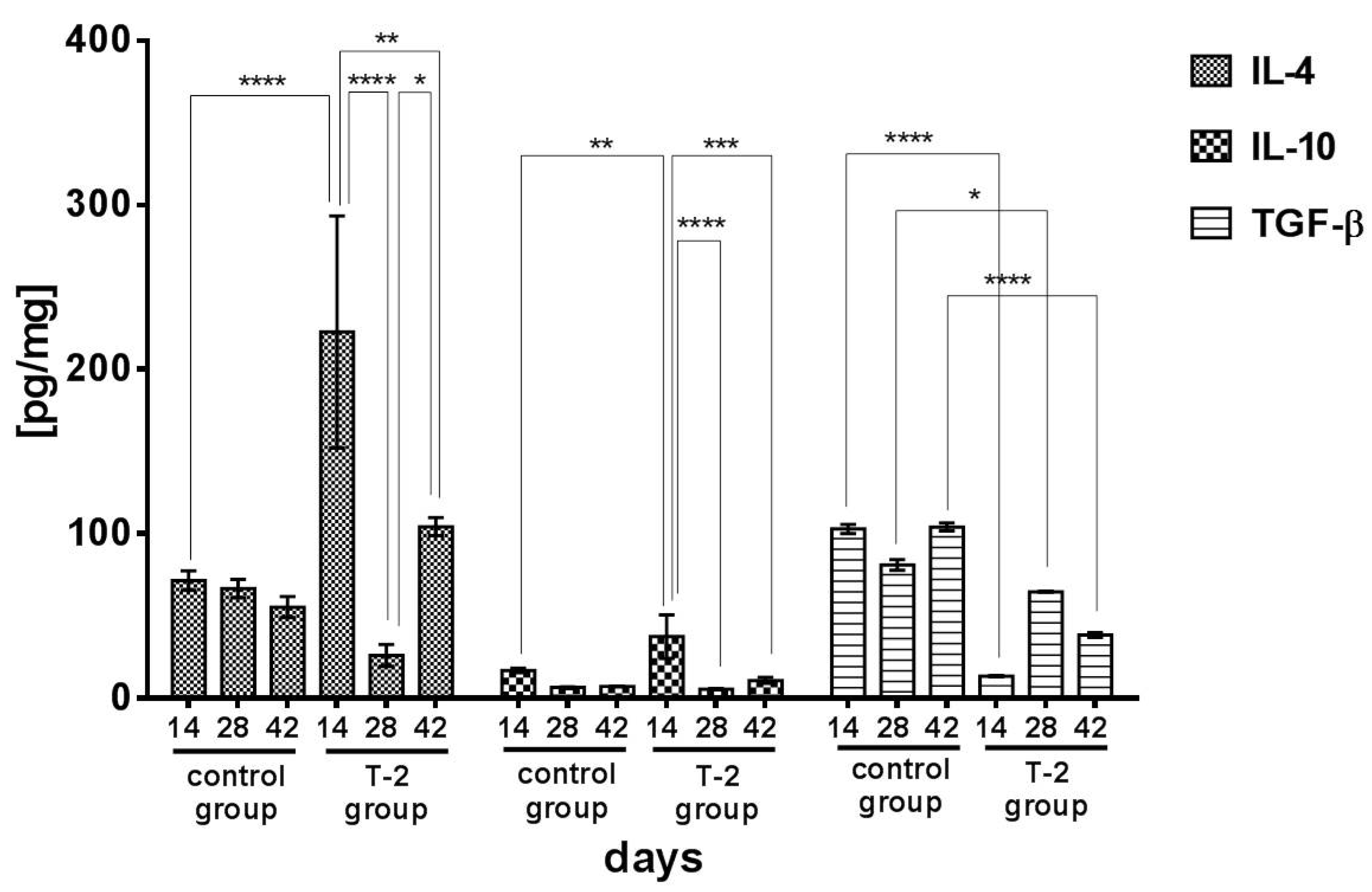
2.3. Anti-Inflammatory and Regulatory Cytokines
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals and Procedures
5.2. Tissue Sampling and Specimen Preparation
5.3. Determination of Protein Levels and Cytokine Concentrations
5.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar] [CrossRef] [Green Version]
- Calvo, A.M.; Cary, J.W. Association of fungal secondary metabolism and sclerotial biology. Front. Microbiol. 2015, 6, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinotti, L.; Ottoboni, M.; Giromini, C.; Dell’Orto, V.; Cheli, F. Mycotoxin Contamination in the EU Feed Supply Chain: A Focus on Cereal Byproducts. Toxins 2016, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Santos Pereira, C.; Cunha, C.S.; Fernandes, J.O. Prevalent Mycotoxins in Animal Feed: Occurrence and Analytical Methods. Toxins 2019, 11, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikari, M.; Negi, B.; Kaushik, N.; Adhikari, A.; Al-Khedhairy, A.A.; Kaushik, N.K.; Choi, E.H. T-2 mycotoxin: Toxicological effects and decontamination strategies. Oncotarget 2017, 8, 33933–33952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moretti, A.; Logrieco, A.F.; Susca, A. Mycotoxins: An underhand food problem. Methods Mol. Biol. 2017, 1542, 3–12. [Google Scholar]
- Zhang, J.; Zhang, H.; Liu, S.; Wu, W.; Zhang, H. Comparison of Anorectic Potencies of Type A Trichothecenes T-2 Toxin, HT-2 Toxin, Diacetoxyscirpenol, and Neosolaniol. Toxins 2018, 10, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From Simple to Complex Mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Richard, J.L. Some major mycotoxins and their mycotoxicoses—An overview. Int. J. Food Microbiol. 2007, 119, 3–10. [Google Scholar] [CrossRef]
- Kuchenbuch, H.S.; Becker, S.; Schulz, M.; Cramer, B.; Humpf, H.U. Thermal stability of T-2 and HT-2 toxins during biscuit- and crunchy muesli-making and roasting. Food Addit. Contam. Part A 2018, 35, 2158–2167. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific opinion on the risks for animal and public health related to the presence of T-2 and HT-2 toxin in food and feed. EFSA J. 2011, 9, 2481. [Google Scholar]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific opinion on the appropriateness to set a group health based guidance value for T2 and HT2 toxin and its modified forms. EFSA J. 2017, 15, 4655. [Google Scholar]
- Makowska, K.; Obremski, K.; Zielonka, L.; Gonkowski, S. The Influence of Low Doses of Zearalenone and T-2 Toxin on Calcitonin Gene Related Peptide-Like Immunoreactive (CGRP-LI) Neurons in the ENS of the Porcine Descending Colon. Toxins 2017, 9, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meissonnier, G.M.; Laffitte, J.; Raymond, I.; Benoit, E.; Cossalter, A.M.; Pinton, P.; Bertin, G.; Oswald, I.P.; Galtier, P. Subclinical doses of T-2 toxin impair acquired immune response and liver cytochrome P450 in pigs. Toxicology 2008, 247, 46–54. [Google Scholar] [CrossRef]
- Rocha, O.; Ansari, K.; Doohan, F.M. Effects of trichothecene mycotoxins on eukaryotic cells: A review. Food Addit. Contam. 2005, 22, 369–378. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol-Induced Proinflammatory Gene Expression: Mechanisms and Pathological Sequelae. Toxins 2010, 2, 1300–1317. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.H.; Wang, X.; Yang, W.; Nussler, A.K.; Xiong, L.Y.; Kuca, K.; Dohnal, V.; Zhang, X.J.; Yuan, Z.H. Oxidative stress-mediated cytotoxicity and metabolism of T-2 toxin and deoxynivalenol in animals and humans: An update. Arch. Toxicol. 2014, 88, 1309–1326. [Google Scholar] [CrossRef]
- Nagata, T.; Suzuki, H.; Ishigami, N.; Shinozuka, J.; Uetsuka, K.; Nakayama, H.; Doi, K. Development of apoptosis and changes in lymphocyte subsets in thymus, mesenteric lymph nodes and Peyer’s patches of mice orally inoculated with T-2 toxin. Exp. Toxicol. Pathol. 2001, 53, 309–315. [Google Scholar] [CrossRef]
- Grizzle, J.M.; Kersten, D.B.; McCracken, M.D.; Houston, A.E.; Saxton, A.M. Determination of the acute 50% lethal dose T-2 toxin in adult bobwhite quail: Additional studies on the effect of T-2 mycotoxin on blood chemistry and the morphology of internal organs. Avian Dis. 2004, 48, 392–399. [Google Scholar] [CrossRef]
- Makowska, K.; Gonkowski, S.; Zielonka, L.; Dabrowski, M.; Calka, J. T2 Toxin-Induced Changes in Cocaineand Amphetamine-Regulated Transcript (CART)-Like Immunoreactivity in the Enteric Nervous System Within Selected Fragments of the Porcine Digestive Tract. Neurotox. Res. 2017, 31, 136–147. [Google Scholar] [CrossRef] [Green Version]
- Weidner, M.; Huwel, S.; Ebert, F.; Schwerdtle, T.; Galla, H.J.; Humpf, H.U. Influence of T-2 and HT-2 toxin on the blood-brain barrier in vitro: New experimental hints for neurotoxic effects. PLoS ONE 2013, 8, e60484. [Google Scholar] [CrossRef]
- Li, M.; Cuff, C.F.; Pestka, J.J. T-2 toxin impairment of enteric reovirus clearance in the mouse associated with suppressed immunoglobulin and IFN-gamma responses. Toxicol. Appl. Pharmacol. 2006, 214, 318–325. [Google Scholar] [CrossRef]
- Rafai, P.; Tuboly, S.; Bata, A.; Tilly, P.; Ványi, A.; Papp, Z.; Jakab, L.; Túry, E. Effect of various levels of T-2 toxin in the immune system of growing pigs. Vet. Rec. 1995, 136, 511–514. [Google Scholar] [CrossRef]
- Hymery, N.; Sibiril, Y.; Parent-Massin, D. In vitro effects of trichothecenes on human dendritic cells. Toxicol. In Vitro 2006, 20, 899–909. [Google Scholar] [CrossRef]
- Murtaugh, M.P. Porcine cytokines. Vet. Immunol. Immunopathol. 1994, 43, 37–44. [Google Scholar] [CrossRef]
- Verma, N.; Rettenmeier, A.W.; Schmitz-Spanke, S. Recent advances in the use of Sus scrofa (pig) as a model system for proteomic studies. Proteomics 2011, 11, 776–793. [Google Scholar] [CrossRef]
- Mowat, A.M. Anatomical Basis of Tolerance and Immunity to Intestinal Antigens. Nat. Rev. Immunol. 2003, 3, 331–341. [Google Scholar] [CrossRef]
- Brandtzaeg, P.; Pabst, R. Let’s Go Mucosal: Communication on Slippery Ground. Trends. Immunol. 2004, 25, 570–577. [Google Scholar] [CrossRef]
- Nagler-Anderson, C. Man the barrier! Strategic defences in the intestinal mucosa. Nat. Rev. Immunol. 2001, 1, 59–67. [Google Scholar] [CrossRef]
- Makala, L.H.; Kamada, T.; Nagasawa, H.; Igarashi, I.; Fujisaki, K.; Suzuki, N.; Mikami, T.; Haverson, K.; Bailey, M.; Stokes, C.R.; et al. Ontogeny of pig discrete Peyer’s patches: Expression of surface antigens. J. Vet. Med. Sci. 2001, 63, 625–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, M.; Plunkett, F.J.; Rothkotter, H.J.; Vega-Lopez, M.A.; Haverson, K.; Stokes, C.R. Regulation of mucosal immune responses in effector sites. Proc. Nutr. Soc. 2001, 60, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Hayday, A.; Theodoridis, E.; Ramsburg, E.; Shires, J. Intraepithelial lymphocytes: Exploring the third way in immunology. Nat. Immunol. 2001, 2, 997–1003. [Google Scholar] [CrossRef]
- Pinton, P.; Nougayrede, J.P.; del Rio, J.C.; Moreno, C.; Marin, D.E.; Ferrier, L.; Bracarense, A.P.; Kolf-Clauw, M.; Oswald, I.P. The food contaminant deoxynivalenol, decreases intestinal barrier permeability and reduces claudin expression. Toxicol. Appl. Pharmacol. 2009, 237, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Maresca, M.; Yahi, N.; Younes-Sakr, L.; Boyron, M.; Caporiccio, B.; Fantini, J. Both direct and indirect effects account for the pro-inflammatory activity of enteropathogenic mycotoxins on the human intestinal epithelium: Stimulation of interleukin-8 secretion, potentiation of interleukin-1beta effect and increase in the transepithelial passage of commensal bacteria. Toxicol. Appl. Pharmacol. 2008, 228, 84–92. [Google Scholar]
- Maresca, M. From the gut to the brain: Journey and pathophysiological effects of the food associated trichothecene mycotoxin deoxynivalenol. Toxins 2013, 5, 784–820. [Google Scholar] [CrossRef] [PubMed]
- Onyiah, J.C.; Colgan, S.P. Cytokine responses and epithelial function in the intestinal mucosa. Cell. Mol. Life Sci. 2016, 73, 4203–4212. [Google Scholar] [CrossRef] [Green Version]
- Islam, Z.; Nagase, M.; Yoshizawa, T.; Yamauchi, K.; Sakato, N. T-2 toxin induces thymic apoptosis in vivo in mice. Toxicol. Appl. Pharmacol. 1998, 148, 205–214. [Google Scholar] [CrossRef]
- Holladay, S.D.; Smith, B.J.; Luster, M.I. B lymphocyte precursor cells represent sensitive targets of T2 mycotoxin exposure. Toxicol. Appl. Pharmacol. 1995, 131, 309–315. [Google Scholar] [CrossRef]
- Ahmadi, K.; Riazipour, M. Effects of T-2 toxin on cytokine production by mice peritoneal macrophages and lymph node T-cells. Iran. J. Immunol. 2008, 5, 177–180. [Google Scholar]
- Edite Bezerra da Rocha, M.; Freire, F.D.C.O.; Erlan, F.M.F.; Izabel, F.G.M.; Rondina, D. Mycotoxins and their effects on human and animal health. Food Control. 2014, 36, 159–165. [Google Scholar] [CrossRef]
- Murtaugh, M.P.; Johnson, C.R.; Xiao, Z.; Scamurra, R.W.; Zhou, Y. Species specialization in cytokine biology: Is interleukin-4 central to the T(H)1-T(H)2 paradigm in swine? Dev. Comp. Immunol. 2009, 33, 344–352. [Google Scholar] [CrossRef]
- Helm, R.M.; Furuta, G.T.; Stanley, J.S.; Ye, J.; Cockrell, G.; Connaughton, C.; Simpson, P.; Bannon, G.A.; Burks, A.W. A neonatal swine model for peanut allergy. J. Allergy Clin. Immunol. 2002, 109, 136–142. [Google Scholar] [CrossRef]
- Chung, Y.J.; Zhou, H.R.; Pestka, J.J. Transcriptional and posttranscriptional roles for p38 mitogen-activated protein kinase in upregulation of TNF-alpha expression by deoxynivalenol (vomitoxin). Toxicol. Appl. Pharmacol. 2003, 193, 188–201. [Google Scholar] [CrossRef]
- Meky, F.A.; Hardie, L.J.; Evans, S.W.; Wild, C.P. Deoxynivalenol-induced immunomodulation of human lymphocyte proliferation and cytokine production. Food Chem. Toxicol. 2001, 39, 827–836. [Google Scholar] [CrossRef]
- Taranu, I.; Marin, D.E.; Bouhet, S.; Pascale, F.; Bailly, J.D.; Miller, J.D.; Pinton, P.; Oswald, I.P. Mycotoxin fumonisin B1 alters the cytokine profile and decreases the vaccinal antibody titer in pigs. Toxicol. Sci. 2005, 84, 301–307. [Google Scholar] [CrossRef] [Green Version]
- McGeachy, M.J.; Bak-Jensen, K.S.; Chen, Y.; Tato, C.M.; Blumenschein, W.; McClanahan, T.; Cua, D.J. TGF-beta and IL-6 drive the production of IL-17 and IL-10 by T cells and restrain TH-17 cell-mediated pathology. Nat. Immunol. 2007, 8, 1390–1397. [Google Scholar] [CrossRef]
- Kuwabara, T.; Ishikawa, F.; Kondo, M.; Kakiuchi, T. The Role of IL-17 and Related Cytokines in Inflammatory Autoimmune Diseases. Mediat. Inflamm. 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- Schuhmacher-Wolz, U.; Heine, K.; Schneider, K. Report on toxicity data on trichothecene mycotoxins HT-2 and T-2 toxins. EFSA Support. Publ. 2010, 7. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef]
- Li, S.J.; Pasmans, F.; Croubels, S.; Verbrugghe, E.; Van Waeyenberghe, L.; Yang, Z.; Haesebrouck, F.; Martel, A. T-2 toxin impairs antifungal activities of chicken macrophages against Aspergillus fumigatus conidia but promotes the pro-inflammatory responses. Avian Pathol. 2013, 42, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Chistiakov, D.A.; Bobryshev, Y.V.; Kozarov, E.; Sobenin, I.A.; Orekhov, A.N. Intestinal mucosal tolerance and impact of gut microbiota to mucosal tolerance. Front. Microbiol. 2015, 5, 781. [Google Scholar] [CrossRef]
- Pabst, O.; Mowat, A.M. Oral tolerance to food protein. Mucosal Immunol. 2012, 5, 232–239. [Google Scholar] [CrossRef]
- Sun, C.M.; Hall, J.A.; Blank, R.B.; Bouladoux, N.; Oukka, M.; Mora, J.R.; Belkaid, Y. Small intestine lamina propria dendritic cells promote de novo generation of Foxp3 T reg cells via retinoic acid. J. Exp. Med. 2007, 204, 1775–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iliev, I.D.; Spadoni, I.; Mileti, E.; Matteoli, G.; Sonzogni, A.; Sampietro, G.M.; Foschi, D.; Caprioli, F.; Viale, G.; Rescigno, M. Human intestinal epithelial cells promote the differentiation of tolerogenic dendritic cells. Gut 2009, 58, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Spadoni, I.; Iliev, I.D.; Rossi, G.; Rescigno, M. Dendritic cells produce TSLP that limits the differentiation of Th17 cells, fosters Treg development, and protects against colitis. Mucosal Immunol. 2012, 5, 184–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shevach, E.M. Mechanisms of foxp3+ T regulatory cell-mediated suppression. Immunity 2009, 30, 636–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadis, U.; Wahl, B.; Schulz, O.; Hardtke-Wolenski, M.; Schippers, A.; Wagner, N.; Müller, W.; Sparwasser, T.; Förster, R.; Pabst, O. Intestinal tolerance requires gut homing and expansion of FoxP3+ regulatory T cells in the lamina propria. Immunity 2011, 34, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Kankkunen, P.; Rintahaka, J.; Aalto, A.; Leino, M.; Majuri, M.L.; Alenius, H.; Wolff, H.; Matikainen, S. Trichothecene mycotoxins activate inflammatory response in human macrophages. J. Immunol. 2009, 182, 6418–6425. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Pestka, J.J. Comparative Induction of 28S Ribosomal RNA Cleavage by Ricin and the Trichothecenes Deoxynivalenol and T-2 Toxin in the Macrophage. Toxicol. Sci. 2008, 105, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Sergent, T.; Parys, M.; Garsou, S.; Pussemier, L.; Schneider, Y.J.; Larondelle, Y. Deoxynivalenol transport across human intestinal Caco-2 cells and its effects on cellular metabolism at realistic intestinal concentrations. Toxicol. Lett. 2006, 164, 167–176. [Google Scholar] [CrossRef]
- Leyva-Illades, D.; Cherla, R.P.; Lee, M.S.; Tesh, V.L. Regulation of cytokine and chemokine expression by the ribotoxic stress response elicited by Shiga toxin type 1 in human macrophage-like THP-1 cells. Infect. Immun. 2012, 80, 2109–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avantaggiato, G.; Havenaar, R.; Visconti, A. Evaluation of the intestinal absorption of deoxynivalenol and nivalenol by an in vitro gastrointestinal model, and the binding efficacy of activated carbon and other adsorbent materials. Food Chem. Toxicol. 2004, 42, 817–824. [Google Scholar] [CrossRef]
- Behrens, M.; Hüwel, S.; Galla, H.J.; Humpf, H.U. Blood-Brain Barrier Effects of the Fusarium Mycotoxins Deoxynivalenol, 3 Acetyldeoxynivalenol, and Moniliformin and Their Transfer to the Brain. PLoS ONE 2015, 10, e0143640. [Google Scholar] [CrossRef]
- Lucioli, J.; Pinton, P.; Callu, P.; Laffitte, J.; Grosjean, F.; Kolf-Clauw, M.; Oswald, I.P.; Bracarense, A.P. The food contaminant deoxynivalenol activates the mitogen activated protein kinases in the intestine: Interest of ex vivo models as an alternative to in vivo experiments. Toxicon 2013, 66, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Abraham, C.; Cho, J.H. IL-23 and autoimmunity: New insights into the pathogenesis of inflammatory bowel disease. Annu. Rev. Med. 2009, 60, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Hamada, H.; Hiroi, T.; Nishiyama, Y.; Takahashi, H.; Masunaga, Y.; Hachimura, S.; Kaminogawa, S.; Takahashi-Iwanaga, H.; Iwanaga, T.; Kiyono, H.; et al. Identification of multiple isolated lymphoid follicles on the antimesenteric wall of the mouse small intestine. J. Immunol. 2002, 168, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowak, D.; Panaszek, B. Anaphylactic reactions to low-molecular weight chemicals. Postepy Hig. I Med. Dosw. (Online) 2015, 69, 197–206. [Google Scholar] [PubMed]
- Ngampongsa, S.; Hanafusa, M.; Ando, K.; Ito, K.; Kuwahara, M.; Yamamoto, Y.; Yamashita, M.; Tsuru, Y.; Tsubone, H. Toxic effects of T-2 toxin and deoxynivalenol on the mitochondrial electron transport system of cardiomyocytes in rats. J. Toxicol. Sci. 2013, 38, 495–502. [Google Scholar] [CrossRef] [Green Version]
- Tannahill, G.M.; Curtis, A.M.; Adamik, J.; Palsson-McDermott, E.M.; McGettrick, A.F.; Goel, G.; Frezza, C.; Bernard, N.J.; Kelly, B.; Foley, N.H.; et al. Succinate is an inflammatory signal that induces IL-1β through HIF-1α. Nature 2013, 496, 238–242. [Google Scholar] [CrossRef]
- Mills, E.L.; O’Neill, L.A. Reprogramming mitochondrial metabolism in macrophages as an anti-inflammatory signal. Eur. J. Immunol. 2016, 46, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Unit | Diet 1 | Diet 2 |
|---|---|---|---|
| Metabolizable energy | MJ | 13.80 | 13.00 |
| Total protein | % | 6.00 | 16.00 |
| Crude fiber | max% | 2.63 | 4.94 |
| Sodium | % | 0.21 | 0,18 |
| Calcium | min% | 0.73 | 0.58 |
| Total phosphorus | min% | 0.65 | 0.73 |
| Lysine | % | 1.27 | 1.04 |
| Methionine + cystine | % | 0.83 | 0.64 |
| Threonine | % | 0.72 | 0.59 |
| Tryptophan | % | 0.26 | 0.20 |
| Vitamin A | IU | 15,000 | 13,500 |
| Vitamin D3 | IU | 2000 | 2000 |
| Vitamin E | IU | 100 | 60 |
| Phytase | present | + | + |
| Enzymes | present | + | + |
| Flavor | present | + | - |
| Acidifier | present | + | + |
| Antigen | Subpopulation |
|---|---|
| CD2+ | T cells |
| CD4+ | Th cells |
| CD8+ | Cytotoxic T cells |
| CD4+CD8+ | Double-positive T cells |
| CD5-TCR+ | TCRγδ+ cells, intraepithelial lymphocytes |
| CD5+CD8- | B1 cells |
| CD21+ | B2 cells |
| Symbol | Detected Antigen | Immunoglobulin Class | Catalog Number |
|---|---|---|---|
| P1 | CD2 | IgG2a | MSA4 |
| P2 | CD4 | IgG2b | 74-12-4 |
| P3 | CD5 | IgG1 | PG114A |
| P4 | CD8α | IgG2a | 76-2-11 |
| P5 | CD21 | IgG1 | BB6-11C9 |
| P6 | TCR1-N7 | IgG1 | 86D |
| Symbol | Immunoglobulin or Ligand Class | Dye | Catalog Number |
|---|---|---|---|
| S1 | Mouse IgG1 | Phycoerythrin (PE) | 550083 |
| S2 | Mouse IgG2a | Fluorescein isothiocyanate (FITC) | 553390 |
| S3 | Mouse IgG2b | Biotin | 550333 |
| S-PE | Biotin | PE | 554061 |
| Antigen | ELISA Kit and Catalog Number | Manufacturer, Country | Assay Range pg/mL |
|---|---|---|---|
| IFN-γ | Porcine IFN-gamma DuoSet ELISA, DY985 | R&D Systems Inc., Minneapolis, MN, USA | 62.5–4.000 Intra-assay CV < 3.4% Inter-assay CV < 4.6% |
| IL-1β | Porcine IL-1 beta/IL-1F2 DuoSet ELISA, DY681 | R&D Systems Inc., Minneapolis, MN, USA | 62.5–4.000 Intra-assay CV < 1.1% Inter-assay CV < 3.2% |
| IL-2 | Swine IL-2 CytoSet™ CSC124 | Invitrogens™, Camarillo, CA, USA | 35.6–570 Intra-assay CV < 4.48% Inter-assay CV < 5.02% |
| IL-12/23p40 | Porcine IL-12/IL-23 p40 DuoSet ELISA, DY912 | R&D Systems Inc., Minneapolis, MN, USA | 78.1–5.000 Intra-assay CV < 3.67% Inter-assay CV < 4.25% |
| IL-4 | Porcine IL-4 DuoSet ELISA, DY654 | R&D Systems Inc., Minneapolis, MN, USA | 156.0–10.000 Intra-assay CV < 5% Inter-assay CV < 6.69% |
| IL-10 | Porcine IL-10 DuoSet ELISA, DY693B | R&D Systems Inc., Minneapolis, MN, USA | 23.4–1.500 Intra-assay CV <3% Inter-assay CV <4.64% |
| TGFβ | TGF beta-1 Multispecies Matched Antibody Pair, CHC1683 | Thermo Fisher Scientific, Waltham, MA, USA | 62.5–4.000 Intra-assay CV < 2.9% Inter-assay CV < 5% |
| IL-17A | Porcine IL-17 (IL-17A) ELISA Kit, ESIL17A | Thermo Fisher Scientific, Waltham, MA, USA | 16.38–4.000 Intra-assay CV < 10% Inter-assay CV < 12% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojtacha, P.; Trybowski, W.; Podlasz, P.; Żmigrodzka, M.; Tyburski, J.; Polak-Śliwińska, M.; Jakimiuk, E.; Bakuła, T.; Baranowski, M.; Żuk-Gołaszewska, K.; et al. Effects of a Low Dose of T-2 Toxin on the Percentage of T and B Lymphocytes and Cytokine Secretion in the Porcine Ileal Wall. Toxins 2021, 13, 277. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13040277
Wojtacha P, Trybowski W, Podlasz P, Żmigrodzka M, Tyburski J, Polak-Śliwińska M, Jakimiuk E, Bakuła T, Baranowski M, Żuk-Gołaszewska K, et al. Effects of a Low Dose of T-2 Toxin on the Percentage of T and B Lymphocytes and Cytokine Secretion in the Porcine Ileal Wall. Toxins. 2021; 13(4):277. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13040277
Chicago/Turabian StyleWojtacha, Paweł, Wojciech Trybowski, Piotr Podlasz, Magdalena Żmigrodzka, Józef Tyburski, Magdalena Polak-Śliwińska, Ewa Jakimiuk, Tadeusz Bakuła, Mirosław Baranowski, Krystyna Żuk-Gołaszewska, and et al. 2021. "Effects of a Low Dose of T-2 Toxin on the Percentage of T and B Lymphocytes and Cytokine Secretion in the Porcine Ileal Wall" Toxins 13, no. 4: 277. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13040277