The Rapid Evolution of Resistance to Vip3Aa Insecticidal Protein in Mythimna separata (Walker) Is Not Related to Altered Binding to Midgut Receptors


 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Response to Selection with Vip3Aa and Cry1 Toxins in M. separata
2.2. Cross-Resistance Evaluation of Vip3Aa-Resistant M. separata Strain to Cry1 Proteins
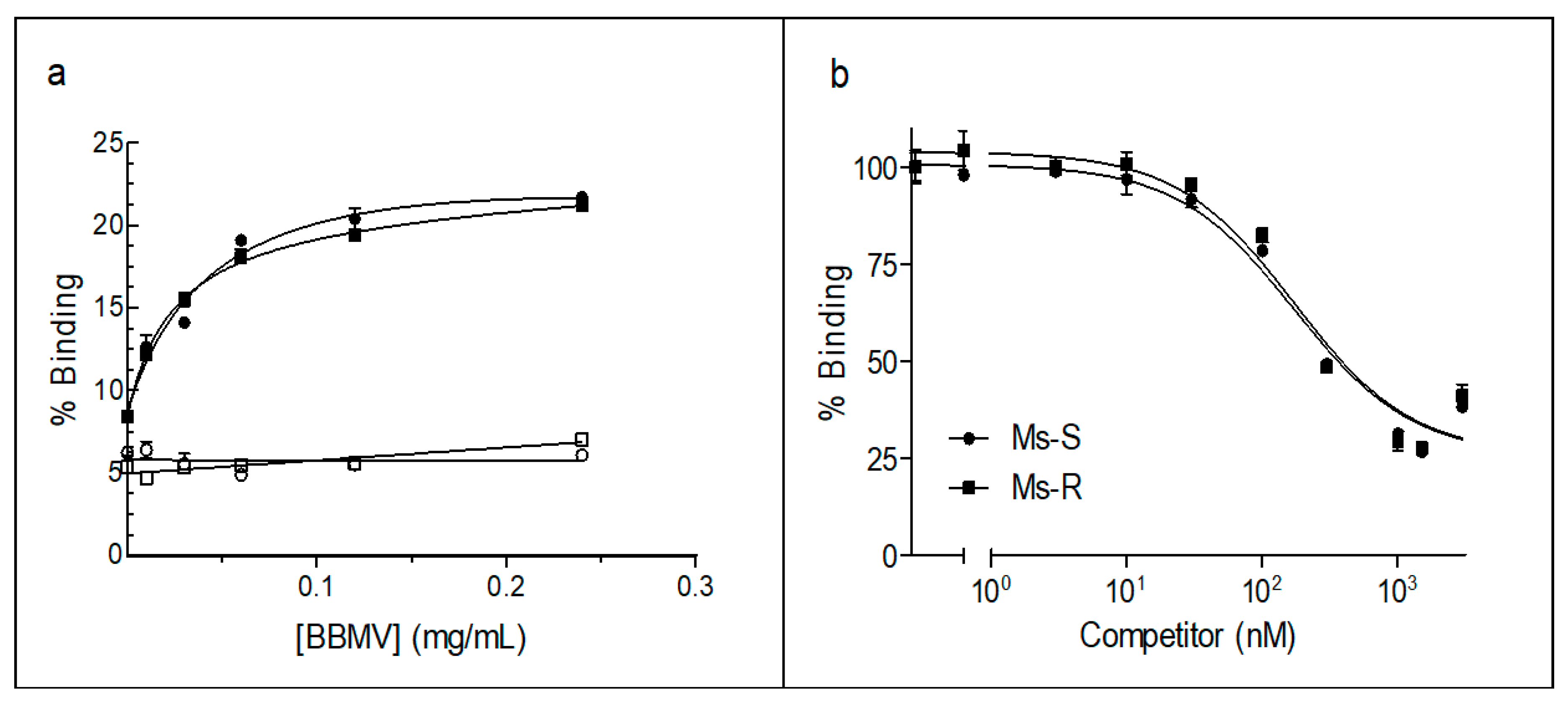
2.3. 125I-Vip3Aa Binding to BBMV of Susceptible and Resistant M. separata
3. Discussion
4. Materials and Methods
4.1. Insect Colonies
4.2. Source of Toxins
4.3. Bioassays and Selection Process
4.4. Protein Purification for Binding Analysis
4.5. Vip3Aa Radiolabeling
4.6. BBMV Preparation
4.7. Binding Assays with 125I-labeled Vip3Aa
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jiang, X.F.; Luo, L.Z.; Zhang, L.; Sappington, T.W.; Hu, Y. Regulation of migration in Mythimna separata (Walker) in China: A review integrating environmental, physiological, hormonal, genetic, and molecular factors. Environ. Entomol. 2011, 40, 516–533. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.; Khan, R.A.; Zhang, Y.L. Physiological and population responses of armyworm Mythimna separata (Lepidoptera: Noctuidae) to a sublethal dose of cantharidin-AC. J. Econ. Entomol. 2013, 106, 2177–2182. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Jiang, Y.Y.; Liu, J. Analysis of the armyworm outbreak in 2012 and suggestions of monitoring and forecasting. Plant Prot. 2013, 39, 117–121. (In Chinese) [Google Scholar]
- Zhang, Z.; Zhang, Y.; Wang, J.; Liu, J.; Tang, Q.; Li, X.; Cheng, D. Insecticide resistance of the field populations of oriental armyworm, Mythimna separata (Walker) in Shaanxi and Shanxi provinces of China. J. Integr. Agric. 2018, 17, 1527–1537. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Jiang, T.; Sun, Y.; Gu, H.; Wu, J.; Yang, Y. Effect of three insect-resistant maizes expressing Cry1Ie, Cry1Ab/Cry2Aj and Cry1Ab on the growth and development of armyworm Mythimna separata (Walker). J. Integr. Agric. 2020, 19, 1842–18491. [Google Scholar] [CrossRef]
- James, C. Global status of commercialization of biotech/GM crops in 2017: Biotech crop adoption surges as economic benefits accumulate in 22 years. In ISAAA Brief No. 53; ISAAA: Ithaca, NY, USA, 2017; pp. 1–4. [Google Scholar]
- Ferré, J.; Van Rie, J. Biochemistry and Genetics of Insect Resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 2002, 47, 501–533. [Google Scholar] [CrossRef] [PubMed]
- Jurat-Fuentes, J.L.; Heckel, D.G.; Ferré, J. Mechanism of resistance to insecticidal proteins from Bacillus thuringiensis. Annu. Rev. Entomol. 2021, 66, 121–140. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Brévault, T.; Carrière, Y. Insect resistance to Bt crops: Lessons from the first billion acres. Nat. Biotechnol. 2013, 31, 510–521. [Google Scholar] [CrossRef]
- Chakroun, M.; Banyuls, N.; Bel, Y.; Escriche, B.; Ferré, J. Bacterial vegetative insecticidal proteins (Vip) from entomopathogenic bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 329–350. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarty, S.; Jin, M.; Wu, C.; Chakraborty, P.; Xiao, Y. Bacillus thuringiensis vegetative insecticidal protein family Vip3A and mode of action against pest Lepidoptera. Pest. Manag. Sci. 2020, 76, 1612–1617. [Google Scholar] [CrossRef]
- Carrière, Y.; Crickmore, N.; Tabashnik, B.E. Optimizing pyramided transgenic Bt crops for sustainable pest management. Nat. Biotechnol. 2015, 33, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Mahon, R.J.; Downes, S.J.; James, B. Vip3A resistance alleles exist at high levels in Australian targets before release of cotton expressing this toxin. PLoS ONE 2012, 7, e39192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Santiago-González, J.C.; Little, N.; Reisig, D.; Payne, G.; Ferreira Dos Santos, R.; Jurat-Fuentes, J.L.; Kurtz, R.; Kerns, D.L. First documentation of major Vip3Aa resistance alleles in field populations of Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae) in Texas, USA. Sci. Rep. 2020, 10, 5867. [Google Scholar] [CrossRef] [Green Version]
- Pickett, B.R.; Gulzar, A.; Ferré, J.; Wright, D.J. Bacillus thuringiensis Vip3Aa toxin resistance in Heliothis virescens (Lepidoptera: Noctuidae). Appl. Environ. Microbiol. 2017, 83, e03506-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkhade, U.P.; Thakare, A.S. Protease mediated resistance mechanism to Cry1C and Vip3A in Spodoptera litura. Egypt. Acad. J. Biol. Sci. 2010, 3, 43–50. [Google Scholar] [CrossRef]
- Bernardi, O.; Bernardi, D.; Horikoshi, R.J.; Okuma, D.M.; Miraldo, L.L.; Fatoretto, J.; Medeiros, F.C.; Burd, T.; Omoto, C. Selection and characterization of resistance to the Vip3Aa20 protein from Bacillus thuringiensis in Spodoptera frugiperda. Pest. Manag. Sci. 2016, 72, 1794–1802. [Google Scholar] [CrossRef]
- Chang, X.; Chang, X.Y.; He, K.L.; Wang, Z.Y.; Bai, S.X. Resistance evaluation of transgenic Bt maize to oriental armyworm. Acta Phytophyl. Sin. 2007, 34, 225–228. [Google Scholar]
- He, K.L.; Wang, Z.Y.; Zhou, D.R.; Wen, L.P.; Song, Y.Y.; Yao, Z.Y. Evaluation of transgenic Bt corn for resistance to the Asian corn borer (Lepidoptera: Pyralidae). J. Econ. Entomol. 2003, 96, 935–940. [Google Scholar] [CrossRef]
- Buschman, L.; Sloderbeck, P.; Witt, M. Efficacy of Cry1F corn for the control of southwestern corn borer and corn earworm, 2000. Arthropod Manage. Tests 2001, 26, M2. [Google Scholar]
- Siebert, M.W.; Babcock, J.M.; Nolting, S.; Santos, A.C.; Adamczyk, J.J.; Neese, P.A.; King, J.E.; Jenkins, J.N.; McCarty, J.; Lorenz, G.M.; et al. Efficacy of Cry1F insecticidal protein in maize and cotton for control of fall armyworm (Lepidoptera: Noctuidae). Fla. Entomol. 2008, 91, 555–565. [Google Scholar]
- Xu, L.; Wang, Z.; Zhang, J.; He, K.; Ferry, N.; Gatehouse, A.M.R. Cross-resistance of Cry1Ab-selected Asian corn borer to other Cry toxins. J. Appl. Entomol. 2010, 134, 429–438. [Google Scholar] [CrossRef]
- Pereira, E.J.G.; Storer, N.P.; Siegfried, B.D. Fitness costs of Cry1F resistance in laboratory-selected European corn borer (Lepidoptera: Crambidae). J. Appl. Entomol. 2011, 135, 17–24. [Google Scholar] [CrossRef]
- Chakroun, M.; Ferré, J. In vivo and in vitro binding of Vip3Aa to Spodoptera frugiperda midgut and characterization of binding sites by 125I radiolabeling. Appl. Environ. Microbiol. 2014, 80, 6258–6265. [Google Scholar] [CrossRef] [Green Version]
- Kahn, T.W.; Chakroun, M.; Williams, J.; Walsh, T.; James, B.; Monserrate, J.; Ferré, J. Efficacy and resistance management potential of a modified Vip3C protein for control of Spodoptera frugiperda in maize. Sci. Rep. 2018, 8, 16204. [Google Scholar] [CrossRef] [PubMed]
- Sena, J.A.D.; Hernández-Rodríguez, C.S.; Ferré, J. Interaction of Bacillus thuringiensis Cry1 and Vip3Aa proteins with Spodoptera frugiperda midgut binding sites. Appl. Environ. Microbiol. 2009, 75, 2236–2237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouffon, C.; Van Vliet, A.; Van Rie, J.; Jansens, S.; Jurat-Fuentes, J.L. Binding sites for Bacillus thuringiensis Cry2Ae toxin on heliothine brush border membrane vesicles are not shared with Cry1A, Cry1F, or Vip3A toxin. Appl. Environ. Microbiol. 2011, 77, 3182–3188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, G.; Sachdev, B.; Sharma, N.; Seth, R.; Bhatnagar, R.K. Interaction of Bacillus thuringiensis vegetative insecticidal protein with Ribosomal S2 protein triggers larvicidal activity in Spodoptera frugiperda. Appl. Environ. Microbiol. 2010, 76, 7202–7209. [Google Scholar] [CrossRef] [Green Version]
- Jiang, K.; Hou, X.; Tan, T.; Cao, Z.; Mei, S.; Yan, B.; Chang, J.; Han, L.; Zhao, D.; Cai, J. Scavenger receptor-C acts as a receptor for Bacillus thuringiensis vegetative insecticidal protein Vip3Aa and mediates the internalization of Vip3Aa via endocytosis. PLoS Pathog. 2018, 14, e1007347. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Hou, X.; Han, L.; Tan, T.; Cao, Z.; Cai, J. Fibroblast growth factor receptor, a novel receptor for vegetative insecticidal protein Vip3Aa. Toxins 2018, 10, 546. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.K.; Walters, F.S.; Hart, H.; Palekar, N.; Chen, J.S. Mode of action of the Bacillus thuringiensis vegetative insecticidal protein Vip3A differs from that of Cry1Ab δ-endotoxin. Appl. Environ. Microbiol. 2003, 69, 4648–4657. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.K.; Miles, P.; Chen, J.S. Brush border membrane binding properties of Bacillus thuringiensis Vip3A toxin to Heliothis virescens and Helicoverpa zea midguts. Biochem. Biophys. Res. Commun. 2006, 339, 1043–1047. [Google Scholar] [CrossRef] [PubMed]
- Chakroun, M.; Banyuls, N.; Walsh, T.; Downes, S.; James, B.; Ferré, J. Characterization of the resistance to Vip3Aa in Helicoverpa armigera from Australia and the role of midgut processing and receptor binding. Sci. Rep. 2016, 6, 24311. [Google Scholar] [CrossRef]
- Pinos, D.; Chakroun, M.; Millán-Leiva, A.; Jurat-Fuentes, J.L.; Wright, D.J.; Hernández-Martínez, P.; Ferré, J. Reduced membrane-bound alkaline phosphatase does not affect binding of Vip3Aa in a Heliothis virescens resistant colony. Toxins 2020, 12, 409. [Google Scholar] [CrossRef] [PubMed]
- Pinos, D.; Andrés-Garrido, A.; Ferré, J.; Hernández-Martínez, P. Response mechanisms of invertebrates to Bacillus thuringiensis and its pesticidal proteins. Microbiol. Mol. Biol. Rev. 2021, 85, e00007-20. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Quan, Y.; Sivaprasath, P.; Shabbir, M.S.; Wang, Z.; Ferré, J.; He, K. Insecticidal activity and synergistic combinations of ten different Bt toxins against Mythimna separata (Walker). Toxins 2018, 10, 454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemes, A.R.N.; Davolos, C.C.; Legori, P.C.B.C.; Fernandes, O.A.; Ferré, J.; Lemos, M.V.F.; Desiderio, J.A. Synergism and Antagonism between Bacillus thuringiensis Vip3A and Cry1 Proteins in Heliothis virescens, Diatraea saccharalis and Spodoptera frugiperda. PLoS ONE 2014, 9, e107196. [Google Scholar]
- Hernández-Martínez, P.; Hernández-Rodríguez, C.S.; Van Rie, J.; Escriche, B.; Ferré, J. Insecticidal activity of Vip3Aa, Vip3Ad, Vip3Ae, and Vip3Af from Bacillus thuringiensis against lepidopteran corn pests. J. Invertebr. Pathol. 2013, 113, 78–81. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. J. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Van Rie, J.; Jansens, S.; Höfte, H.; Degheele, D.; Van Mellaert, H. Specificity of Bacillus thuringiensis delta-endotoxins. Importance of specific receptors on the brush border membrane of the mid-gut of target insects. Eur. J. Biochem. 1989, 186, 239–247. [Google Scholar]
- Wolfersberger, M.; Luethy, P.; Maurer, A.; Parenti, P.; Sacchi, F.V.; Giordana, B.; Hanozet, G.M. Preparation and partial characterization of amino acid transporting brush border membrane vesicles from the larval midgut of the cabbage butterfly (Pieris brassicae). Comp. Biochem. Physiol. 1987, 86, 301–308. [Google Scholar] [CrossRef]
- Munson, P.; Rodbard, D. LIGAND: A versatile computerized approach for characterization of ligand-binding systems. Anal. Biochem. 1980, 107, 220–239. [Google Scholar] [CrossRef]


{kind=link}
| Generation a | Protein | LC50 (95% FL) µg/cm2 | LC95 (95% FL) µg/cm2 | RR50 b | Slope ± SE | χ2 |
|---|---|---|---|---|---|---|
| Ms-S | Vip3Aa | 0.55 (0.26, 1.14) | 213 (66, 850) | / c | 0.71 ± 0.07 | 7.8 |
| F9 | Vip3Aa | >1684 | − d | >3061 | − | − |
| Ms-S | Cry1Ab | 0.14 (0.08, 0.23) | 6.65 (3.04, 20.8) | / | 0.99 ± 0.11 | 5.9 |
| F8 | Cry1Ab | 0.39 (0.20, 0.60) | 49 (15, 139) | 2.7 | 0.76 ± 0.08 | 6.9 |
| Ms-S | Cry1F | 1.73 (0.70, 4.25) | 271 (55, 8540) | / | 0.75 ± 0.10 | 13.9 |
| F9 | Cry1F | 2.60 (1.77, 3.69) | 60 (35, 125) | 1.5 | 1.29 ± 0.12 | 5.1 |
| Strains | Proteins | LC50 (95% FL) µg/cm2 | RR50 a | Slope ± SE | χ2 |
|---|---|---|---|---|---|
| Ms-S | Cry1Ab | 0.14 (0.08, 0.23) | / | 0.99 ± 0.11 | 5.9 |
| Cry1F | 1.95 (1.25, 3.08) | / | 0.97 ± 0.11 | 7.8 | |
| Ms-R | Cry1Ab | 0.39 (0.24, 0.62) | 3 | 0.93 ± 0.12 | 4.1 |
| Cry1F | 1.15 (0.93, 2.26) | / | 1.15 ± 0.12 | 10.5 |
| Insects | Kd (nM) | Rt (pmol/mg) |
|---|---|---|
| Ms-S | 40 ± 6 | 75 ± 22 |
| Ms-R | 41 ± 6 | 79 ± 17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quan, Y.; Yang, J.; Wang, Y.; Hernández-Martínez, P.; Ferré, J.; He, K. The Rapid Evolution of Resistance to Vip3Aa Insecticidal Protein in Mythimna separata (Walker) Is Not Related to Altered Binding to Midgut Receptors. Toxins 2021, 13, 364. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050364
Quan Y, Yang J, Wang Y, Hernández-Martínez P, Ferré J, He K. The Rapid Evolution of Resistance to Vip3Aa Insecticidal Protein in Mythimna separata (Walker) Is Not Related to Altered Binding to Midgut Receptors. Toxins. 2021; 13(5):364. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050364
Chicago/Turabian StyleQuan, Yudong, Jing Yang, Yueqin Wang, Patricia Hernández-Martínez, Juan Ferré, and Kanglai He. 2021. "The Rapid Evolution of Resistance to Vip3Aa Insecticidal Protein in Mythimna separata (Walker) Is Not Related to Altered Binding to Midgut Receptors" Toxins 13, no. 5: 364. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13050364