Concentration of Zearalenone, Alpha-Zearalenol and Beta-Zearalenol in the Myocardium and the Results of Isometric Analyses of the Coronary Artery in Prepubertal Gilts

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Experimental Feed
2.2. Clinical Observations
2.3. Concentrations of Zearalenone and Its Metabolites in the Heart Muscle
Carry-Over Factor
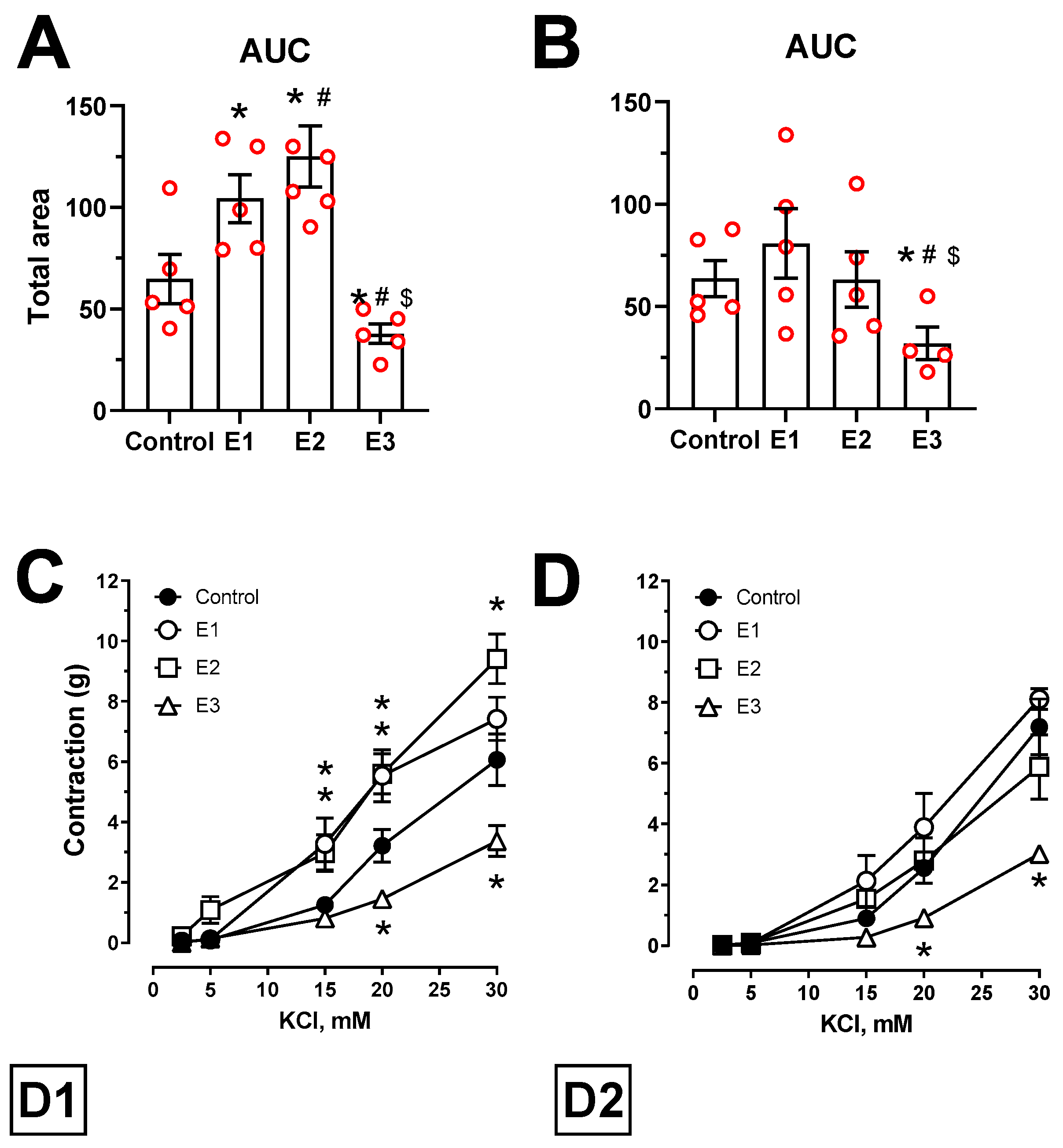
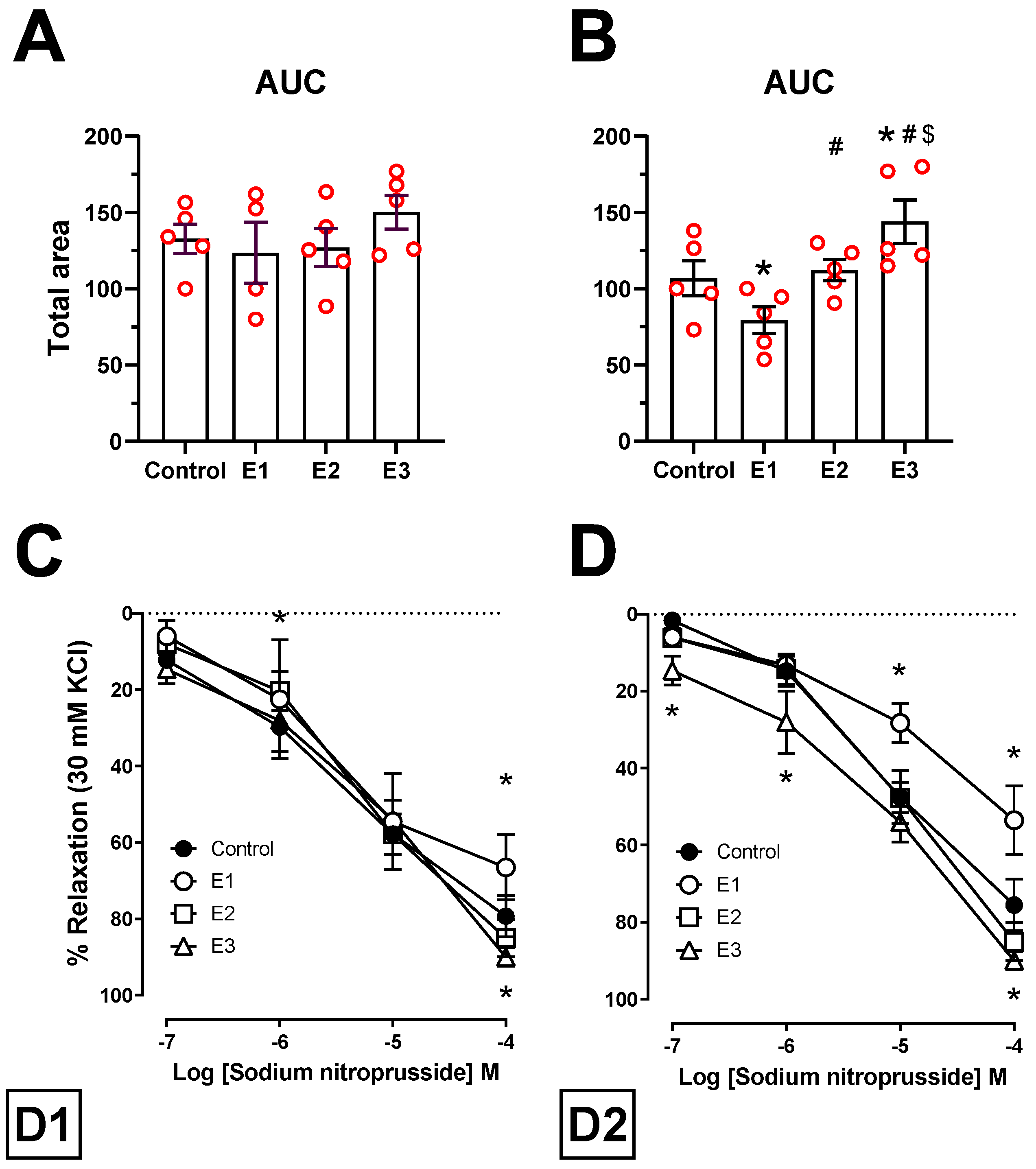
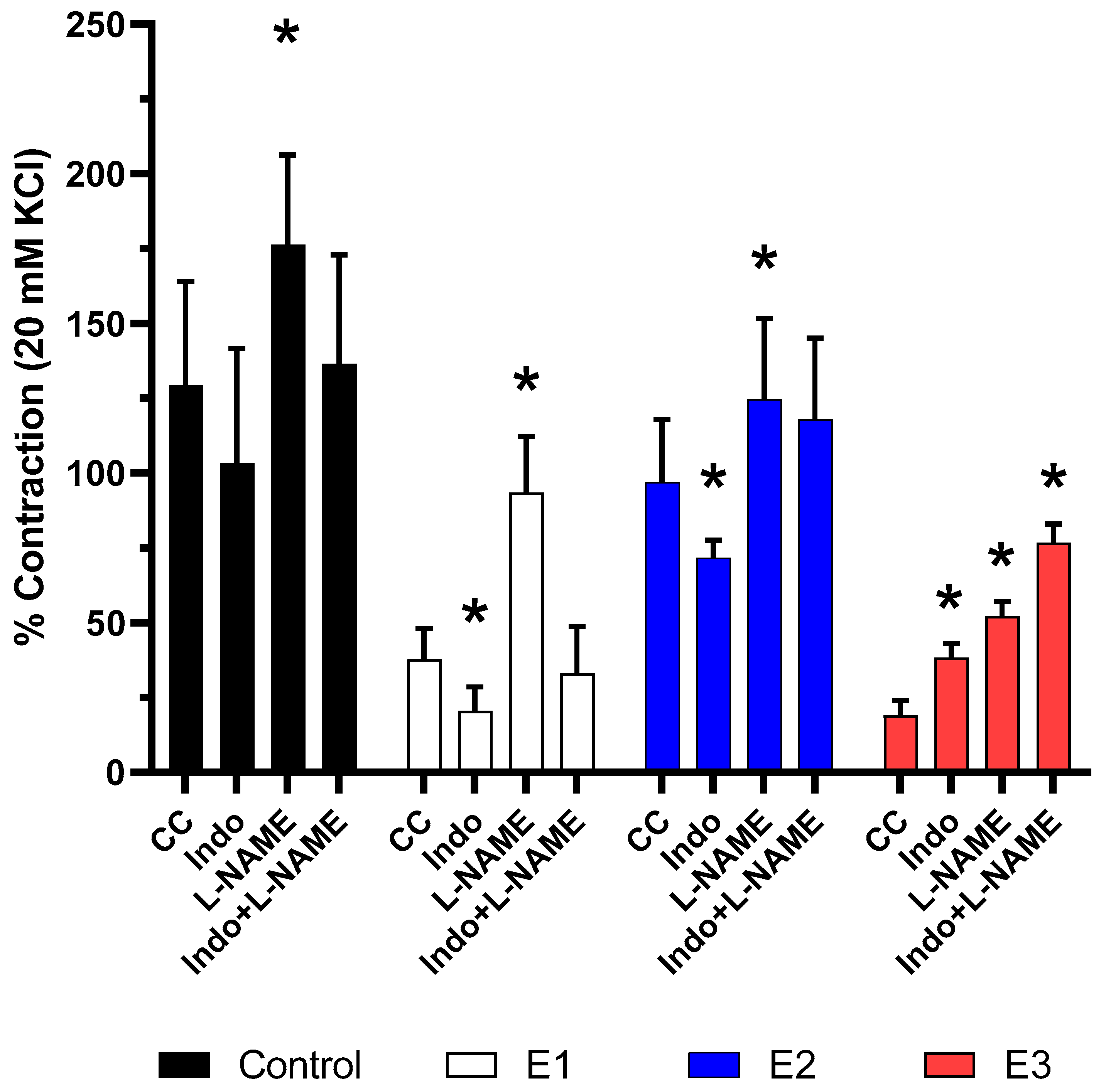
2.4. Vascular Reactivity Analyses
3. Discussion
3.1. Zearalenone and Its Metabolites in the Heart Muscle
3.2. Isometric Tension Analyses
3.3. Conclusions
4. Materials and Methods
4.1. In Vivo Study
4.1.1. General Information
4.1.2. Experimental Animals and Feed
4.1.3. Toxicological Analysis of Feed
4.1.4. Toxicological Analysis of the Apex of the Heart
Tissues Samples
Extraction Procedure
Chromatographic Quantification of ZEN and Its Metabolites
Carry-Over Factor
Statistical Analysis
4.2. Laboratory Analyses
4.2.1. Sampling for In Vitro Analyses
4.2.2. In Vitro Analysis
4.2.3. Drugs
4.2.4. Vascular Reactivity Analyses
4.2.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ropejko, K.; Twarużek, M. Zearalenone and Its Metabolites—General Overview, Occurrence, and Toxicity. Toxins 2021, 13, 35. [Google Scholar] [CrossRef] [PubMed]
- Wielogórska, E.; Elliott, C.; Danaher, M.; Connolly, L. Validation and application of a reporter gene assay for the determination of estrogenic endocrine disruptor activity in milk. Food Chem. Toxicol. 2014, 69, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. Activity of Zearalenone in the Porcine Intestinal Tract. Molecules 2016, 22, 18. [Google Scholar] [CrossRef] [Green Version]
- Rykaczewska, A.; Gajęcka, M.; Onyszek, E.; Cieplińska, K.; Dąbrowski, M.; Lisieska-Żołnierczyk, S.; Bulińska, M.; Babuchowski, A.; Gajęcki, M.T.; Zielonka, Ł. Imbalance in the Blood Concentrations of Selected Steroids in Pre-pubertal Gilts Depending on the Time of Exposure to Low Doses of Zearalenone. Toxins 2019, 11, 561. [Google Scholar] [CrossRef] [Green Version]
- Flores-Flores, M.E.; Lizarraga, E.; de Cerain, A.L.; González-Peñas, E. Presence of mycotoxins in animal milk: A review. Food Control. 2015, 53, 163–176. [Google Scholar] [CrossRef]
- Broekaert, N.; Devreese, M.; De Baere, S.; De Backer, P.; Croubels, S. Modified Fusarium mycotoxins unmasked: From occurrence in cereals to animal and human excretion. Food Chem. Toxicol. 2015, 80, 17–31. [Google Scholar] [CrossRef]
- Marin, D.E.; Pistol, G.; Neagoe, I.V.; Calin, L.; Taranu, I. Effects of zearalenone on oxidative stress and inflammation in weanling piglets. Food Chem. Toxicol. 2013, 58, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Martín, L.; Wood, K.; McEwen, P.; Smith, T.; Mandell, I.; Yannikouris, A.; Swanson, K. Effects of feeding corn naturally contaminated with Fusarium mycotoxins and/or a modified yeast cell wall extract on the performance, immunity and carcass characteristics of grain-fed veal calves. Anim. Feed. Sci. Technol. 2010, 159, 27–34. [Google Scholar] [CrossRef]
- Dunbar, B.; Patel, M.; Fahey, J.; Wira, C. Endocrine control of mucosal immunity in the female reproductive tract: Impact of environmental disruptors. Mol. Cell. Endocrinol. 2012, 354, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Dąbrowski, M.; Obremski, K.; Gajęcka, M.; Gajęcki, M.T.; Zielonka, Ł. Changes in the Subpopulations of Porcine Peripheral Blood Lymphocytes Induced by Exposure to Low Doses of Zearalenone (ZEN) and Deoxynivalenol (DON). Molecules 2016, 21, 557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alm, H.; Brüssow, K.-P.; Torner, H.; Vanselow, J.; Tomek, W.; Dänicke, S.; Tiemann, U. Influence of Fusarium-toxin contaminated feed on initial quality and meiotic competence of gilt oocytes. Reprod. Toxicol. 2006, 22, 44–50. [Google Scholar] [CrossRef]
- Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. The Effect of Low Monotonic Doses of Zearalenone on Selected Reproductive Tissues in Pre-Pubertal Female Dogs—A Review. Molecules 2015, 20, 20669–20687. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, K.; Habrowska-Górczyńska, D.E.; Piastowska-Ciesielska, A.W. Zearalenone as an endocrine disruptor in humans. Environ. Toxicol. Pharmacol. 2016, 48, 141–149. [Google Scholar] [CrossRef]
- Gajęcki, M. The effect of experimentally induced Fusarium mycotoxicosis on selected diagnostic and morphological parameters of the porcine digestive tract. In Proceedings of the Final Report for the National Centre for Research and Development in Warsaw, Poland, Development Project NR12-0080-10 entitled, Warsaw, Poland, 30 November 2013; pp. 1–180. [Google Scholar]
- Rykaczewska, A.; Gajęcka, M.; Dąbrowski, M.; Wiśniewska, A.; Szcześniewska, J.; Gajęcki, M.T.; Zielonka, Ł. Growth performance, selected blood biochemical parameters and body weights of pre-pubertal gilts fed diets supplemented with different doses of zearalenone (ZEN). Toxicon 2018, 152, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Zachariasova, M.; Dzumana, Z.; Veprikova, Z.; Hajkovaa, K.; Jiru, M.; Vaclavikova, M.; Zachariasova, A.; Pospichalova, M.; Florian, M.; Hajslova, J. Occurrence of multiple mycotoxins in European feedingstuffs, assessment of dietary intake by farm animals. Anim. Feed. Sci. Technol. 2014, 193, 124–140. [Google Scholar] [CrossRef]
- Gajęcka, M.; Tarasiuk, M.; Zielonka, Ł.; Dąbrowski, M.; Gajęcki, M. Risk assessment for changes in the metabolic profile and body weights of pre-pubertal gilts during long-term monotonic exposure to low doses of zearalenone (ZEN). Res. Veter. Sci. 2016, 109, 169–180. [Google Scholar] [CrossRef]
- Frizzell, C.; Ndossi, D.; Verhaegen, S.; Dahl, E.; Eriksen, G.S.; Sørlie, M.; Ropstad, E.; Muller, M.; Elliott, C.; Connolly, L. Endocrine disrupting effects of zearalenone, alpha- and beta-zearalenol at the level of nuclear receptor binding and steroidogenesis. Toxicol. Lett. 2011, 206, 210–217. [Google Scholar] [CrossRef]
- Kolle, S.N.; Ramirez, T.; Kamp, H.G.; Buesen, R.; Flick, B.; Strauss, V.; Van Ravenzwaay, B. A testing strategy for the identification of mammalian, systemic endocrine disruptors with particular focus on steroids. Regul. Toxicol. Pharmacol. 2012, 63, 259–278. [Google Scholar] [CrossRef]
- Gajęcka, M.; Zielonka, Ł.; Dabrowski, M.; Mróz, M.; Gajęcki, M. The effect of low doses of zearalenone and its metabolites on progesterone and 17β-estradiol concentrations in peripheral blood and body weights of pre-pubertal female Beagle dogs. Toxicon 2013, 76, 260–269. [Google Scholar] [CrossRef]
- Zielonka, Ł.; Waśkiewicz, A.; Beszterda, M.; Kostecki, M.; Dabrowski, M.; Obremski, K.; Goliński, P.; Gajęcki, M. Zearalenone in the Intestinal Tissues of Immature Gilts Exposed per os to Mycotoxins. Toxins 2015, 7, 3210–3223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchais-Oberwinkler, S.; Henn, C.; Möller, G.; Klein, T.; Negri, M.; Oster, A.; Spadaro, A.; Werth, R.; Wetzel, M.; Xu, K.; et al. 17β-Hydroxysteroid dehydrogenases (17β-HSDs) as therapeutic targets: Protein structures, functions, and recent progress in inhibitor development. J. Steroid Biochem. Mol. Biol. 2011, 125, 66–82. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Knutsen, H.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; DiNovi, M.; Edler, L.; et al. Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, e04851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaller, T.H.; Snyder, D.J.; Spasojevic, I.; Gedeon, P.C.; Sanchez-Perez, L.; Sampson, J.H. First in human dose calculation of a single-chain bispecific antibody targeting glioma using the MABEL approach. J. Immunother. Cancer 2019, 8, e000213. [Google Scholar] [CrossRef] [Green Version]
- Cieplińska, K.; Gajęcka, M.; Nowak, A.; Dąbrowski, M.; Łukasz, Z.; Gajęcki, M.T. The Genotoxicity of Caecal Water in Gilts Exposed to Low Doses of Zearalenone. Toxins 2018, 10, 350. [Google Scholar] [CrossRef] [Green Version]
- Cieplińska, K.; Gajęcka, M.; Dąbrowski, M.; Rykaczewska, A.; Lisieska-Żołnierczyk, S.; Bulińska, M.; Zielonka, Ł.; Gajęcki, M.T. Time-Dependent Changes in the Intestinal Microbiome of Gilts Exposed to Low Zearalenone Doses. Toxins 2019, 11, 296. [Google Scholar] [CrossRef] [Green Version]
- Stopa, E.; Babińska, I.; Zielonka, Ł.; Gajecki, M.; Gajecka, M. Immunohistochemical evaluation of apoptosis and proliferation in the mucous membrane of selected uterine regions in pre-pubertal bitches exposed to low doses of zearalenone. Pol. J. Veter. Sci. 2016, 19, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Alassane-Kpembi, I.; Pinton, P.; Oswald, I.P. Effects of Mycotoxins on the Intestine. Toxins 2019, 11, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, H.; Ham, W.V.D.; Slob, W.; Pieters, M. Conversion Factors Estimating Indicative Chronic No-Observed-Adverse-Effect Levels from Short-Term Toxicity Data. Regul. Toxicol. Pharmacol. 1996, 23, 249–255. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R., Jr.; Lee, D.H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and Endocrine-Disrupting Chemicals: Low-Dose Effects and Nonmonotonic Dose Responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Applegate, T.J. Modulation of Intestinal Functions Following Mycotoxin Ingestion: Meta-Analysis of Published Experiments in Animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickey, G.L.; Craig, P.S.; Luttik, R.; De Zwart, D. On the quantification of intertest variability in ecotoxicity data with application to species sensitivity distributions. Environ. Toxicol. Chem. 2012, 31, 1903–1910. [Google Scholar] [CrossRef] [Green Version]
- Pastoor, T.P.; Bachman, A.N.; Bell, D.R.; Cohen, S.M.; Dellarco, M.; Dewhurst, I.C.; Doe, J.E.; Doerrer, N.G.; Embry, M.R.; Hines, R.N.; et al. A 21st century roadmap for human health risk assessment. Crit. Rev. Toxicol. 2014, 44, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Chain, E.P.O.C.I.T.F. Scientific Opinion on the risks for public health related to the presence of zearalenone in food. EFSA J. 2011, 9, 1–124. [Google Scholar] [CrossRef]
- Gajęcka, M.; Przybylska-Gornowicz, B. The low doses effect of experimental zearalenone (ZEN) intoxication on the presence of Ca2+ in selected ovarian cells from pre-pubertal bitches. Pol. J. Veter. Sci. 2012, 15, 711–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qadri, S.M.; Bissinger, R.; Solh, Z.; Oldenborg, P.-A. Eryptosis in health and disease: A paradigm shift towards understanding the (patho)physiological implications of programmed cell death of erythrocytes. Blood Rev. 2017, 31, 349–361. [Google Scholar] [CrossRef]
- Pyrshev, K.A.; Klymchenko, A.S.; Csúcs, G.; Demchenko, A.P. Apoptosis and eryptosis: Striking differences on biomembrane level. Biochim. Biophys. Acta (BBA) Biomembr. 2018, 1860, 1362–1371. [Google Scholar] [CrossRef] [PubMed]
- Lang, E.; Lang, F. Mechanisms and pathophysiological significance of eryptosis, the suicidal erythrocyte death. Semin. Cell Dev. Biol. 2015, 39, 35–42. [Google Scholar] [CrossRef]
- Gajęcka, M.; Rybarczyk, L.; Jakimiuk, E.; Zielonka, Ł.; Obremski, K.; Zwierzchowski, W.; Gajęcki, M. The effect of experimental long-term exposure to low-dose zearalenone on uterine histology in sexually immature gilts. Exp. Toxicol. Pathol. 2012, 64, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Stopa, E.; Tarasiuk, M.; Zielonka, Ł.; Gajęcki, M. The Expression of Type-1 and Type-2 Nitric Oxide Synthase in Selected Tissues of the Gastrointestinal Tract during Mixed Mycotoxicosis. Toxins 2013, 5, 2281–2292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broadley, K.J.; Broadley, H.D. Non-adrenergic vasoconstriction and vasodilatation of guinea-pig aorta by β-phenylethylamine and amphetamine—Role of nitric oxide determined with L-NAME and NO scavengers. Eur. J. Pharmacol. 2018, 818, 198–205. [Google Scholar] [CrossRef]
- Lawrenz, B.; Melado, L.; Fatemi, H. Premature progesterone rise in ART-cycles. Reprod. Biol. 2018, 18, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. Hormesis: Path and Progression to Significance. Int. J. Mol. Sci. 2018, 19, 2871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.; Wang, L.; Wang, J.; Tan, Y.; Yu, D.; Chang, X.; Fan, Y.; Zhao, D.; Wang, C.; De Boevre, M.; et al. A QuEChERS-Based Liquid Chromatography-Tandem Mass Spectrometry Method for the Simultaneous Determination of Nine Zearalenone-Like Mycotoxins in Pigs. Toxins 2018, 10, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Ao, X.; Lei, Y.; Ji, C.; Ma, Q. Bacillus subtilis ANSB01G culture alleviates oxidative stress and cell apoptosis induced by dietary zearalenone in first-parity gestation sows. Anim. Nutr. 2020, 6, 372–378. [Google Scholar] [CrossRef]
- Lee, H.-J.; Park, J.-H.; Oh, S.-Y.; Cho, D.-H.; Kim, S.; Jo, I. Zearalenone-Induced Interaction between PXR and Sp1 Increases Binding of Sp1 to a Promoter Site of the eNOS, Decreasing Its Transcription and NO Production in BAECs. Toxins 2020, 12, 421. [Google Scholar] [CrossRef]
- Gajęcki, M.T.; Gajęcka, M.; Zielonka, Ł. Mycotoxins Occurence in Feed and Their Influence on Animal Health. In Printed Edition of the Special Issue Published in Toxins; Multidisciplinary Digital Publishing Institute: Basel, Switzerland, 2021; pp. 1–242. ISBN 978-3-03943-848-8. [Google Scholar]
- Villarreal, F. In Pursuit of Understanding the Role of Estrogens in Regulating Cardiac Structure and Function. JACC Basic Transl. Sci. 2020, 5, 913–915. [Google Scholar] [CrossRef]
- Firth, J.M.; Yang, H.-Y.; Francis, A.J.; Islam, N.; MacLeod, K.T. The Effect of Estrogen on Intracellular Ca2+ and Na+ Regulation in Heart Failure. JACC Basic Transl. Sci. 2020, 5, 901–912. [Google Scholar] [CrossRef] [PubMed]
- Mahmoodzadeh, S.; Dworatzek, E. The Role of 17β-Estradiol and Estrogen Receptors in Regulation of Ca2+ Channels and Mitochondrial Function in Cardiomyocytes. Front. Endocrinol. 2019, 10, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielonka, Ł.; Gajęcka, M.; Lisieska-Żołnierczyk, S.; Dąbrowski, M.; Gajęcki, M.T. The Effect of Different Doses of Zearalenone in Feed on the Bioavailability of Zearalenone and Alpha-Zearalenol, and the Concentrations of Estradiol and Testosterone in the Peripheral Blood of Pre-Pubertal Gilts. Toxins 2020, 12, 144. [Google Scholar] [CrossRef] [Green Version]
- Benagiano, M.; Bianchi, P.; D’Elios, M.M.; Brosens, I.; Benagiano, G. Autoimmune diseases: Role of steroid hormones. Best Pract. Res. Clin. Obstet. Gynaecol. 2019, 60, 24–34. [Google Scholar] [CrossRef]
- Long, V.; Fiset, C. Contribution of estrogen to the pregnancy-induced increase in cardiac automaticity. J. Mol. Cell. Cardiol. 2020, 147, 27–34. [Google Scholar] [CrossRef]
- Ueda, K.; Adachi, Y.; Liu, P.; Fukuma, N.; Takimoto, E. Regulatory Actions of Estrogen Receptor Signaling in the Cardiovascular System. Front. Endocrinol. 2020, 10, 909. [Google Scholar] [CrossRef]
- Luisetto, M.; Naseer, A.; Nili, A.B.; Abdul, H.G.; Rasool, M.G.; Rehman, K.K.; Ahmadabadi, B.N.; Luca, C. Endogenus toxicology: Modern physio-pathological aspects and relationship with new therapeutic strategies. An integrative discipline incorporating concepts from different research discipline like Biochemistry, Pharmacology and Toxicology. Arch. Cancer Sci. Ther. 2019, 3, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.P.; Schuhmacher, M.; Kumar, V. Review on crosstalk and common mechanisms of endocrine disruptors: Scaffolding to improve PBPK/PD model of EDC mixture. Environ. Int. 2017, 99, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gräser, T.; Leisner, H.; Vedernikov, Y.P.; Tiedt, N. The action of acetylcholine on isolated coronary arteries of different species. Cor. Vasa 1987, 29, 70–80. [Google Scholar] [PubMed]
- Bryan, R.M.; You, J.; Golding, E.M.; Marrelli, S.P. Endothelium-derived Hyperpolarizing Factor. Anesthesiology 2005, 102, 1261–1277. [Google Scholar] [CrossRef]
- Meerpoel, C.; Vidal, A.; Tangni, E.K.; Huybrechts, B.; Couck, L.; De Rycke, R.; De Bels, L.; De Saeger, S.; Broeck, W.V.D.; Devreese, M.; et al. A Study of Carry-Over and Histopathological Effects after Chronic Dietary Intake of Citrinin in Pigs, Broiler Chickens and Laying Hens. Toxins 2020, 12, 719. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weeks of Exposure | Feed Intake [kg/day] | Total doses of ZEN in Groups Respectively [µg/kg BW] | Group E1 [ng/g] (%) | Carry-over Factor | Group E2 [ng/g]/(%) | Carry-over Factor | Group E3 [ng/g]/(%) | Carry-over Factor |
|---|---|---|---|---|---|---|---|---|
| Zearalenone | ||||||||
| D1 | 0.8 | 80.5/161.9/242.7 | 0.691 ± 0.635 (100%) | 8 × 10−6 | 2.275 ± 2.22 (86.69%) | 14 × 10−6 | 1.387 ± 1.93(78.22%) | 17 × 10−6 |
| D2 | 1.1 | 101.01/196.9/298.2 | 0.977 ± 0.579 (73.62%) | 9 × 10−6 | 2.621 ± 1.499 (91.48%) | 13 × 10−6 | 4.89 ± 4.405(93.42%) | 16 × 10−6 |
| α-ZEL | ||||||||
| D1 | not applicable | not applicable | 0.0 ± 0.0 (0%) | 0 | 0.316 ± 0.061 (12.04%) | 19 × 10−7 | 0.353 ± 0.104(19.9%) | 14 × 10−7 |
| D2 | not applicable | not applicable | 0.146 ± 0.143 (11%) | 14 × 10−7 | 0.167 ± 0.146 (5.82%) | 1 × 10−6 | 0.312 ± 0.213(5.96%) | 1 × 10−6 |
| β-ZEL | ||||||||
| D1 | not applicable | not applicable | 0.0 ± 0.0 (0%) | 0 | 0.033 ± 0.029 (1.25%) | 2 × 10−7 | 0.033 ± 0.004(1.86%) | 1 × 10−7 |
| D2 | not applicable | not applicable | 0.204 ± 0.046 (15.37%) | 2 × 10−6 | 0.077 ± 0.017 **(2.68%) | 4 × 10−7 | 0.032 ± 0.004 **(0.61%) | 1 × 10−7 |
| Control | Group E1 | Group E2 | Group E3 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AUC | Emax (%) | pD2 | AUC | Emax (%) | pD2 | AUC | Emax (%) | pD2 | AUC | Emax (%) | pD2 | |
| D1ACh | 122.6 ± 15.01 | 147.8 ± 11.33 | 5.801 ± 0.077 | 69.93 ± 11.20* | 82.87 ± 7.853* | 5.788 ± 0.103 | 162.6 ± 14.58* | 155.1 ± 10.69 | 6.087 ± 0.074 | 32.73 ± 6.205* | 51.08 ± 13.19* | 5.606 ± 0.210 |
| D2ACh | 119.8 ± 19.10 | 135 ± 15.39 | 5.859 ± 0.112 | 58.29 ± 13.33* | 70.89 ± 10.28* | 5.831 ± 0.149 | 137.6 ± 9.264 | 111.3 ± 10.30 | 6.263 ± 0.101* | 29.81 ± 5.434* | 33.13 ± 4.47* | 5.876 ± 0.129 |
| D1SNP | 137.5 ± 10.36 | 76.9 ± 5.03 | 5.469 ± 0.2543 | 116.3 ± 21.18 | 67.74 ± 9.994 | 5.605 ± 0.433 | 128.6 ± 14.23 | 89.5 ± 5.012 | 5.217 ± 0.153 | 134.3 ± 12.20 | 97.78 ± 7.674 | 4.952 ± 0.175 |
| D2SNP | 101.6 ± 18.32 | 79.99 ± 6.529 | 5.171 ± 0.175 | 74.25 ± 11.65* | 60.12 ± 8.205* | 4.833 ± 0.273 | 110.5 ± 13.74 | 93.53 ± 5.480 | 4.963 ± 0.116 | 141.7 ± 12.62* | 97.78 ± 7.674* | 4.952 ± 0.175 |
| Ingredient | Manufacturer’s Declared Composition (%) |
|---|---|
| Soybean meal | 16 |
| Wheat | 55 |
| Barley | 22 |
| Wheat bran | 4.0 |
| Chalk | 0.3 |
| Zitrosan | 0.2 |
| Vitamin-mineral premix 1 | 2.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gajęcka, M.; Majewski, M.S.; Zielonka, Ł.; Grzegorzewski, W.; Onyszek, E.; Lisieska-Żołnierczyk, S.; Juśkiewicz, J.; Babuchowski, A.; Gajęcki, M.T. Concentration of Zearalenone, Alpha-Zearalenol and Beta-Zearalenol in the Myocardium and the Results of Isometric Analyses of the Coronary Artery in Prepubertal Gilts. Toxins 2021, 13, 396. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13060396
Gajęcka M, Majewski MS, Zielonka Ł, Grzegorzewski W, Onyszek E, Lisieska-Żołnierczyk S, Juśkiewicz J, Babuchowski A, Gajęcki MT. Concentration of Zearalenone, Alpha-Zearalenol and Beta-Zearalenol in the Myocardium and the Results of Isometric Analyses of the Coronary Artery in Prepubertal Gilts. Toxins. 2021; 13(6):396. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13060396
Chicago/Turabian StyleGajęcka, Magdalena, Michał S. Majewski, Łukasz Zielonka, Waldemar Grzegorzewski, Ewa Onyszek, Sylwia Lisieska-Żołnierczyk, Jerzy Juśkiewicz, Andrzej Babuchowski, and Maciej T. Gajęcki. 2021. "Concentration of Zearalenone, Alpha-Zearalenol and Beta-Zearalenol in the Myocardium and the Results of Isometric Analyses of the Coronary Artery in Prepubertal Gilts" Toxins 13, no. 6: 396. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13060396