Helicobacter pylori BabA–SabA Key Roles in the Adherence Phase: The Synergic Mechanism for Successful Colonization and Disease Development

 , , and
, , and 
Abstract
:1. Introduction
2. BabA: History, Gene, and Structure
3. SabA: History, Gene, and Structure
4. Production and Regulation of BabA and SabA in Transcriptional and Translational Levels
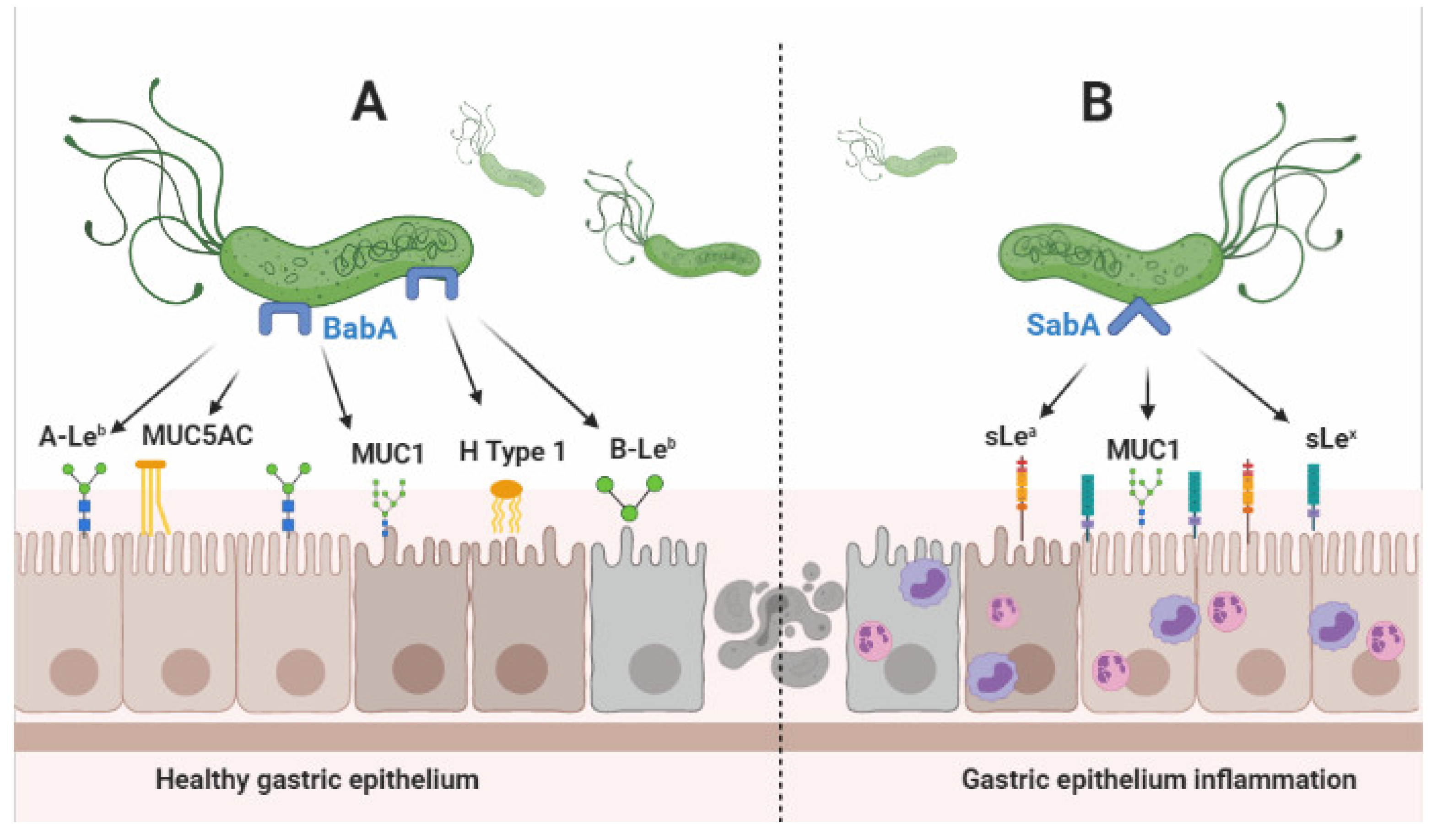
5. BabA and SabA Interaction with Host Cell
6. BabA, SabA, and Disease Development
7. Association with Other Virulence Factors
8. Future Insights
9. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hooi, J.K.Y.; Lai, W.Y.; Ng, W.K.; Suen, M.M.Y.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.S.; Wu, J.C.Y.; et al. Global Prevalence of Helicobacter pylori Infection: Systematic Review and Meta-Analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef] [Green Version]
- Hessey, S.J.; Spencer, J.; Wyatt, J.I.; Sobala, G.; Rathbone, B.J.; Axon, A.T.; Dixon, M.F. Bacterial adhesion and disease activity in Helicobacter associated chronic gastritis. Gut 1990, 31, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, S.; Konradt, M.; Groll, C.; Scheid, P.; Hanauer, G.; Werling, H.O.; Josenhans, C.; Suerbaum, S. The spatial orientation of Helicobacter pylori in the gastric mucus. Proc. Natl. Acad. Sci. USA 2004, 101, 5024–5029. [Google Scholar] [CrossRef] [Green Version]
- Peek, R.M., Jr.; Blaser, M.J. Helicobacter pylori and gastrointestinal tract adenocarcinomas. Nat. Rev. Cancer 2002, 2, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Blaser, M.J. Helicobacter pylori and gastric diseases. BMJ 1998, 316, 1507–1510. [Google Scholar] [CrossRef] [PubMed]
- Wroblewski, L.E.; Peek, R.M., Jr.; Wilson, K.T. Helicobacter pylori and gastric cancer: Factors that modulate disease risk. Clin. Microbiol. Rev. 2010, 23, 713–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilver, D.; Arnqvist, A.; Ogren, J.; Frick, I.M.; Kersulyte, D.; Incecik, E.T.; Berg, D.E.; Covacci, A.; Engstrand, L.; Borén, T. Helicobacter pylori adhesin binding fucosylated histo-blood group antigens revealed by retagging. Science 1998, 279, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Hage, N.; Howard, T.; Phillips, C.; Brassington, C.; Overman, R.; Debreczeni, J.; Gellert, P.; Stolnik, S.; Winkler, G.S.; Falcone, F.H. Structural basis of Lewis(b) antigen binding by the Helicobacter pylori adhesin BabA. Sci. Adv. 2015, 1, e1500315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pride, D.T.; Meinersmann, R.J.; Blaser, M.J. Allelic Variation within Helicobacter pylori babA and babB. Infect. Immun. 2001, 69, 1160–1171. [Google Scholar] [CrossRef] [Green Version]
- Backert, S.; Clyne, M.; Tegtmeyer, N. Molecular mechanisms of gastric epithelial cell adhesion and injection of CagA by Helicobacter pylori. Cell Commun. Signal. CCS 2011, 9, 28. [Google Scholar] [CrossRef] [Green Version]
- Yamaoka, Y. Roles of Helicobacter pylori BabA in gastroduodenal pathogenesis. World J. Gastroenterol. 2008, 14, 4265–4272. [Google Scholar] [CrossRef] [Green Version]
- Matteo, M.J.; Armitano, R.I.; Romeo, M.; Wonaga, A.; Olmos, M.; Catalano, M. Helicobacter pylori bab genes during chronic colonization. Int. J. Mol. Epidemiol. Genet. 2011, 2, 286–291. [Google Scholar] [PubMed]
- Ansari, S.; Kabamba, E.T.; Shrestha, P.K.; Aftab, H.; Myint, T.; Tshering, L.; Sharma, R.P.; Ni, N.; Aye, T.T.; Subsomwong, P.; et al. Helicobacter pylori bab characterization in clinical isolates from Bhutan, Myanmar, Nepal and Bangladesh. PLoS ONE 2017, 12, e0187225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennig, E.E.; Allen, J.M.; Cover, T.L. Multiple chromosomal loci for the babA gene in Helicobacter pylori. Infect. Immun. 2006, 74, 3046–3051. [Google Scholar] [CrossRef] [Green Version]
- Colbeck, J.C.; Hansen, L.M.; Fong, J.M.; Solnick, J.V. Genotypic profile of the outer membrane proteins BabA and BabB in clinical isolates of Helicobacter pylori. Infect. Immun. 2006, 74, 4375–4378. [Google Scholar] [CrossRef] [Green Version]
- Kim, A.; Servetas, S.L.; Kang, J.; Kim, J.; Jang, S.; Cha, H.J.; Lee, W.J.; Kim, J.; Romero-Gallo, J.; Peek, R.M., Jr.; et al. Helicobacter pylori bab Paralog Distribution and Association with cagA, vacA, and homA/B Genotypes in American and South Korean Clinical Isolates. PLoS ONE 2015, 10, e0137078. [Google Scholar] [CrossRef]
- Pang, S.S.; Nguyen, S.T.S.; Perry, A.J.; Day, C.J.; Panjikar, S.; Tiralongo, J.; Whisstock, J.C.; Kwok, T. The three-dimensional structure of the extracellular adhesion domain of the sialic acid-binding adhesin SabA from Helicobacter pylori. J. Biol. Chem. 2014, 289, 6332–6340. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, A.C.; Weinberger, D.M.; Ford, C.B.; Nelson, J.C.; Snider, J.D.; Hall, J.D.; Paules, C.I.; Peek, R.M.; Forsyth, M.H. Expression of the Helicobacter pylori adhesin SabA is controlled via phase variation and the ArsRS signal transduction system. Microbiology 2008, 154, 2231–2240. [Google Scholar] [CrossRef] [Green Version]
- Mahdavi, J.; Sondén, B.; Hurtig, M.; Olfat, F.O.; Forsberg, L.; Roche, N.; Angstrom, J.; Larsson, T.; Teneberg, S.; Karlsson, K.A.; et al. Helicobacter pylori SabA adhesin in persistent infection and chronic inflammation. Science 2002, 297, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Bäckström, A.; Lundberg, C.; Kersulyte, D.; Berg, D.E.; Borén, T.; Arnqvist, A. Metastability of Helicobacter pylori bab adhesin genes and dynamics in Lewis b antigen binding. Proc. Natl. Acad. Sci. USA 2004, 101, 16923–16928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solnick, J.V.; Hansen, L.M.; Salama, N.R.; Boonjakuakul, J.K.; Syvanen, M. Modification of Helicobacter pylori outer membrane protein expression during experimental infection of rhesus macaques. Proc. Natl. Acad. Sci. USA 2004, 101, 2106–2111. [Google Scholar] [CrossRef] [Green Version]
- Styer, C.M.; Hansen, L.M.; Cooke, C.L.; Gundersen, A.M.; Choi, S.S.; Berg, D.E.; Benghezal, M.; Marshall, B.J.; Peek, R.M., Jr.; Borén, T.; et al. Expression of the BabA adhesin during experimental infection with Helicobacter pylori. Infect. Immun. 2010, 78, 1593–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohno, T.; Vallstrom, A.; Rugge, M.; Ota, H.; Graham, D.Y.; Arnqvist, A.; Yamaoka, Y. Effects of blood group antigen-binding adhesin expression during Helicobacter pylori infection of Mongolian gerbils. J. Infect. Dis. 2011, 203, 726–735. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Kwon, D.H.; Graham, D.Y. A M(r) 34,000 proinflammatory outer membrane protein (oipA) of Helicobacter pylori. Proc. Natl. Acad. Sci. USA 2000, 97, 7533–7538. [Google Scholar] [CrossRef] [Green Version]
- Alm, R.A.; Ling, L.S.; Moir, D.T.; King, B.L.; Brown, E.D.; Doig, P.C.; Smith, D.R.; Noonan, B.; Guild, B.C.; deJonge, B.L.; et al. Genomic-sequence comparison of two unrelated isolates of the human gastric pathogen Helicobacter pylori. Nature 1999, 397, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Tomb, J.F.; White, O.; Kerlavage, A.R.; Clayton, R.A.; Sutton, G.G.; Fleischmann, R.D.; Ketchum, K.A.; Klenk, H.P.; Gill, S.; Dougherty, B.A.; et al. The complete genome sequence of the gastric pathogen Helicobacter pylori. Nature 1997, 388, 539–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooke, C.L.; An, H.J.; Kim, J.; Canfield, D.R.; Torres, J.; Lebrilla, C.B.; Solnick, J.V. Modification of gastric mucin oligosaccharide expression in rhesus macaques after infection with Helicobacter pylori. Gastroenterology 2009, 137, 1061–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loh, J.T.; Beckett, A.C.; Scholz, M.B.; Cover, T.L. High-Salt Conditions Alter Transcription of Helicobacter pylori Genes Encoding Outer Membrane Proteins. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loh, J.T.; Torres, V.J.; Cover, T.L. Regulation of Helicobacter pylori cagA expression in response to salt. Cancer Res. 2007, 67, 4709–4715. [Google Scholar] [CrossRef] [Green Version]
- Yamaoka, Y.; Ojo, O.; Fujimoto, S.; Odenbreit, S.; Haas, R.; Gutierrez, O.; El-Zimaity, H.M.; Reddy, R.; Arnqvist, A.; Graham, D.Y. Helicobacter pylori outer membrane proteins and gastroduodenal disease. Gut 2006, 55, 775–781. [Google Scholar] [CrossRef] [Green Version]
- Yamaoka, Y.; Kita, M.; Kodama, T.; Imamura, S.; Ohno, T.; Sawai, N.; Ishimaru, A.; Imanishi, J.; Graham, D.Y. Helicobacter pylori infection in mice: Role of outer membrane proteins in colonization and inflammation. Gastroenterology 2002, 123, 1992–2004. [Google Scholar] [CrossRef]
- Aspholm-Hurtig, M.; Dailide, G.; Lahmann, M.; Kalia, A.; Ilver, D.; Roche, N.; Vikström, S.; Sjöström, R.; Lindén, S.; Bäckström, A.; et al. Functional adaptation of BabA, the H. pylori ABO blood group antigen binding adhesin. Science 2004, 305, 519–522. [Google Scholar] [CrossRef]
- Skindersoe, M.E.; Rasmussen, L.; Andersen, L.P.; Krogfelt, K.A. A Novel Assay for Easy and Rapid Quantification of Helicobacter pylori Adhesion. Helicobacter 2015, 20, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Saberi, S.; Schmidt, A.; Eybpoosh, S.; Esmaili, M.; Talebkhan, Y.; Mohajerani, N.; Oghalaie, A.; Eshagh Hosseini, M.; Mohagheghi, M.A.; Bugaytova, J.; et al. Helicobacter pylori Strains from Duodenal Ulcer Patients Exhibit Mixed babA/B Genotypes with Low Levels of BabA Adhesin and Lewis b Binding. Dig. Dis. Sci. 2016, 61, 2868–2877. [Google Scholar] [CrossRef] [PubMed]
- Lindén, S.; Nordman, H.; Hedenbro, J.; Hurtig, M.; Borén, T.; Carlstedt, I. Strain- and blood group-dependent binding of Helicobacter pylori to human gastric MUC5AC glycoforms. Gastroenterology 2002, 123, 1923–1930. [Google Scholar] [CrossRef] [PubMed]
- Lindén, S.K.; Wickström, C.; Lindell, G.; Gilshenan, K.; Carlstedt, I. Four modes of adhesion are used during Helicobacter pylori binding to human mucins in the oral and gastric niches. Helicobacter 2008, 13, 81–93. [Google Scholar] [CrossRef]
- Sakamoto, S.; Watanabe, T.; Tokumaru, T.; Takagi, H.; Nakazato, H.; Lloyd, K.O. Expression of Lewisa, Lewisb, Lewisx, Lewisy, siayl-Lewisa, and sialyl-Lewisx blood group antigens in human gastric carcinoma and in normal gastric tissue. Cancer Res. 1989, 49, 745–752. [Google Scholar]
- Ota, H.; Nakayama, J.; Momose, M.; Hayama, M.; Akamatsu, T.; Katsuyama, T.; Graham, D.Y.; Genta, R.M. Helicobacter pylori infection produces reversible glycosylation changes to gastric mucins. Virchows Arch. Int. J. Pathol. 1998, 433, 419–426. [Google Scholar] [CrossRef]
- Lindén, S.; Borén, T.; Dubois, A.; Carlstedt, I. Rhesus monkey gastric mucins: Oligomeric structure, glycoforms and Helicobacter pylori binding. Biochem. J. 2004, 379, 765–775. [Google Scholar] [CrossRef] [Green Version]
- Falk, P.G.; Bry, L.; Holgersson, J.; Gordon, J.I. Expression of a human alpha-1,3/4-fucosyltransferase in the pit cell lineage of FVB/N mouse stomach results in production of Leb-containing glycoconjugates: A potential transgenic mouse model for studying Helicobacter pylori infection. Proc. Natl. Acad. Sci. USA 1995, 92, 1515–1519. [Google Scholar] [CrossRef] [Green Version]
- Bugaytsova, J.A.; Björnham, O.; Chernov, Y.A.; Gideonsson, P.; Henriksson, S.; Mendez, M.; Sjöström, R.; Mahdavi, J.; Shevtsova, A.; Ilver, D.; et al. Helicobacter pylori Adapts to Chronic Infection and Gastric Disease via pH-Responsive BabA-Mediated Adherence. Cell Host Microbe 2017, 21, 376–389. [Google Scholar] [CrossRef] [Green Version]
- Benktander, J.; Ångström, J.; Breimer, M.E.; Teneberg, S. Redefinition of the carbohydrate binding specificity of Helicobacter pylori BabA adhesin. J. Biol. Chem. 2012, 287, 31712–31724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindén, S.K.; Sheng, Y.H.; Every, A.L.; Miles, K.M.; Skoog, E.C.; Florin, T.H.; Sutton, P.; McGuckin, M.A. MUC1 limits Helicobacter pylori infection both by steric hindrance and by acting as a releasable decoy. PLoS Pathog. 2009, 5, e1000617. [Google Scholar] [CrossRef] [PubMed]
- Walz, A.; Odenbreit, S.; Mahdavi, J.; Borén, T.; Ruhl, S. Identification and characterization of binding properties of Helicobacter pylori by glycoconjugate arrays. Glycobiology 2005, 15, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Sheu, B.S.; Sheu, S.M.; Yang, H.B.; Huang, A.H.; Wu, J.J. Host gastric Lewis expression determines the bacterial density of Helicobacter pylori in babA2 genopositive infection. Gut 2003, 52, 927–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimoto, S.; Olaniyi Ojo, O.; Arnqvist, A.; Wu, J.Y.; Odenbreit, S.; Haas, R.; Graham, D.Y.; Yamaoka, Y. Helicobacter pylori BabA expression, gastric mucosal injury, and clinical outcome. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2007, 5, 49–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Woo, C.W.; Lee, Y.M.; Son, B.R.; Kim, J.W.; Chae, H.B.; Youn, S.J.; Park, S.M. Genotyping CagA, VacA subtype, IceA1, and BabA of Helicobacter pylori isolates from Korean patients, and their association with gastroduodenal diseases. J. Korean Med. Sci. 2001, 16, 579–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.Y.; He, C.Y.; Meng, X.; Yuan, Y. Association of Helicobacter pylori babA2 with peptic ulcer disease and gastric cancer. World J. Gastroenterol. 2013, 19, 4242–4251. [Google Scholar] [CrossRef]
- Talebi Bezmin Abadi, A.; Taghvaei, T.; Mohabbati Mobarez, A.; Vaira, G.; Vaira, D. High correlation of babA 2-positive strains of Helicobacter pylori with the presence of gastric cancer. Intern. Emerg. Med. 2013, 8, 497–501. [Google Scholar] [CrossRef]
- Sugimoto, M.; Ohno, T.; Graham, D.Y.; Yamaoka, Y. Helicobacter pylori outer membrane proteins on gastric mucosal interleukin 6 and 11 expression in Mongolian gerbils. J. Gastroenterol. Hepatol. 2011, 26, 1677–1684. [Google Scholar] [CrossRef] [Green Version]
- Unemo, M.; Aspholm-Hurtig, M.; Ilver, D.; Bergström, J.; Borén, T.; Danielsson, D.; Teneberg, S. The sialic acid binding SabA adhesin of Helicobacter pylori is essential for nonopsonic activation of human neutrophils. J. Biol. Chem. 2005, 280, 15390–15397. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, T.; Naito, Y. The role of neutrophils and inflammation in gastric mucosal injury. Free Radic. Res. 2000, 33, 785–794. [Google Scholar] [CrossRef]
- Zambon, C.F.; Navaglia, F.; Basso, D.; Rugge, M.; Plebani, M. Helicobacter pylori babA2, cagA, and s1 vacA genes work synergistically in causing intestinal metaplasia. J. Clin. Pathol. 2003, 56, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.H.; Liu, W.Z.; Zhu, H.Y.; Xiao, S.D. Clinical relevance of iceA and babA2 genotypes of Helicobacter pylori in a Shanghai population. Chin. J. Dig. Dis. 2004, 5, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Erzin, Y.; Koksal, V.; Altun, S.; Dobrucali, A.; Aslan, M.; Erdamar, S.; Dirican, A.; Kocazeybek, B. Prevalence of Helicobacter pylori vacA, cagA, cagE, iceA, babA2 genotypes and correlation with clinical outcome in Turkish patients with dyspepsia. Helicobacter 2006, 11, 574–580. [Google Scholar] [CrossRef] [PubMed]
- Con, S.A.; Takeuchi, H.; Nishioka, M.; Morimoto, N.; Sugiura, T.; Yasuda, N.; Con-Wong, R. Clinical relevance of Helicobacter pylori babA2 and babA2/B in Costa Rica and Japan. World J. Gastroenterol. 2010, 16, 474–478. [Google Scholar] [CrossRef]
- Sohrabi, M.; Khashei, R.; Alizadeh, M.; Asl, M.H.; Nejati, M.A.; Dara, M.; Bazargani, A. Low Rate of babA2 Genotype among Iranian Helicobacter pylori Clinical Isolates. J. Clin. Diagn. Res. JCDR 2017, 11, DC32–DC36. [Google Scholar] [CrossRef]
- De Jonge, R.; Pot, R.G.; Loffeld, R.J.; van Vliet, A.H.; Kuipers, E.J.; Kusters, J.G. The functional status of the Helicobacter pylori sabB adhesin gene as a putative marker for disease outcome. Helicobacter 2004, 9, 158–164. [Google Scholar] [CrossRef]
- Sheu, B.S.; Odenbreit, S.; Hung, K.H.; Liu, C.P.; Sheu, S.M.; Yang, H.B.; Wu, J.J. Interaction between host gastric Sialyl-Lewis X and H. pylori SabA enhances H. pylori density in patients lacking gastric Lewis B antigen. Am. J. Gastroenterol. 2006, 101, 36–44. [Google Scholar] [CrossRef]
- Yanai, A.; Maeda, S.; Hikiba, Y.; Shibata, W.; Ohmae, T.; Hirata, Y.; Ogura, K.; Yoshida, H.; Omata, M. Clinical relevance of Helicobacter pylori sabA genotype in Japanese clinical isolates. J. Gastroenterol. Hepatol. 2007, 22, 2228–2232. [Google Scholar] [CrossRef]
- Pakbaz, Z.; Shirazi, M.H.; Ranjbar, R.; Pourmand, M.R.; Khalifeh Gholi, M.; Aliramezani, A.; Vaise Malekshahi, Z. Frequency of sabA Gene in Helicobacter pylori Strains Isolated from Patients in Tehran, Iran. Iran. Red Crescent Med. J. 2013, 15, 767–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, S.; Osaki, T.; Kamiya, S.; Zhang, X.S.; Blaser, M.J. Helicobacter pylori sabA gene is associated with iron deficiency anemia in childhood and adolescence. PLoS ONE 2017, 12, e0184046. [Google Scholar] [CrossRef] [Green Version]
- Ishijima, N.; Suzuki, M.; Ashida, H.; Ichikawa, Y.; Kanegae, Y.; Saito, I.; Borén, T.; Haas, R.; Sasakawa, C.; Mimuro, H. BabA-mediated adherence is a potentiator of the Helicobacter pylori type IV secretion system activity. J. Biol. Chem. 2011, 286, 25256–25264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toller, I.M.; Neelsen, K.J.; Steger, M.; Hartung, M.L.; Hottiger, M.O.; Stucki, M.; Kalali, B.; Gerhard, M.; Sartori, A.A.; Lopes, M.; et al. Carcinogenic bacterial pathogen Helicobacter pylori triggers DNA double-strand breaks and a DNA damage response in its host cells. Proc. Natl. Acad. Sci. USA 2011, 108, 14944–14949. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, P.W.; Snelling, W.J.; Canchaya, C.; Forde, B.M.; Hardie, K.R.; Josenhans, C.; Graham, R.; McMullan, G.; Parkhill, J.; Belda, E.; et al. Comparative genomics and proteomics of Helicobacter mustelae, an ulcerogenic and carcinogenic gastric pathogen. BMC Genom. 2010, 11, 164. [Google Scholar] [CrossRef]
- Cao, D.M.; Lu, Q.F.; Li, S.B.; Wang, J.P.; Chen, Y.L.; Huang, Y.Q.; Bi, H.K. Comparative Genomics of H. pylori and Non-Pylori Helicobacter Species to Identify New Regions Associated with Its Pathogenicity and Adaptability. BioMed Res. Int. 2016, 2016, 6106029. [Google Scholar] [CrossRef] [Green Version]
- Pompaiah, M.; Bartfeld, S. Gastric Organoids: An Emerging Model System to Study Helicobacter pylori Pathogenesis. Curr. Top. Microbiol. Immunol. 2017, 400, 149–168. [Google Scholar] [CrossRef]
- Naz, A.; Awan, F.M.; Obaid, A.; Muhammad, S.A.; Paracha, R.Z.; Ahmad, J.; Ali, A. Identification of putative vaccine candidates against Helicobacter pylori exploiting exoproteome and secretome: A reverse vaccinology based approach. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2015, 32, 280–291. [Google Scholar] [CrossRef]
- Keikha, M.; Eslami, M.; Yousefi, B.; Ghasemian, A.; Karbalaei, M. Potential antigen candidates for subunit vaccine development against Helicobacter pylori infection. J. Cell. Physiol. 2019, 234, 21460–21470. [Google Scholar] [CrossRef]
- Urrutia-Baca, V.H.; Gomez-Flores, R.; De La Garza-Ramos, M.A.; Tamez-Guerra, P.; Lucio-Sauceda, D.G.; Rodríguez-Padilla, M.C. Immunoinformatics Approach to Design a Novel Epitope-Based Oral Vaccine Against Helicobacter pylori. J. Comput. Biol. A J. Comput. Mol. Cell Biol. 2019, 26, 1177–1190. [Google Scholar] [CrossRef] [Green Version]
- Hage, N.; Renshaw, J.G.; Winkler, G.S.; Gellert, P.; Stolnik, S.; Falcone, F.H. Improved expression and purification of the Helicobacter pylori adhesin BabA through the incorporation of a hexa-lysine tag. Protein Expr. Purif. 2015, 106, 25–30. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Adhesin | Reference | Receptor | Receptor Location |
|---|---|---|---|
| BabA | Ilver et al. [7], Bugaytsova et al. [41] | Lewis B antigen | Stomach |
| Ilver et al. [7], Benktander et al. [42] | H type 1 | Stomach | |
| Linden et al. [35] | MUC5AC | Stomach | |
| Linden et al. [43] | MUC1 | Stomach | |
| Benktander et al. [42] | Globo H | Stomach | |
| Benktander et al. [42] | Globo A | Stomach | |
| Walz et al. [44] | MUC5B | Saliva | |
| Linden et al. [36] | MUC7 | Saliva | |
| SabA | Mahdavi et al. [19], Sheu et al. [45] | Sialylated Lewis x | Stomach |
| Benktander et al. [42] | Sialyl-neolactohexaosylceramide | Stomach | |
| Benktander et al. [42] | Sialyl-neolactooctaosylceramide | Stomach | |
| Aspholm-Hurtig et al. [32] | Sialylated Lewis a | Stomach | |
| Aspholm-Hurtig et al. [32] | Sialyl-lactosamine | Stomach | |
| Linden et al. [43] | MUC1 | Stomach | |
| Walz et al. [44] | MUC5B | Saliva |
| Adhesin | Published Year | Country | Total Sample | Association with Disease | Reference |
|---|---|---|---|---|---|
| BabA | 2001 | Korea | 41 | No | Kim et al. [47] |
| 2001 | Italy | 167 | Yes, with diffuse gastritis, peptic ulcer, and duodenitis | Zambon et al. [53] | |
| 2004 | China | 141 | Yes, with duodenal ulcer | Han et al. [54] | |
| 2006 | Turkey | 93 | Yes, with gastric cancer | Erzin et al. [55] | |
| 2010 | Costa Rica | 95 | Yes, with atrophic gastritis | Con et al. [56] | |
| 2010 | Japan | 95 | No | Con et al. [56] | |
| 2011 | Iran | 160 | Yes, with gastric cancer | Talebi Bezmin Abadi et al. [49] | |
| 2017 | Bhutan, Myanmar, Nepal, Bangladesh | 903 | Yes, with peptic ulcer disease | Ansari et al. [13] | |
| 2017 | Iran | 100 | No | Sohrabi et al. [57] | |
| SabA | 2004 | Netherlands | 96 | No | de Jonge et al. [58] |
| 2006 | Colombia and USA | 200 | Yes, with intestinal metaplasia and atrophic gastritis | Yamaoka et al. [30] | |
| 2006 | Taiwan | 145 | No | Sheu et al. [59] | |
| 2007 | Japan | 108 | No | Yanai et al. [60] | |
| 2013 | Iran | 120 | No | Pakbaz et al. [61] | |
| 2017 | Japan | 4 | Yes, with iron deficiency anemia | Kato et al. [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doohan, D.; Rezkitha, Y.A.A.; Waskito, L.A.; Yamaoka, Y.; Miftahussurur, M. Helicobacter pylori BabA–SabA Key Roles in the Adherence Phase: The Synergic Mechanism for Successful Colonization and Disease Development. Toxins 2021, 13, 485. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13070485
Doohan D, Rezkitha YAA, Waskito LA, Yamaoka Y, Miftahussurur M. Helicobacter pylori BabA–SabA Key Roles in the Adherence Phase: The Synergic Mechanism for Successful Colonization and Disease Development. Toxins. 2021; 13(7):485. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13070485
Chicago/Turabian StyleDoohan, Dalla, Yudith Annisa Ayu Rezkitha, Langgeng Agung Waskito, Yoshio Yamaoka, and Muhammad Miftahussurur. 2021. "Helicobacter pylori BabA–SabA Key Roles in the Adherence Phase: The Synergic Mechanism for Successful Colonization and Disease Development" Toxins 13, no. 7: 485. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13070485