Interactions between Filter-Feeding Bivalves and Toxic Diatoms: Influence on the Feeding Behavior of Crassostrea gigas and Pecten maximus and on Toxin Production by Pseudo-nitzschia

Abstract
:1. Introduction
2. Results
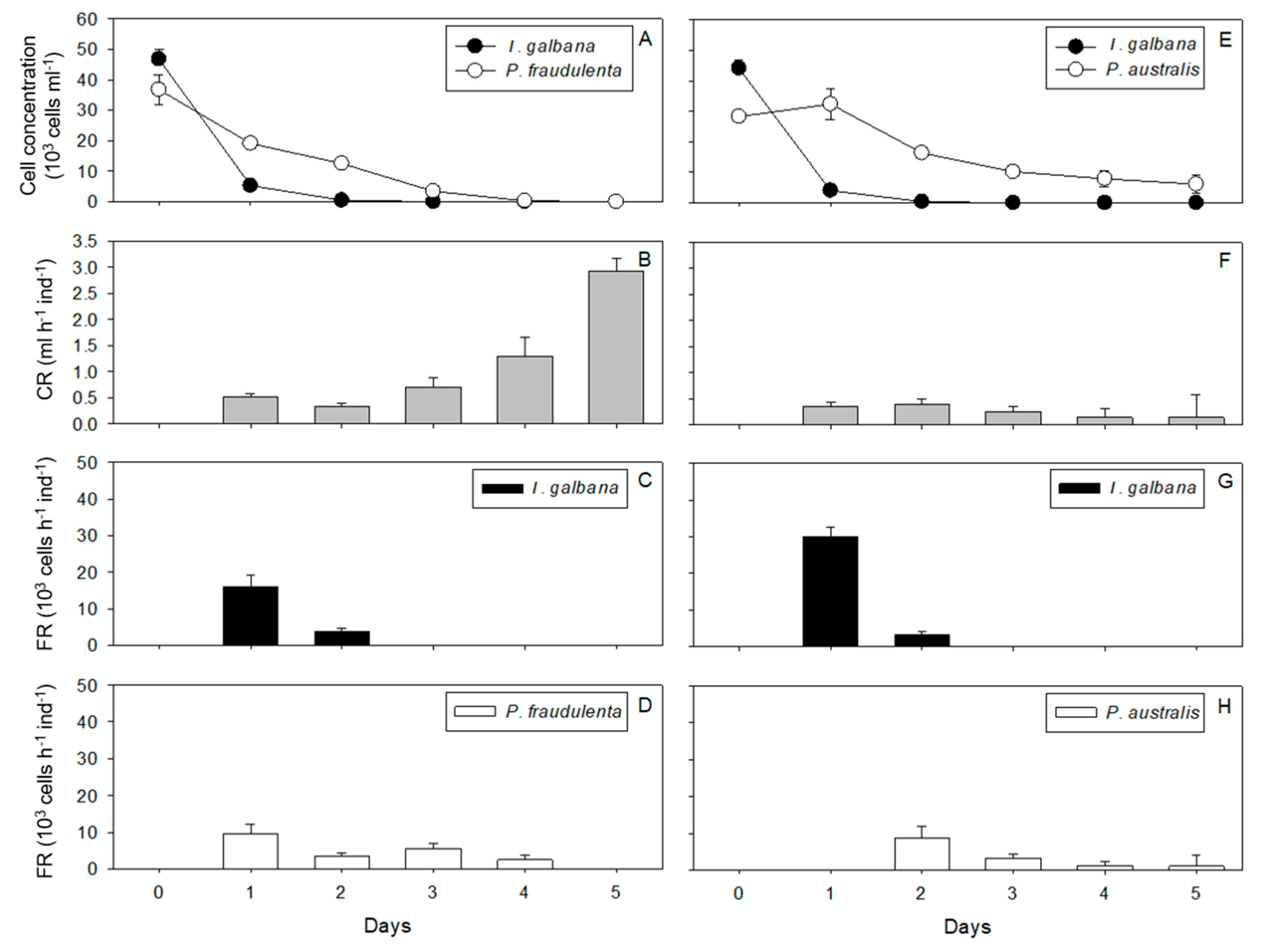
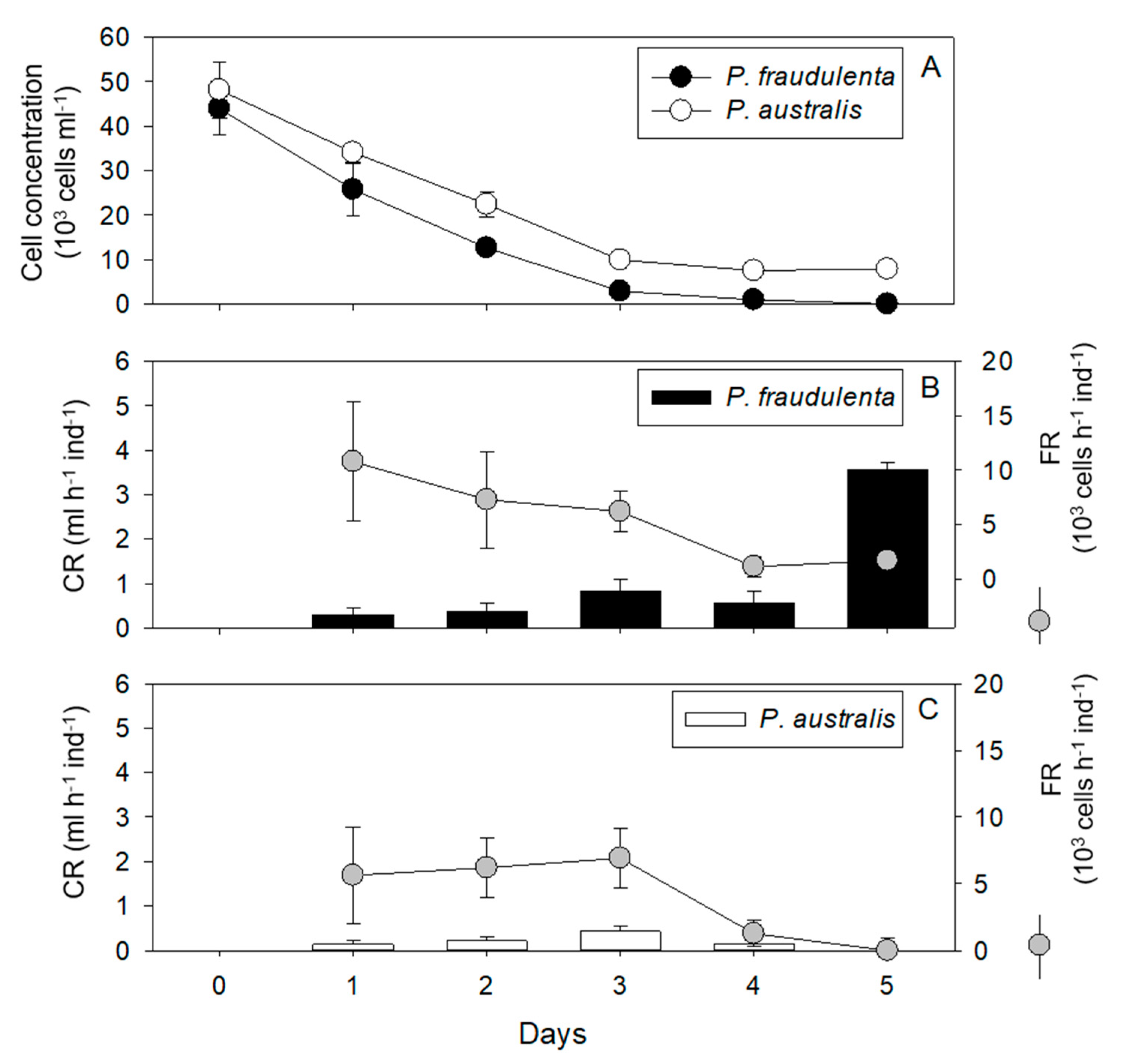
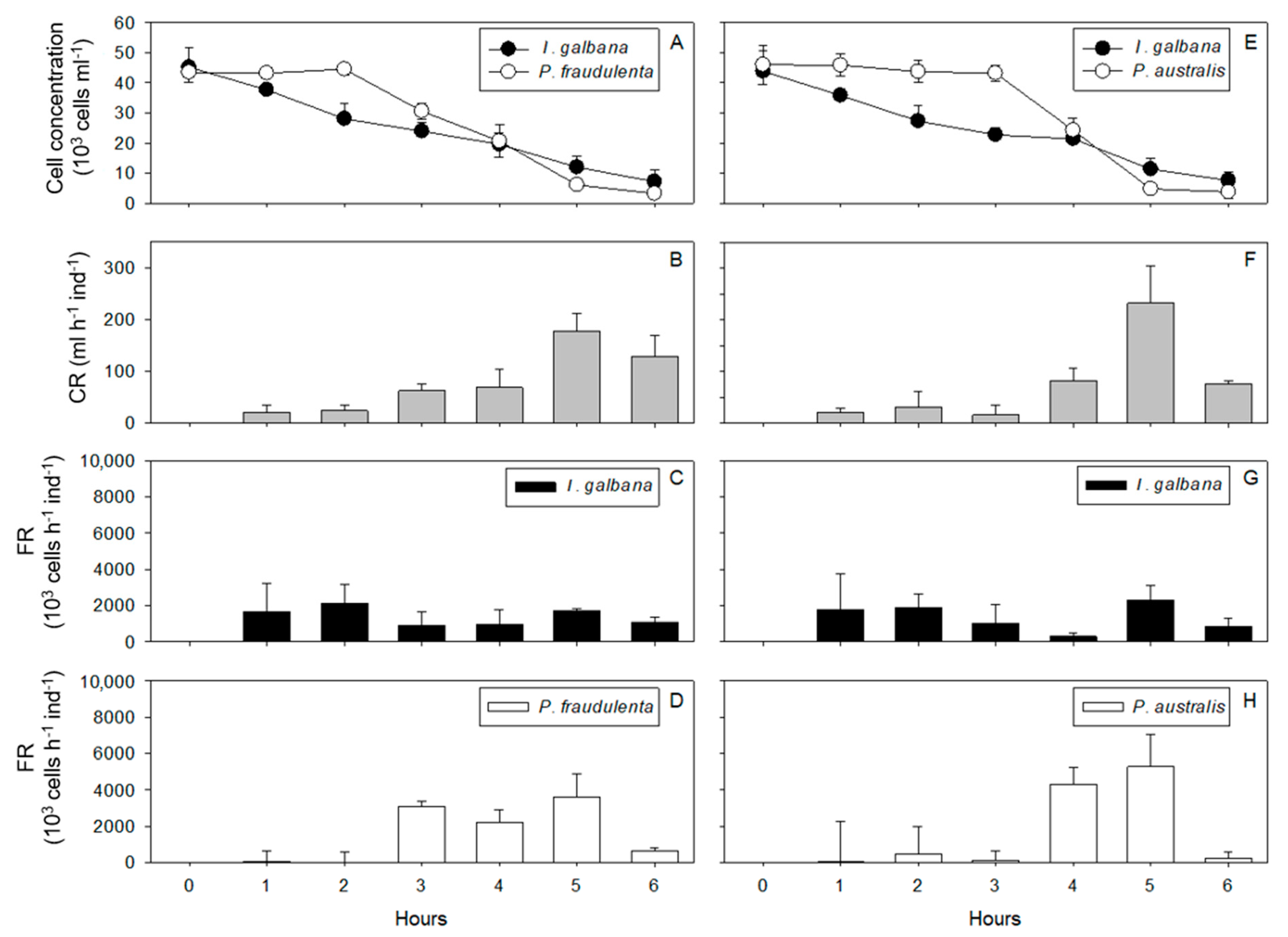
2.1. Cell Concentrations, Clearance Rates, Filtration Rates
2.1.1. Crassostrea gigas
2.1.2. Pecten maximus
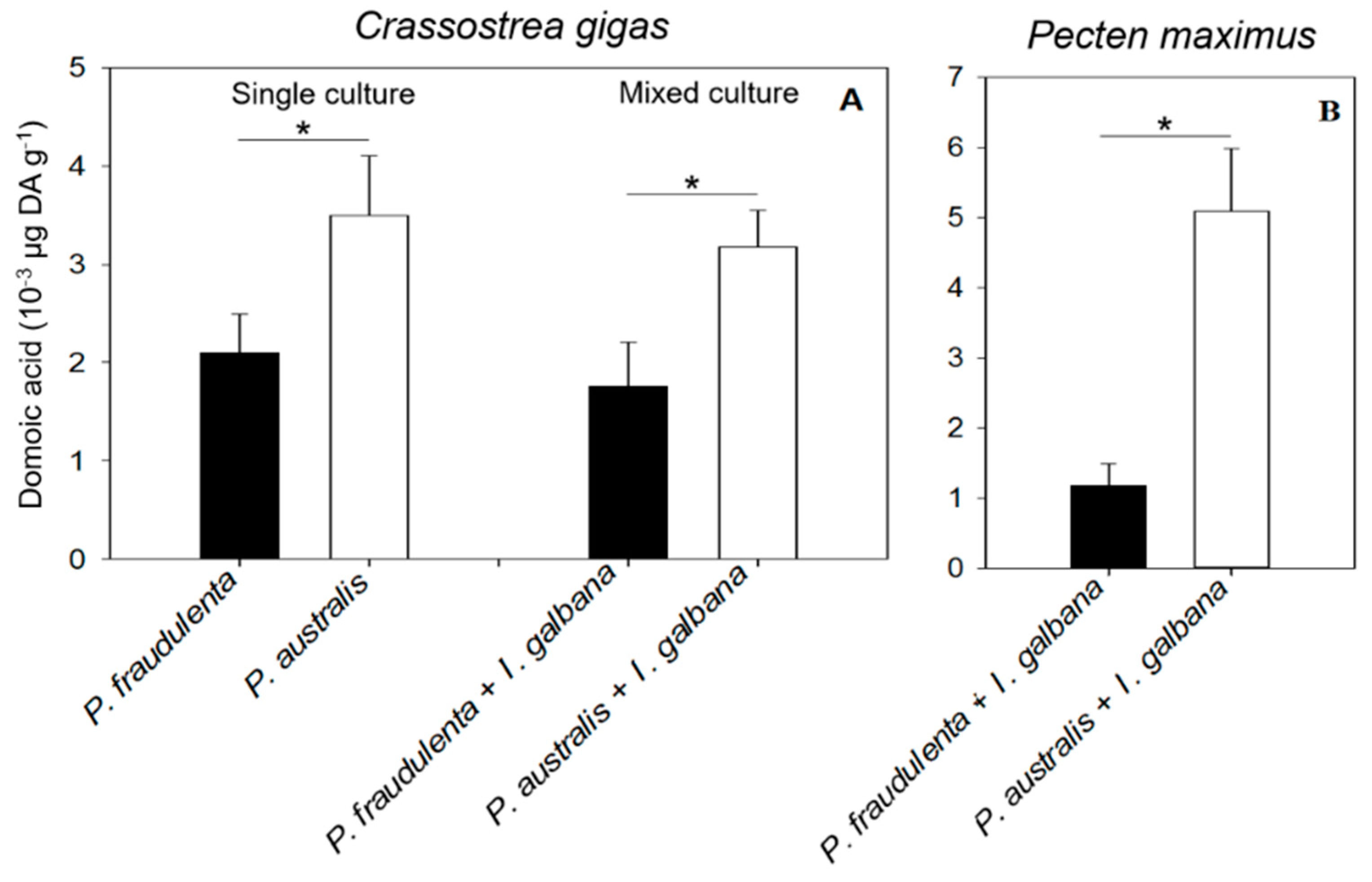
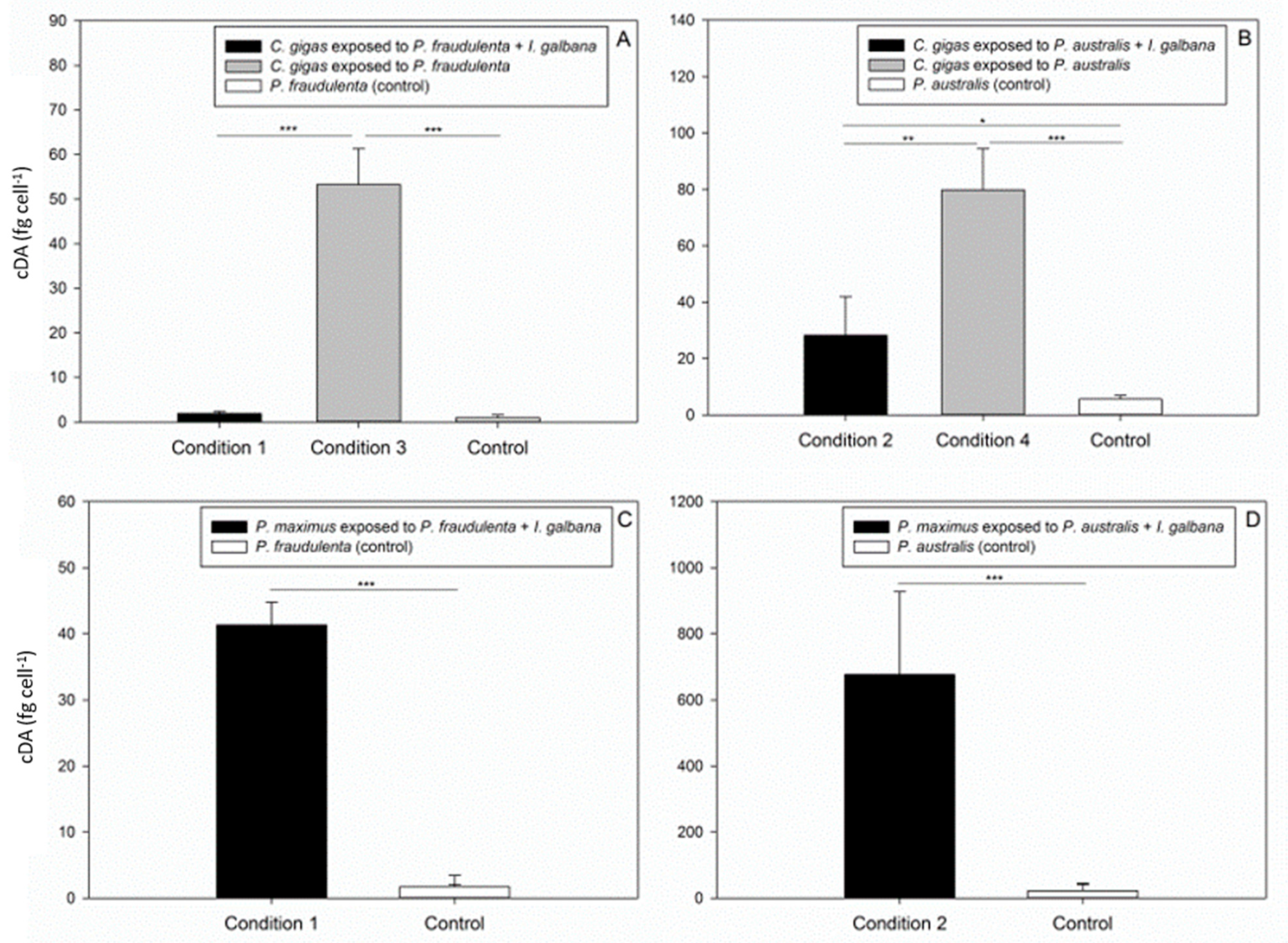
2.2. DA Accumulation in Crassostrea gigas and Pecten maximus
2.3. DA Concentrations in Pseudo-nitzschia Cells (cDA) and in the Medium (dDA), and Nutrient Concentrations
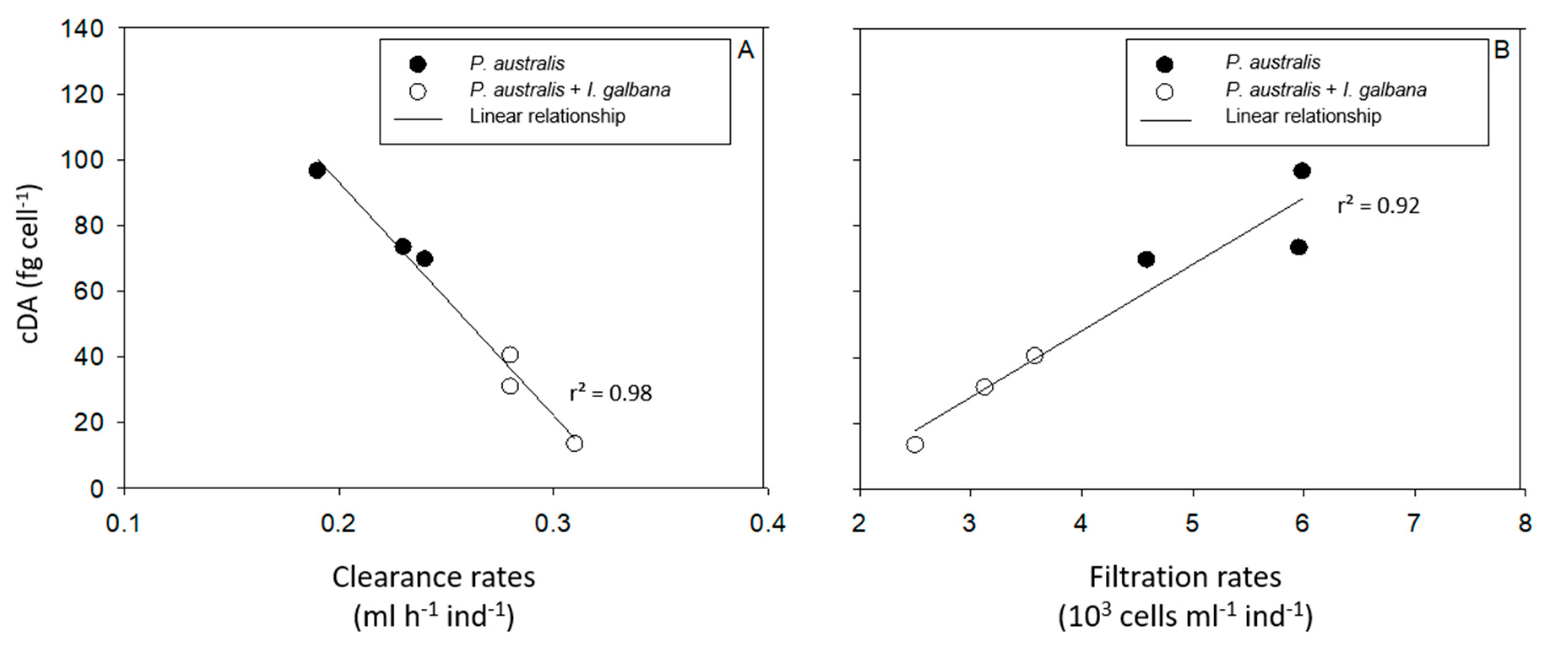
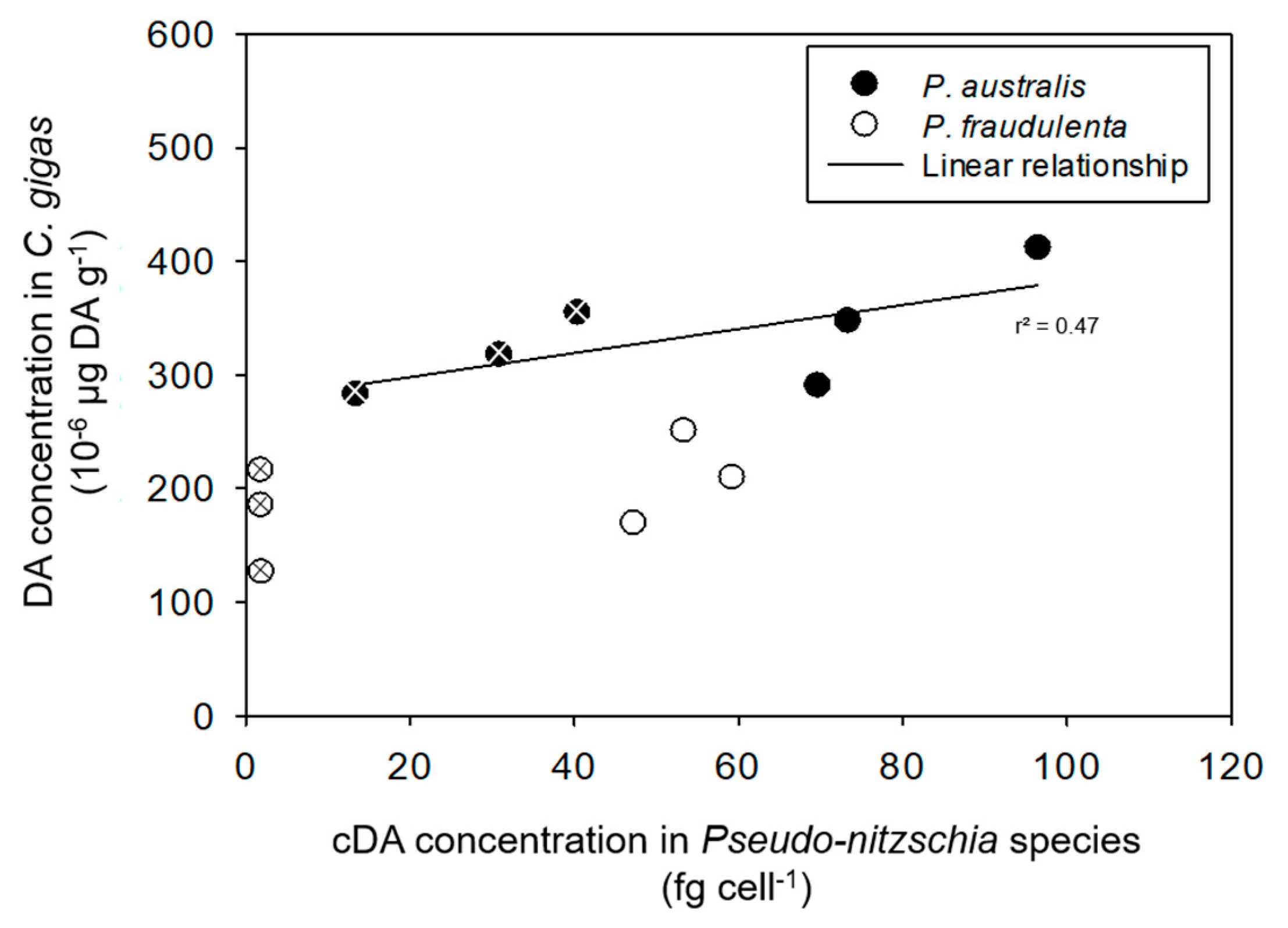
2.4. Potential Relationship between cDA Concentrations in P. australis and P. fraudulenta on the One Hand, and the CR, the FR, and DA Accumulation in Bivalve Species on the Other Hand
3. Discussion
3.1. Feeding Responses of the Bivalves Exposed to Pseudo-nitzschia Species
3.2. DA Accumulation in the Bivalves
3.3. Induction of DA Production in Pseudo-nitzschia in the Presence of Bivalves
4. Conclusions
5. Materials and Methods
5.1. Organisms
5.1.1. Phytoplankton Cultures
5.1.2. Juvenile Filter-Feeding Bivalves
5.2. Experimental Procedure
5.2.1. Experimental Protocols of Contact between Bivalves and Microalgae
5.2.2. Sampling
5.3. Data Analysis
5.3.1. Cell Concentrations, Clearance Rates, Filtration Rates
5.3.2. Dissolved Inorganic Nutrient Analysis
5.3.3. Domoic Acid Analysis
5.3.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bates, S.S.; Hubbard, K.A.; Lundholm, N.; Montresor, M.; Leaw, C.P. Pseudo-nitzschia, Nitzschia, and domoic acid: New research since 2011. Harmful Algae 2018, 79, 3–43. [Google Scholar] [CrossRef]
- Lelong, A.; Hégaret, H.; Soudant, P.; Bates, S.S. Pseudo-nitzschia (Bacillariophyceae) species, domoic acid and amnesic shellfish poisoning: Revisiting previous paradigms. Phycologia 2012, 51, 168–216. [Google Scholar] [CrossRef] [Green Version]
- Trainer, V.L.; Bates, S.S.; Lundholm, N.; Thessen, A.E.; Cochlan, W.P.; Adams, N.G.; Trick, C.G. Pseudo-nitzschia physiological ecology, phylogeny, toxicity, monitoring and impacts on ecosystem health. Harmful Algae 2012, 14, 271–300. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.M.; Pang, J.X.; Huang, C.X.; Lundholm, N.; Teng, S.T.; Li, A.; Li, Y. Two new and non-toxigenic Pseudo-nitzschia species (Bacillariophyceae) from Chinese southeast coastal waters. J. Phycol. 2020, 57, 335–344. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. Available online: www.algaebase.org (accessed on 13 January 2021).
- Bates, S.S.; Bird, C.J.; de Freitas, A.S.W.; Foxall, R.; Gilgan, M.; Hanic, L.A.; Johnson, G.R.; McCulloch, A.W.; Odense, P.; Pocklington, R.; et al. Pennate diatom Nitzschia pungens as the primary source of domoic acid, a toxin in shellfish from eastern Prince Edward Island, Canada. Can. J. Fish. Aquat. Sci. 1989, 46, 1203–1215. [Google Scholar] [CrossRef]
- Visciano, P.; Schirone, M.; Berti, M.; Milandri, A.; Tofalo, R.; Suzzi, G. Marine biotoxins: Occurrence, toxicity, regulatory limits and reference methods. Front. Microbiol. 2016, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delegrange, A.; Lefebvre, A.; Gohin, F.; Courcot, L.; Vincent, D. Pseudo-nitzschia sp. diversity and seasonality in the southern North Sea, domoic acid levels and associated phytoplankton communities. Estuar. Coast. Shelf Sci. 2018, 214, 194–206. [Google Scholar] [CrossRef] [Green Version]
- Geuer, J.K.; Trimborn, S.; Koch, F.; Brenneis, T.; Krock, B.; Koch, B.P. Dissolved domoic acid does not improve growth rates and iron content in iron-stressed Pseudo-nitzschia subcurvata. Front. Mar. Sci. 2020, 7, 1–13. [Google Scholar] [CrossRef]
- Umhau, B.P.; Benitez-Nelson, C.R.; Anderson, C.R.; McCabe, K.; Burrell, C. A time series of water column distributions and sinking particle flux of Pseudo-nitzschia and domoic acid in the Santa Barbara Basin, California. Toxins 2018, 10, 480. [Google Scholar] [CrossRef] [Green Version]
- De Rijcke, M.; Vandegehuchte, M.B.; Vanden Bussche, J.; Nevejan, N.; Vanhaecke, L.; De Schamphelaere, K.A.C.; Janssen, C.R. Common European harmful algal blooms affect the viability and innate immune responses of Mytilus edulis larvae. Fish Shellfish Immunol. 2015, 47, 175–181. [Google Scholar] [CrossRef]
- Liu, H.; Kelly, M.S.; Campbell, D.A.; Dong, S.L.; Zhu, J.X.; Wang, S.F. Exposure to domoic acid affects larval development of king scallop Pecten maximus (Linnaeus, 1758). Aquat. Toxicol. 2007, 81, 152–158. [Google Scholar] [CrossRef]
- Panlilio, J.M.; Aluru, N.; Hahn, M.E. Developmental neurotoxicity of the harmful algal bloom toxin domoic acid: Cellular and molecular mechanisms underlying altered behavior in the zebrafish model. Environ. Health Perspect. 2020, 128, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Tester, P.A.; Pan, Y.; Doucette, G.J. Accumulation of domoic acid activity in copepods. Harmful Algal Bloom. 2000, 418–421. Available online: https://www.researchgate.net/publication/258178061_Accumulation_of_domoic_acid_activity_in_copepods (accessed on 11 June 2021).
- Lincoln, J.A.; Turner, J.T.; Bates, S.S.; Léger, C.; Gauthier, D.A. Feeding, egg production, and egg hatching success of the copepods Acartia tonsa and Temora longicornis on diets of the toxic diatom Pseudo-nitzschia multiseries and the non-toxic diatom Pseudo-nitzschia pungens. Hydrobiologia 2001, 453–454, 107–120. [Google Scholar] [CrossRef]
- Maneiro, I.; Iglesias, P.; Guisande, C.; Riveiro, I.; Barreiro, A.; Zervoudaki, S.; Granéli, E. Fate of domoic acid ingested by the copepod Acartia clausi. Mar. Biol. 2005, 148, 123–130. [Google Scholar] [CrossRef]
- Leandro, L.F.; Teegarden, G.J.; Roth, P.B.; Wang, Z.; Doucette, G.J. The copepod Calanus finmarchicus: A potential vector for trophic transfer of the marine algal biotoxin, domoic acid. J. Exp. Mar. Biol. Ecol. 2010, 382, 88–95. [Google Scholar] [CrossRef]
- Tammilehto, A.; Nielsen, T.G.; Krock, B.; Møller, E.F.; Lundholm, N. Induction of domoic acid production in the toxic diatom Pseudo-nitzschia seriata by calanoid copepods. Aquat. Toxicol. 2015, 159, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Tammilehto, A.; Nielsen, T.G.; Krock, B.; Møller, E.F.; Lundholm, N. Calanus spp.-Vectors for the biotoxin, domoic acid, in the Arctic marine ecosystem? Harmful Algae 2012, 20, 165–174. [Google Scholar] [CrossRef]
- Harðardóttir, S.; Pančić, M.; Tammilehto, A.; Krock, B.; Møller, E.F.; Nielsen, T.G.; Lundholm, N. Dangerous relations in the Arctic marine food web: Interactions between toxin producing Pseudo-nitzschia diatoms and Calanus copepodites. Mar. Drugs 2015, 13, 3809–3835. [Google Scholar] [CrossRef] [Green Version]
- Lundholm, N.; Krock, B.; John, U.; Skov, J.; Cheng, J.; Pančić, M.; Wohlrab, S.; Rigby, K.; Nielsen, T.G.; Selander, E.; et al. Induction of domoic acid production in diatoms—Types of grazers and diatoms are important. Harmful Algae 2018, 79, 64–73. [Google Scholar] [CrossRef]
- Harðardóttir, S.; Krock, B.; Wohlrab, S.; John, U.; Nielsen, T.G.; Lundholm, N. Can domoic acid affect escape response in copepods? Harmful Algae 2018, 79, 50–52. [Google Scholar] [CrossRef] [Green Version]
- Mafra, L.L.; Bricelj, V.M.; Ouellette, C.; Bates, S.S. Feeding mechanics as the basis for differential uptake of the neurotoxin domoic acid by oysters, Crassostrea virginica, and mussels, Mytilus edulis. Aquat. Toxicol. 2010, 97, 160–171. [Google Scholar] [CrossRef]
- Mafra, L.L.; Bricelj, V.M.; Ouellette, C.; Léger, C.; Bates, S.S. Mechanisms contributing to low domoic acid uptake by oysters feeding on Pseudo-nitzschia cells. I. Filtration and pseudofeces production. Aquat. Biol. 2009, 6, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Mafra, L.L.; Bricelj, V.M.; Ward, J.E. Mechanisms contributing to low domoic acid uptake by oysters feeding on Pseudo-nitzschia cells. II. Selective rejection. Aquat. Biol. 2009, 6, 213–226. [Google Scholar] [CrossRef]
- Thessen, A.E.; Soniat, T.M.; Dortch, Q.; Doucette, G.J. Crassostrea virginica grazing on toxic and non-toxic diatoms. Toxicon 2010, 55, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Bardouil, M.; Bohec, M.; Bougrier, S.; Lassus, P.; Truquet, P. Feeding responses of Crassostrea gigas (Thunberg) to inclusion of different proportions of toxic dinoflagellates in their diet. Oceanol. Acta 1996, 19, 177–182. [Google Scholar]
- Bricelj, V.M.; MacQuarrie, S.P.; Schaffner, R.A. Differential effects of Aureococcus anophagefferens isolates (“brown tide”) in unialgal and mixed suspensions on bivalve feeding. Mar. Biol. 2001, 139, 605–615. [Google Scholar] [CrossRef]
- Contreras, A.M.; Marsden, I.D.; Munro, M.H.G. Effects of short-term exposure to paralytic shellfish toxins on clearance rates and toxin uptake in five species of New Zealand bivalve. Mar. Freshw. Res. 2012, 63, 166–174. [Google Scholar] [CrossRef]
- Gainey, L.F.; Shumway, S.E. The physiological effect of Aureococcus anophagefferens (“brown tide”) on the lateral cilia of bivalve mollusks. Biol. Bull. 1991, 181, 298–306. [Google Scholar] [CrossRef] [Green Version]
- Shumway, S.E.; Burkholder, J.A.M.; Springer, J. Effects of the estuarine dinoflagellate Pfiesteria shumwayae (Dinophyceae) on survival and grazing activity of several shellfish species. Harmful Algae 2006, 5, 442–458. [Google Scholar] [CrossRef]
- May, S.P.; Burkholder, J.A.M.; Shumway, S.E.; Hégaret, H.; Wikfors, G.H.; Frank, D. Effects of the toxic dinoflagellate Alexandrium monilatum on survival, grazing and behavioral response of three ecologically important bivalve molluscs. Harmful Algae 2010, 9, 281–293. [Google Scholar] [CrossRef]
- Navarro, J.M.; Widdows, J.; Chaparro, O.R.; Ortíz, A.; Mellado, C.; Villanueva, P.A. Pre-ingestive selection capacity and endoscopic analysis in the sympatric bivalves Mulinia edulis and Mytilus chilensis exposed to diets containing toxic and non-toxic dinoflagellates. PLoS ONE 2018, 13, 0193370. [Google Scholar] [CrossRef] [Green Version]
- Hégaret, H.; Wikfors, G.H.; Shumway, S.E. Diverse feeding responses of five species of bivalve mollusc when exposed to three species of harmful algae. J. Shellfish Res. 2007, 26, 549–559. [Google Scholar] [CrossRef]
- Selander, E.; Cervin, G.; Pavia, H. Effects of nitrate and phosphate on grazer-induced toxin production in Alexandrium minutum. Limnol. Oceanogr. 2008, 53, 523–530. [Google Scholar] [CrossRef]
- Selander, E.; Fagerberg, T.; Wohlra, S.; Paviae, H. Fight and flight in dinoflagellates? Kinetics of simultaneous grazer-induced responses in Alexandrium tamarense. Limnol. Oceanogr. 2012, 57, 58–64. [Google Scholar] [CrossRef]
- Selander, E.; Thor, P.; Toth, G.; Pavia, H. Copepods induce paralytic shellfish toxin production in marine dinoflagellates. Proc. R. Soc. B Biol. Sci. 2006, 273, 1673–1680. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.X.; Dong, H.C.; Lundholm, N.; Teng, S.T.; Zheng, G.C.; Tan, Z.J.; Lim, P.T.; Li, Y. Species composition and toxicity of the genus Pseudo-nitzschia in Taiwan Strait, including P. chiniana sp. nov. and P. qiana sp. nov. Harmful Algae 2019, 84, 195–209. [Google Scholar] [CrossRef]
- Husson, B.; Hernández-Fariñas, T.; Le Gendre, R.; Schapira, M.; Chapelle, A. Two decades of Pseudo-nitzschia spp. blooms and king scallop (Pecten maximus) contamination by domoic acid along the French Atlantic and English Channel coasts: Seasonal dynamics, spatial heterogeneity and interannual variability. Harmful Algae 2016, 51, 26–39. [Google Scholar] [CrossRef] [Green Version]
- Caruana, A.M.N.; Ayache, N.; Raimbault, V.; Rétho, M.; Hervé, F.; Bilien, G.; Amzil, Z.; Chomérat, N. Direct evidence for toxin production by Pseudo-nitzschia plurisecta (Bacillariophyceae) and extension of its distribution area. Eur. J. Phycol. 2019, 54, 585–594. [Google Scholar] [CrossRef]
- Downes-Tettmar, N.; Rowland, S.; Widdicombe, C.; Woodward, M.; Llewellyn, C. Seasonal variation in Pseudo-nitzschia spp. and domoic acid in the western English Channel. Cont. Shelf Res. 2013, 53, 40–49. [Google Scholar] [CrossRef]
- Klein, C.; Claquin, P.; Bouchart, V.; Le Roy, B.; Véron, B. Dynamics of Pseudo-nitzschia spp. and domoic acid production in a macrotidal ecosystem of the eastern English Channel (Normandy, France). Harmful Algae 2010, 9, 218–226. [Google Scholar] [CrossRef]
- Nezan, E.; Antoine, E.; Fiant, L.; Billard, C. Identification of Pseudo-nitzschia australis and P. multiseries in the Bay of Seine. Was there a relation to presence of domoic acid in King scallops in autumn 2004? Harmful Algae News 2006, 31, 1–12. [Google Scholar]
- Thorel, M.; Claquin, P.; Schapira, M.; Le Gendre, R.; Riou, P.; Goux, D.; Le Roy, B.; Raimbault, V.; Deton-Cabanillas, A.-F.; Bazin, P.; et al. Nutrient ratios influence variability in Pseudo-nitzschia species diversity and particulate domoic acid production in the Bay of Seine (France). Harmful Algae 2017, 68, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Lema, K.A.; Latimier, M.; Nezan, E.; Fauchot, J.; Le Gac, M. Inter and intra-specific growth and domoic acid production in relation to nutrient ratios and concentrations in Pseudo-nitzschia: Phosphate an important factor. Harmful Algae 2017, 64, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauvey, A.; Claquin, P.; Le Roy, B.; Le Gac, M.; Fauchot, J. Differential influence of life cycle on growth and toxin production of three Pseudo-nitzschia species (Bacillariophyceae). J. Phycol. 2019, 55, 1126–1139. [Google Scholar] [CrossRef] [PubMed]
- Belin, C.; Amzil, Z. Phycotoxin monitoring in France: Risk-based strategy and main results (2006–2008). In Proceedings of the 7th International Conference on Molluscan Shellfish Safety, Nantes, France, 14–19 June 2009; Lassus, P., Ed.; Ifremer: Nantes, France, 2009; pp. 149–156. [Google Scholar]
- Amzil, Z.; Fresnel, J.; Le Gal, D.; Billard, C. Domoic acid accumulation in french shellfish in relation to toxic species of Pseudo-nitzschia multiseries and P. pseudodelicatissima. Toxicon 2001, 39, 1245–1251. [Google Scholar] [CrossRef]
- Rosa, A.M.; Ward, J.E.; Shumway, S.E. Selective capture and ingestion of particles by suspension-feeding bivalve molluscs: A review. J. Shellfish Res. 2018, 37, 727–746. [Google Scholar] [CrossRef]
- Ward, J.E.; Shumway, S.E. Separating the grain from the chaff: Particle selection in suspension- and deposit-feeding bivalves. J. Exp. Mar. Biol. Ecol. 2004, 300, 83–130. [Google Scholar] [CrossRef]
- Shumway, S.E.; Cucci, T.L.; Newell, R.C.; Selvin, R.C.; Guillard, R.R.L.; Yentsch, C.M. Flow cytometry: A new method for characterization of differential ingestion, digestion and egestion by suspension feeders. Mar. Ecol. Prog. Ser. 1985, 24, 201–204. [Google Scholar]
- Leverone, J.R.; Shumway, S.E.; Blake, N.J. Comparative effects of the toxic dinoflagellate, Karenia brevis, on bivalve molluscs from Florida, USA. Toxicon 2007, 49, 634–645. [Google Scholar] [CrossRef] [PubMed]
- Pousse, E.; Flye-Sainte-Marie, J.; Alunno-Bruscia, M.; Hégaret, H. Sources of paralytic shellfish toxin accumulation variability in the Pacific oyster Crassostrea gigas. Toxicon 2018, 144, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Shumway, S.E.; Barter, J.; Sherman-Caswell, S. Auditing the impact of toxic algal blooms on oysters. Environ. Audit. 1990, 2, 41–56. [Google Scholar]
- Borcier, E.; Morvezen, R.; Boudry, P.; Miner, P.; Charrier, G.; Laroche, J.; Hégaret, H. Effects of bioactive extracellular compounds and paralytic shellfish toxins produced by Alexandrium minutum on growth and behaviour of juvenile great scallops Pecten maximus. Aquat. Toxicol. 2017, 184, 142–154. [Google Scholar] [CrossRef] [Green Version]
- Douglas, D.J.; Kenchington, E.R.; Bird, C.J.; Pocklington, R.; Bradford, B.; Silvert, W. Accumulation of domoic acid by the sea scallop (Placopecten magellanicus) fed cultured cells of toxic Pseudo-nitzschia multiseries. Can. J. Fish. Aquat. Sci. 1997, 54, 907–913. [Google Scholar] [CrossRef]
- Jones, T.O.; Whyte, J.N.C.; Townsend, L.D.; Gintherb, N.G.; Iwamaa, G.K. Effects of domoic acid on haemolymph pH, PCO2, and PO in the Pacific oyster, Crassostrea gigas and the California mussel, Mytilus californianus. Aquat. Toxicol. 1995, 31, 43–55. [Google Scholar] [CrossRef]
- Mafra, L.L.; Bricelj, V.M.; Fennel, K. Domoic acid uptake and elimination kinetics in oysters and mussels in relation to body size and anatomical distribution of toxin. Aquat. Toxicol. 2010, 100, 17–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beninger, P.G.; Ward, E.; MacDonald, B.A.; Thompson, R.J. Gill function and particle transport in Placopecten magellanicus (Mollusca: Bivalvia) as revealed using video endoscopy. Mar. Biol. 1992, 114, 281–288. [Google Scholar] [CrossRef]
- Stewart, J.E.; Marks, L.J.; Gilgan, M.W.; Pfeiffer, E.; Zwicker, B.M. Microbial utilization of the neurotoxin domoic acid: Blue mussels (Mytilus edulis) and soft shell clams (Mya arenaria) as sources of the microorganisms. Can. J. Microbiol. 1998, 44, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Bates, S.; Garrison, D.; Horner, R. Bloom dynamics and physiology producing Pseudo-nitzschia species. NATO ASI Ser. G Ecol. Sci. 1998, 41, 267–292. [Google Scholar]
- Fehling, J.; Davidson, K.; Bolch, C.J.; Bates, S.S. Growth and domoic acid production by Pseudo-nitzschia seriata (Bacillariophyceae) under phosphate and silicate limitation. J. Phycol. 2004, 40, 674–683. [Google Scholar] [CrossRef]
- Hagström, J.A.; Granéli, E.; Moreira, M.O.P.; Odebrecht, C. Domoic acid production and elemental composition of two Pseudo-nitzschia multiseries strains, from the NW and SW Atlantic Ocean, growing in phosphorus or nitrogen-limited chemostat cultures. J. Plankton Res. 2011, 33, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Kudela, R.; Roberts, A.; Armstrong, M. Laboratory analyses of nutrient stress and toxin production in Pseudo-nitzschia spp. from Monterey Bay, California. Harmful Algae 2002, 136–138. Available online: https://people.ucsc.edu/~kudela/reprints/Kudela_XHAB2_2003.pdf (accessed on 11 June 2021).
- Pan, Y.; Subba Rao, D.V.; Mann, K.H.; Brown, R.G.; Pocklington, R. Effects of silicate limitation on production of domoic acid, a neurotoxin, by the diatom Pseudo-nitzschia multiseries. I. Batch culture studies. Mar. Ecol. Prog. Ser. 1996, 131, 225–233. [Google Scholar] [CrossRef]
- Pan, Y.; Subba Rao, D.V.; Mann, K.H.; Li, W.K.W.; Harrison, W.G. Effects of silicate limitation on production of domoic acid, a neurotoxin, by the diatom Pseudo-nitzschia multiseries. II. Continuous culture studies. Mar. Ecol. Prog. Ser. 1996, 131, 235–243. [Google Scholar] [CrossRef]
- Senft-Batoh, C.D.; Dam, H.G.; Shumway, S.E.; Wikfors, G.H. A multi-phylum study of grazer-induced paralytic shellfish toxin production in the dinoflagellate Alexandrium fundyense: A new perspective on control of algal toxicity. Harmful Algae 2015, 44, 20–31. [Google Scholar] [CrossRef]
- Miesner, A.K.; Lundholm, N.; Krock, B.; Nielsen, T.G. The effect of Pseudo-nitzschia seriata on grazing and fecundity of Calanus finmarchicus and Calanus glacialis. J. Plankton Res. 2016, 38, 564–574. [Google Scholar] [CrossRef] [Green Version]
- Selander, E.; Kubanek, J.; Hamberg, M.; Andersson, M.X.; Cervin, G.; Pavia, H. Predator lipids induce paralytic shellfish toxins in bloom-forming algae. Proc. Natl. Acad. Sci. USA 2015, 112, 6395–6400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wohlrab, S.; Iversen, M.H.; John, U. A Molecular and co-evolutionary context for grazer induced toxin production in Alexandrium tamarense. PLoS ONE 2010, 5, e15039. [Google Scholar] [CrossRef] [PubMed]
- Grebner, W.; Berglund, E.C.; Berggren, F.; Eklund, J.; Harðadóttir, S.; Andersson, M.X.; Selander, E. Induction of defensive traits in marine plankton—new copepodamide structures. Limnol. Oceanogr. 2018, 64, 820–831. [Google Scholar] [CrossRef]
- Keller, M.D.; Selvin, R.C.; Claus, W.; Guillard, R.R.L. Media for the culture of oceanic ultraphytoplankton. J. Phycol. 1987, 23, 633–638. [Google Scholar] [CrossRef]
- Lundholm, N.; Daugbjerg, N.; Moestrup, Ø. Phylogeny of the Bacillariaceae with emphasis on the genus Pseudo-nitzschia (Bacillariophyceae) based on partial LSU RDNA. Eur. J. Phycol. 2002, 37, 115–134. [Google Scholar] [CrossRef] [Green Version]
- Coughlan, J. The estimation of filtering rate from the clearance of suspensions. Mar. Biol. 1969, 2, 356–358. [Google Scholar] [CrossRef]
- Aminot, A.; Kérouel, R. Dosage Automatique Des Nutriments Dans Les Eaux Marines: Méthodes en Flux Continu; Editions Quae: Paris, France, 2007. [Google Scholar]
- Garthwaite, L.; Ross, K.M.; Miles, C.O.; Hansen, R.P.; Foster, D.; Wilkins, A.L.; Towers, N.R. Polyclonal antibodies to domoic acid, and their use in immunoassays for domoic acid in sea water and shellfish. Nat. Toxins 1998, 6, 93–104. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. gigas | P. maximus | |||
|---|---|---|---|---|
| Experimental data | Duration | 5 days | 6 h | |
| Volume | 250 mL | 220 mL | ||
| Number of individuals per condition | 20 C. gigas | 1 P. maximus | ||
| Average size of individuals | 1.0 ± 0.15 cm | 4.10 ± 0.75 cm | ||
| Average weight of individuals | 18 ± 4 mg | 1.0 ± 0.2 g | ||
| Experimental conditions | Bivalves exposed to a mixed culture of microalgae | Condition 1 | 38 × 103 cells mL−1 P. fraudulenta 47 × 103 cells mL−1 I. galbana | 43 × 103 cells mL−1 P. fraudulenta 45 × 103 cells mL−1 I. galbana |
| Condition 2 | 28 × 103 cells mL−1 P. australis 44 × 103 cells mL−1 I. galbana | 46 × 103 cells mL−1 P. australis 44 × 103 cells mL−1 I. galbana | ||
| Bivalves exposed to a single culture of Pseudo-nitzschia | Condition 3 | 42 × 103 cells mL−1 P. fraudulenta | - | |
| Condition 4 | 48 × 103 cells mL−1 P. australis | - | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sauvey, A.; Denis, F.; Hégaret, H.; Le Roy, B.; Lelong, C.; Jolly, O.; Pavie, M.; Fauchot, J. Interactions between Filter-Feeding Bivalves and Toxic Diatoms: Influence on the Feeding Behavior of Crassostrea gigas and Pecten maximus and on Toxin Production by Pseudo-nitzschia. Toxins 2021, 13, 577. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080577
Sauvey A, Denis F, Hégaret H, Le Roy B, Lelong C, Jolly O, Pavie M, Fauchot J. Interactions between Filter-Feeding Bivalves and Toxic Diatoms: Influence on the Feeding Behavior of Crassostrea gigas and Pecten maximus and on Toxin Production by Pseudo-nitzschia. Toxins. 2021; 13(8):577. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080577
Chicago/Turabian StyleSauvey, Aurore, Françoise Denis, Hélène Hégaret, Bertrand Le Roy, Christophe Lelong, Orianne Jolly, Marie Pavie, and Juliette Fauchot. 2021. "Interactions between Filter-Feeding Bivalves and Toxic Diatoms: Influence on the Feeding Behavior of Crassostrea gigas and Pecten maximus and on Toxin Production by Pseudo-nitzschia" Toxins 13, no. 8: 577. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080577