Inhibition of Diarrheal Shellfish Toxins Accumulation in the Mussel Perna viridis by Curcumin and Underlying Mechanisms


Abstract
:1. Introduction
2. Results
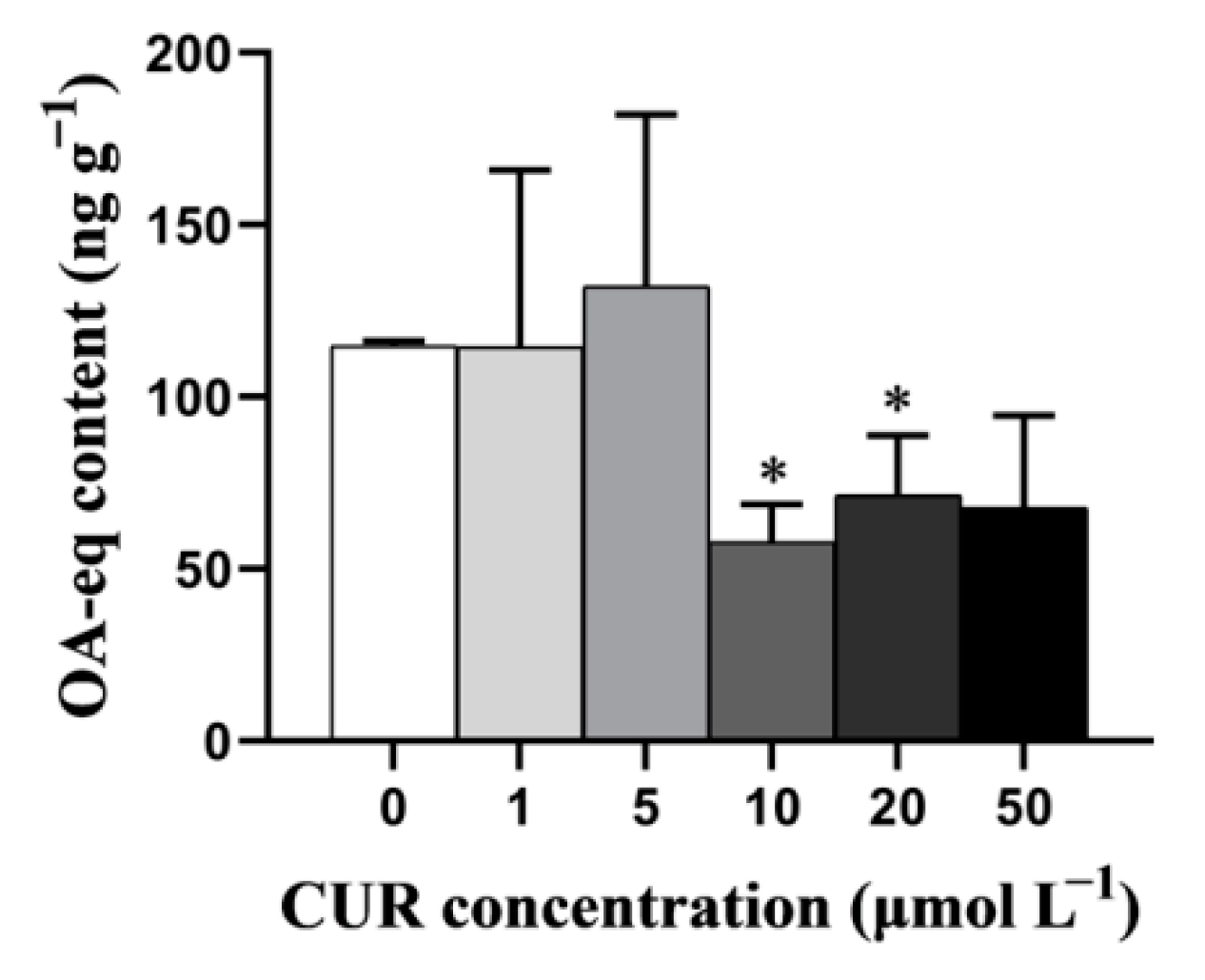
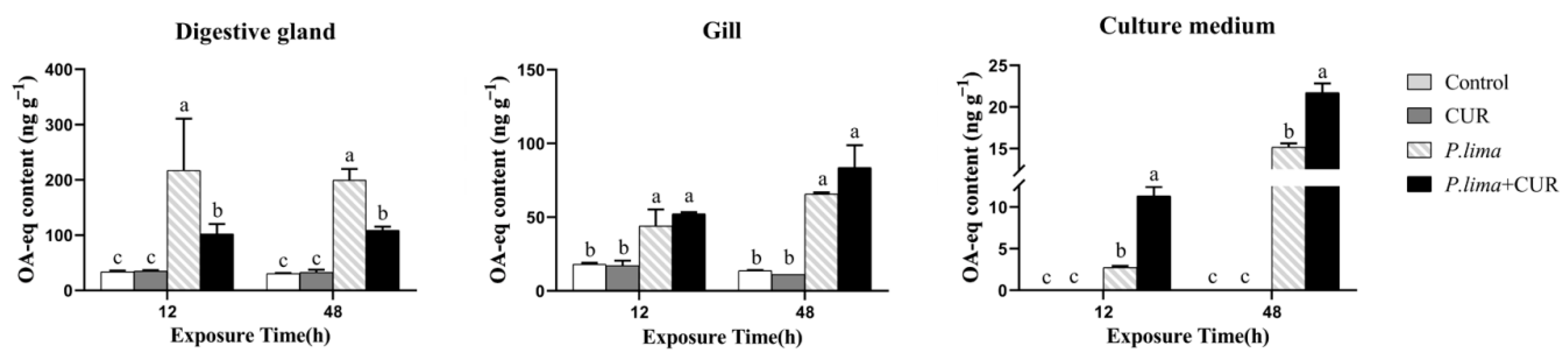
2.1. CUR Decreases the Accumulation of DSTs in the Digestive Gland but Not Gills of Mussels
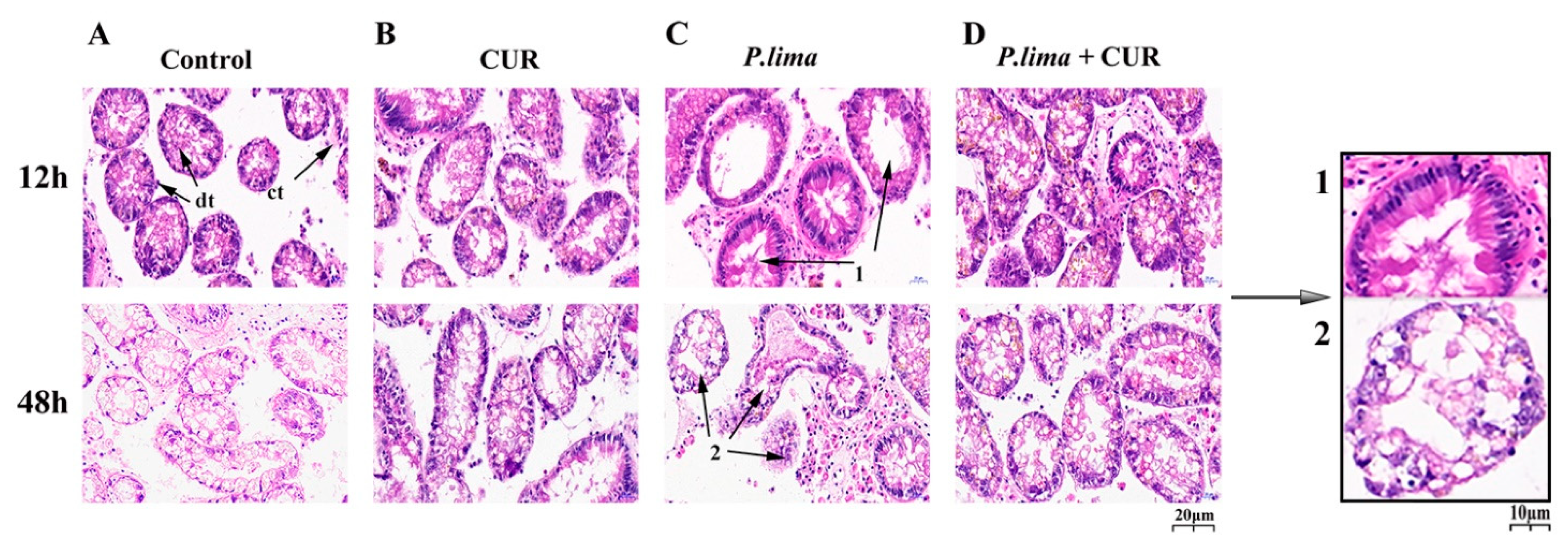
2.2. CUR Reduced Damage to the Digestive Gland in the P. lima-Exposed Mussels
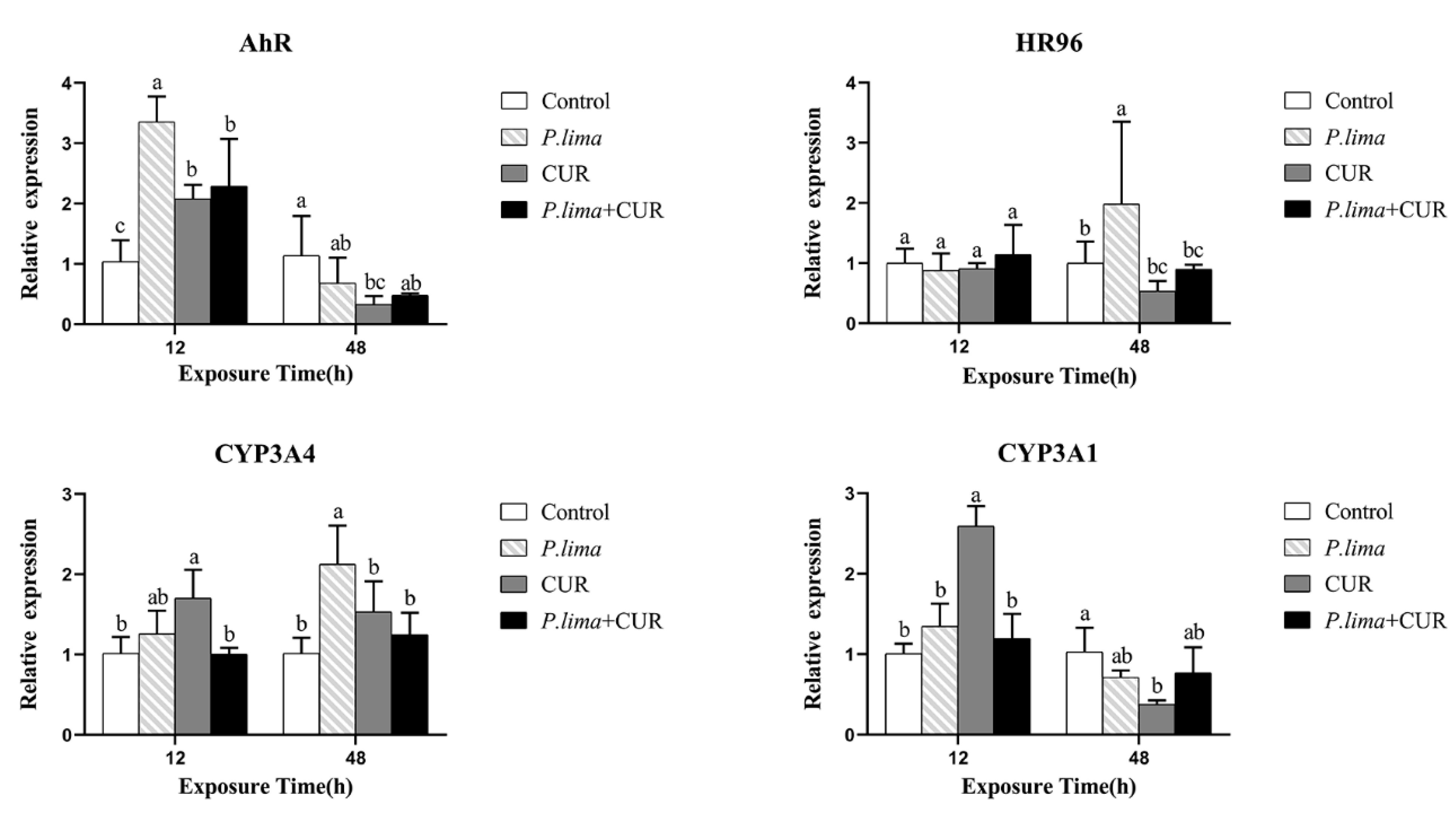
2.3. CUR Might Affect the Metabolic Process of DSTs in Mussels by Altering the Expression of Genes Associated with Metabolism
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials and Animals
5.2. Experimental Design
5.3. Detection of DSTs
5.4. qPCR
5.5. Histological Examination
5.6. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shumway, S.E. A review of the effects of algal blooms on shellfish andaquaculture. J. World Aquacult. Soc. 1990, 21, 65–104. [Google Scholar] [CrossRef]
- Gerssen, A.; Pol-Hofstad, I.E.; Poelman, M.; Mulder, P.P.; Van den Top, H.J.; De Boer, J. Marine toxins: Chemistry, toxicity, occurrence and detection, with special reference to the Dutch situation. Toxins 2010, 2, 878–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Xu, X.; Wei, J.; Chen, J.; Miu, R.; Huang, L.; Zhou, X.; Fu, Y.; Yan, R.; Wang, Z.; et al. Food-borne disease outbreak of diarrhetic shellfish poisoning due to toxic mussel consumption: The first recorded outbreak in China. PLoS ONE 2013, 8, e65049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, M.; McIntyre, L.; Ritson, M.; Stone, J.; Bronson, R.; Bitzikos, O.; Rourke, W.; Galanis, E.; Outbreak Investigation, T. Outbreak of diarrhetic shellfish poisoning associated with mussels, British Columbia, Canada. Mar. Drugs 2013, 11, 1669–1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reguera, B.; Velo-Suárez, L.; Raine, R.; Park, M.G. Harmful Dinophysis species: A review. Harmful Algae 2012, 14, 87–106. [Google Scholar] [CrossRef]
- Nascimento, S.M.; Purdie, D.A.; Morris, S. Morphology, toxin composition and pigment content of Prorocentrum lima strains isolated from a coastal lagoon in southern UK. Toxicon 2005, 45, 633–649. [Google Scholar] [CrossRef]
- Kamat, P.K.; Rai, S.; Nath, C. Okadaic acid induced neurotoxicity: An emerging tool to study Alzheimer’s disease pathology. Neurotoxicology 2013, 37, 163–172. [Google Scholar] [CrossRef]
- Ma, J.; Li, J.H.; Hao, Y.M.; Nie, Y.Z.; Li, Z.S.; Qian, M.R.; Liang, Q.Y.; Yu, J.; Zeng, M.S.; Wu, K.C. Differentiated tumor immune microenvironment of Epstein–Barr virus-associated and negative gastric cancer: Implication in prognosis and immunotherapy. Oncotarget 2017, 8, 67094–67103. [Google Scholar] [CrossRef]
- Bernard, F.R. Uptake and elimination of coliform bacteria by four marine bivalve mollusks. Can. J. Fish. Aquat. Sci. 1989, 46, 1592–1599. [Google Scholar] [CrossRef]
- McCarron, P.; Kilcoyne, J.; Hess, P. Effects of cooking and heat treatment on concentration and tissue distribution of okadaic acid and dinophysistoxin-2 in mussels (Mytilus edulis). Toxicon 2008, 51, 1081–1089. [Google Scholar] [CrossRef]
- Reboreda, A.; Lago, J.; Chapela, M.J.; Vieites, J.M.; Botana, L.M.; Alfonso, A.; Cabado, A.G. Decrease of marine toxin content in bivalves by industrial processes. Toxicon 2010, 55, 235–243. [Google Scholar] [CrossRef]
- Blanco, J.; Arévalo, F.; Correa, J.; Porro, M.C.; Cabado, A.G.; Vieites, J.M.; Moroño, A. Effect of the industrial steaming on the toxicity, estimated by LC-MS/MS, of mussels exposed for a long time to diarrhetic shellfish poisoning (DSP) toxins. Food Chem. 2015, 177, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Fan, H.; Liu, T.; Liang, X.; Meng, F.; Quilliam, M.A.; Li, A. Application of activated carbon to accelerate detoxification of paralytic shellfish toxins from mussels Mytilus galloprovincialis and scallops Chlamys farreri. Ecotoxicol. Environ. Saf. 2018, 148, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.; Ruiz, Y.; Suárez, P.; Martinez, A.A.; Rossignoli, A.; Blanco, J.; Garcia, O.; San Juan, F. Accumulation of okadaic acid and detoxifying enzymes in the digestive gland of Mytilus galloprovincialis during exposure to DSP. In Molluscan Shellfish Safety; Springer: Berlin/Heidelberg, Germany, 2014; pp. 217–225. [Google Scholar]
- Chi, C.; Giri, S.S.; Jun, J.W.; Kim, S.W.; Kim, H.J.; Kang, J.W.; Park, S.C. Detoxification- and immune-related transcriptomic analysis of gills from bay scallops (Argopectenirradians) in response to algal toxin okadaic acid. Toxins 2018, 10, 308. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.M.; Lu, M.Y.; Duan, G.F.; Li, H.Y.; Liu, J.S.; Yang, W.D. Responses of CYP450 in the mussel Perna viridis after short-term exposure to the DSP toxins-producing dinoflagellate Prorocentrum lima. Ecotoxicol. Environ. Saf. 2019, 176, 178–185. [Google Scholar] [CrossRef]
- Gupta, A.K.; Lyons, D.C. The rise and fall of oral ketoconazole. J. Cutan. Med. Surg. 2015, 19, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Duan, G.F.; Liu, Y.; Zhang, L.N.; Li, H.Y.; Liu, J.S.; Yang, W.D. Cinnamaldehyde could reduce the accumulation of diarrhetic shellfish toxins in the digestive gland of the mussel Perna viridis under laboratory conditions. Mar. Drugs 2021, 19, 63. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.H.; Xia, J.; Sun, X.C.; Li, X.N.; Zhang, C.; Zhao, H.S.; Zhu, S.Y.; Li, J.L. A novel nuclear xenobiotic receptors (AhR/PXR/CAR)-mediated mechanism of DEHP-induced cerebellar toxicity in quails (Coturnix japonica) via disrupting CYP enzyme system homeostasis. Environ. Pollut. 2017, 226, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.V.; Boverhof, D.R.; Burgoon, L.D.; Fielden, M.R.; Zacharewski, T.R. Comparative analysis of dioxin response elements in human, mouse and rat genomic sequences. Nucleic Acids Res. 2004, 32, 4512–4523. [Google Scholar] [CrossRef] [Green Version]
- Esser, C.; Rannug, A. The aryl hydrocarbon receptor in barrier organ physiology, immunology, and toxicology. Pharmacol. Rev. 2015, 67, 259–279. [Google Scholar] [CrossRef] [Green Version]
- Islam, J.; Sato, S.; Watanabe, K.; Watanabe, T.; Ardiansyah; Hirahara, K.; Aoyama, Y.; Tomita, S.; Aso, H.; Komai, M.; et al. Dietary tryptophan alleviates dextran sodium sulfate-induced colitis through aryl hydrocarbon receptor in mice. J. Nutr. Biochem. 2017, 42, 43–50. [Google Scholar] [CrossRef]
- Zanette, J.; Jenny, M.J.; Goldstone, J.V.; Parente, T.; Woodin, B.R.; Bainy, A.C.; Stegeman, J.J. Identification and expression of multiple CYP1-like and CYP3-like genes in the bivalve mollusk Mytilus edulis. Aquat. Toxicol. 2013, 128–129, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Wang, K.; Wan, Y.J. Retinoids activate RXR/CAR-mediated pathway and induce CYP3A. Biochem. Pharmacol. 2010, 79, 270–276. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, S.; Brunet, F.G.; Escriva, H.; Parmentier, G.; Laudet, V.; Robinson-Rechavi, M. Evolutionary genomics of nuclear receptors: From twenty-five ancestral genes to derived endocrine systems. Mol. Biol. Evol. 2004, 21, 1923–1937. [Google Scholar] [CrossRef]
- Hernandez, J.P.; Mota, L.C.; Baldwin, W.S. Activation of CAR and PXR by dietary, environmental and occupational chemicals alters drug metabolism, intermediary metabolism, and cell proliferation. Curr. Pharmacogenom. Person Med. 2009, 7, 81–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruzeiro, C.; Lopes-Marques, M.; Ruivo, R.; Rodrigues-Oliveira, N.; Santos, M.M.; Rocha, M.J.; Rocha, E.; Castro, L.F. A mollusk VDR/PXR/CAR-like (NR1J) nuclear receptor provides insight into ancient detoxification mechanisms. Aquat. Toxicol. 2016, 174, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Oladimeji, P.O.; Chen, T.S. PXR: More than just a master xenobiotic receptor. Mol. Pharmacol. 2018, 93, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.A.; Gescher, A.J.; Steward, W.P. Curcumin: The story so far. Eur. J. Cancer 2005, 41, 1955–1968. [Google Scholar] [CrossRef]
- Zhou, H.; Beevers, C.S.; Huang, S. The targets of curcumin. Curr. Drug Targets 2011, 12, 332–347. [Google Scholar] [CrossRef]
- Appiah-Opong, R.; Commandeur, J.N.; van Vugt-Lussenburg, B.; Vermeulen, N.P. Inhibition of human recombinant cytochrome P450s by curcumin and curcumin decomposition products. Toxicology 2007, 235, 83–91. [Google Scholar] [CrossRef]
- Cho, Y.A.; Lee, W.; Choi, J.S. Effects of curcumin on the pharmacokinetics of tamoxifen and its active metabolite, 4-hydroxytamoxifen, in rats: Possible role of CYP3A4 and P-glycoprotein inhibition by curcumin. Pharmazie 2012, 67, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.C.; Liu, X.L.; Yang, X.H.; Zhang, C.H.; Bian, Z.Y. Accumulation and depuration of paralytic shellfish poisoning toxins in the oyster Ostrea rivularis Gould—Chitosan facilitates the toxin depuration. Food Control 2013, 30, 446–452. [Google Scholar] [CrossRef]
- Di, Y.; Aminot, Y.; Schroeder, D.C.; Readman, J.W.; Jha, A.N. Integrated biological responses and tissue-specific expression of p53 and ras genes in marine mussels following exposure to benzo(α)pyrene and C60 fullerenes, either alone or in combination. Mutagenesis 2017, 32, 77–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Geraldo Rde, J.; Garcia-Lagunas, N.; Hernandez-Saavedra, N.Y. Crassostrea gigas exposure to the dinoflagellate Prorocentrum lima: Histological and gene expression effects on the digestive gland. Mar. Environ. Res. 2016, 120, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Neves, R.A.F.; Santiago, T.C.; Carvalho, W.F.; Silva, E.D.S.; da Silva, P.M.; Nascimento, S.M. Impacts of the toxic benthic dinoflagellate Prorocentrum lima on the brown mussel Perna perna: Shell-valve closure response, immunology, and histopathology. Mar. Environ. Res. 2019, 146, 35–45. [Google Scholar] [CrossRef]
- Cuevas, N.; Zorita, I.; Costa, P.M.; Franco, J.; Larreta, J. Development of histopathological indices in the digestive gland and gonad of mussels: Integration with contamination levels and effects of confounding factors. Aquat. Toxicol. 2015, 162, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Zorita, I.; Ortiz-Zarragoitia, M.; Soto, M.; Cajaraville, M.P. Biomarkers in mussels from a copper site gradient (Visnes, Norway): An integrated biochemical, histochemical and histological study. Aquat. Toxicol. 2006, 78, S109–S116. [Google Scholar] [CrossRef] [PubMed]
- Carella, F.; Sardo, A.; Mangoni, O.; Di Cioccio, D.; Urciuolo, G.; De Vico, G.; Zingone, A. Quantitative histopathology of the Mediterranean mussel (Mytilus galloprovincialis L.) exposed to the harmful dinoflagellate Ostreopsis cf. ovata. J. Invertebr. Pathol. 2015, 127, 130–140. [Google Scholar] [CrossRef]
- Nelson, D.R.; Koymans, L.; Kamataki, T.; Stegeman, J.J.; Fey-ereisen, R.; Waxman, D.J.; Waterman, M.R.; Gotoh, O.; Coon, M.J.; Estabrook, R.W.; et al. P450 superfamily: Update on new sequences, genemapping, accession numbers and nomenclature. Pharmacogenet. Genom. 1996, 6, 1–42. [Google Scholar] [CrossRef] [PubMed]
- Ferron, P.J.; Hogeveen, K.; De Sousa, G.; Rahmani, R.; Dubreil, E.; Fessard, V.; Le Hegarat, L. Modulation of CYP3A4 activity alters the cytotoxicity of lipophilic phycotoxins in human hepatic HepaRG cells. Toxicol. In Vitro 2016, 33, 136–146. [Google Scholar] [CrossRef]
- Cubero-Leon, E.; Puinean, A.M.; Labadie, P.; Ciocan, C.; Itoh, N.; Kishida, M.; Osada, M.; Minier, C.; Hill, E.M.; Rotchell, J.M. Two CYP3A-like genes in the marine mussel Mytilus edulis: mRNA expression modulation following short-term exposure to endocrine disruptors. Mar. Environ. Res. 2012, 74, 32–39. [Google Scholar] [CrossRef]
- Kumagai, T.; Suzuki, H.; Sasaki, T.; Sakaguchi, S.; Miyairi, S.; Yamazoe, Y.; Nagata, K. Polycyclic aromatic hydrocarbons activate CYP3A4 gene transcription through human pregnane X receptor. Drug Metab. Pharmacokinet. 2012, 27, 200–206. [Google Scholar] [CrossRef] [Green Version]
- Ciolino, H.P.; Daschner, P.J.; Wang, T.Y.; Yeh, G.C. Effect of curcumin on the aryl hydrocarbon receptor and cytochrome P450 1A1 in MCF-7 human breast carcinoma cells. Biochem. Pharmacol. 1998, 56, 197–206. [Google Scholar] [CrossRef]
- Rinaldi, A.L.; Morse, M.A.; Fields, H.W.; Rothas, D.A.; Pei, P.; Rodrigo, K.A.; Renner, R.J.; Mallery, S.R. Curcumin activates the aryl hydrocarbon receptor yet significantly inhibits (-)-benzo(a)pyrene-7R-trans-7,8-dihydrodiol bioactivation in oral squamous cell carcinoma cells and oral mucosa. Cancer Res. 2002, 62, 5451–5456. [Google Scholar]
- Busbee, P.B.; Rouse, M.; Nagarkatti, M.; Nagarkatti, P.S. Use of natural AhR ligands as potential therapeutic modalities against inflammatory disorders. Nutr. Rev. 2013, 71, 353–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schug, T.T.; Berry, D.C.; Shaw, N.S.; Travis, S.N.; Noy, N. Opposing effects of retinoic acid on cell growth result from alternate activation of two different nuclear receptors. Cell 2007, 129, 723–733. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Chen, J.; Bao, Y.; Li, J. Genomic analysis of NF-κB signaling pathway reveals its complexity in Crassostrea gigas. Fish. Shellfish Immunol. 2018, 72, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, D.; Zhang, Z.; Pan, L. In vitro recombinant yeast assay reveals the binding of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) and aryl hydrocarbon receptor (AhR) from scallop Chlamys farreri. Toxicol. In Vitro 2019, 59, 64–69. [Google Scholar] [CrossRef]
- Fonseca, E.S.S.; Ruivo, R.; Machado, A.M.; Conrado, F.; Tay, B.H.; Venkatesh, B.; Santos, M.M.; Castro, L.F.C. Evolutionary plasticity in detoxification gene modules: The preservation and loss of the pregnane X receptor in chondrichthyes lineages. Int. J. Mol. Sci. 2019, 20, 2331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Chen, J.; Li, Z.; Wang, Y.; Fu, B.; Han, X.; Zheng, L. Cultivation of the benthic microalga Prorocentrum lima for the production of diarrhetic shellfish poisoning toxins in a vertical flat photobioreactor. Bioresour. Technol. 2015, 179, 243–248. [Google Scholar] [CrossRef]
- Chen, J.; Wang, Y.; Pan, L.; Shen, H.; Fu, D.; Fu, B.; Sun, C.; Zheng, L. Separation and purification of two minor typical diarrhetic shellfish poisoning toxins from harmful marine microalgae via combined liquid chromatography with mass spectrometric detection. J. Sep. Sci. 2017, 40, 2906–2913. [Google Scholar] [CrossRef] [PubMed]
- Danielli, N.M.; Trevisan, R.; Mello, D.F.; Fischer, K.; Deconto, V.S.; da Silva Acosta, D.; Bianchini, A.; Bainy, A.C.; Dafre, A.L. Upregulating Nrf2-dependent antioxidant defenses in Pacific oysters Crassostrea gigas: Investigating the Nrf2/Keap1 pathway in bivalves. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2017, 195, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Bauder, A.G.; Cembella, A.D.; Bricelj, V.M.; Quilliam, M.A. Uptake and fate of diarrhetic shellfish poisoning toxins from the dinoflagellate Prorocentrum lima in the bay scallop Argopecten irradians. Mar. Ecol. Prog. Ser. 2001, 213, 39–52. [Google Scholar] [CrossRef] [Green Version]
- Fang, L.; Yao, X.; Wang, L.; Li, J. Solid-phase extraction-based ultra-sensitive detection of four lipophilic marine biotoxins in bivalves by high-performance liquid chromatography-tandem mass spectrometry. J. Chromatogr. Sci. 2015, 53, 373–379. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.0031. [Google Scholar] [CrossRef] [Green Version]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Derveaux, S.; Vandesompele, J.; Hellemans, J. How to do successful gene expression analysis using real-time PCR. Methods 2010, 50, 227–230. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (5′–3′) | Amplicon Size (bp) | |
|---|---|---|---|
| EF1-Fα | F: | CACTCCGTCTTCCACTCCA | 131 |
| R: | CCTCTGGCATTGACTCGTG | ||
| Tubulin-β | F: | AGGAAGGAGGCTGAGAGTTGT | 135 |
| R: | TTTGGAGATGAGCAGGGTTC | ||
| RPL3 | F: | GGTGGCACTATCTCCCAGAA | 98 |
| R: | GCCATCTGGACGTTACACCT | ||
| UBA52 | F: | TTACATTTGGTCCTGCGTCTC | 135 |
| R: | CAGTTGGTAGCCCTTTGATGA | ||
| RPL13 | F: | TAAAGACTGGCAACGCTATGT | 155 |
| R: | TCACAACTGGTCGGAGAAG | ||
| RPL37 | F: | GTCGCAATAAGACGCACACGTTG | 179 |
| R: | GTGCCTCATTCGACCAGTTCCG | ||
| AhR | F: | GAGAGTTCTGGAGTGTTCGTGGAC | 109 |
| R: | TGAGGACACGGTTCAGTCAG | ||
| HR96 | F: | TGGAGCCTGATGAAGAATACC | 194 |
| R: | ACTGAGAGAGAGCCTTGAATG | ||
| CYP3A4 | F: | ACCAATGCAAATGCGTGGTC | 99 |
| R: | TCAGCAGGCAGTATTCCAGGAGAG | ||
| CYP3A1 | F: | CGCTGCTGTGACGATCTGGTAG | 141 |
| R: | TCTCTGCGAATTCACCTGCAACC | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, K.-K.; Duan, G.-F.; Liu, Q.-Y.; Li, H.-Y.; Yang, W.-D. Inhibition of Diarrheal Shellfish Toxins Accumulation in the Mussel Perna viridis by Curcumin and Underlying Mechanisms. Toxins 2021, 13, 578. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080578
Yuan K-K, Duan G-F, Liu Q-Y, Li H-Y, Yang W-D. Inhibition of Diarrheal Shellfish Toxins Accumulation in the Mussel Perna viridis by Curcumin and Underlying Mechanisms. Toxins. 2021; 13(8):578. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080578
Chicago/Turabian StyleYuan, Kuan-Kuan, Guo-Fang Duan, Qing-Yuan Liu, Hong-Ye Li, and Wei-Dong Yang. 2021. "Inhibition of Diarrheal Shellfish Toxins Accumulation in the Mussel Perna viridis by Curcumin and Underlying Mechanisms" Toxins 13, no. 8: 578. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13080578