N-Acetylcysteine Inhibits Patulin-Induced Apoptosis by Affecting ROS-Mediated Oxidative Damage Pathway

Abstract
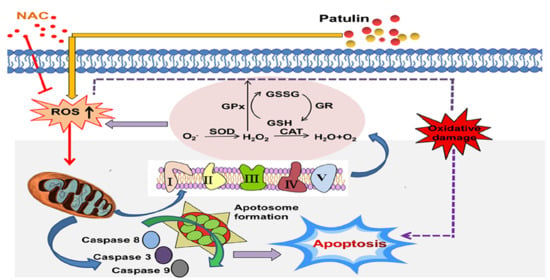
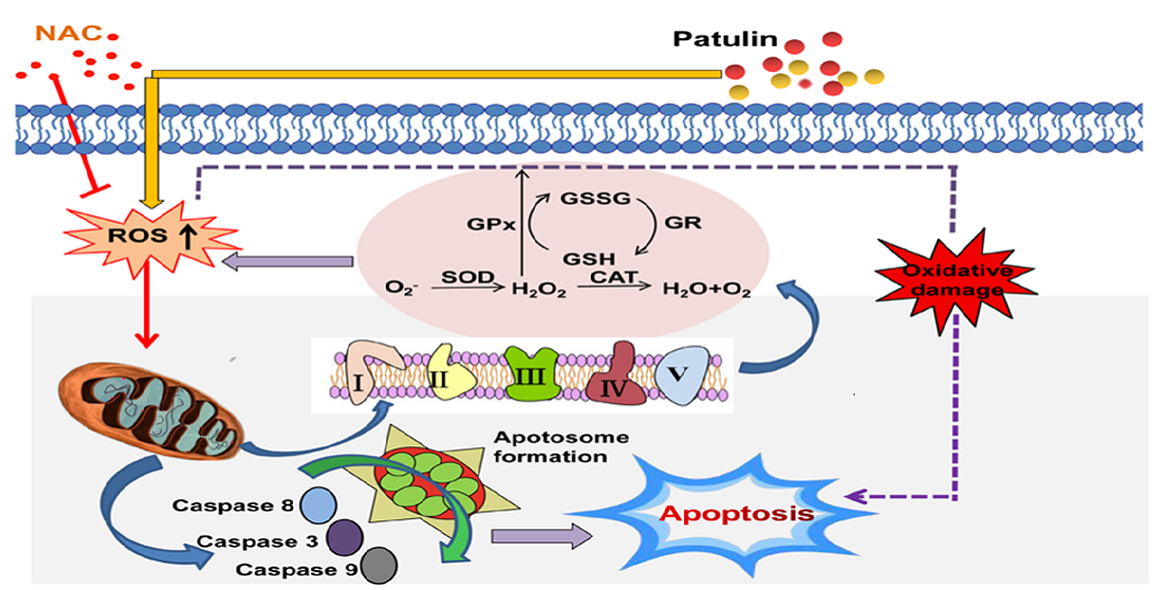
:
1. Introduction
2. Result
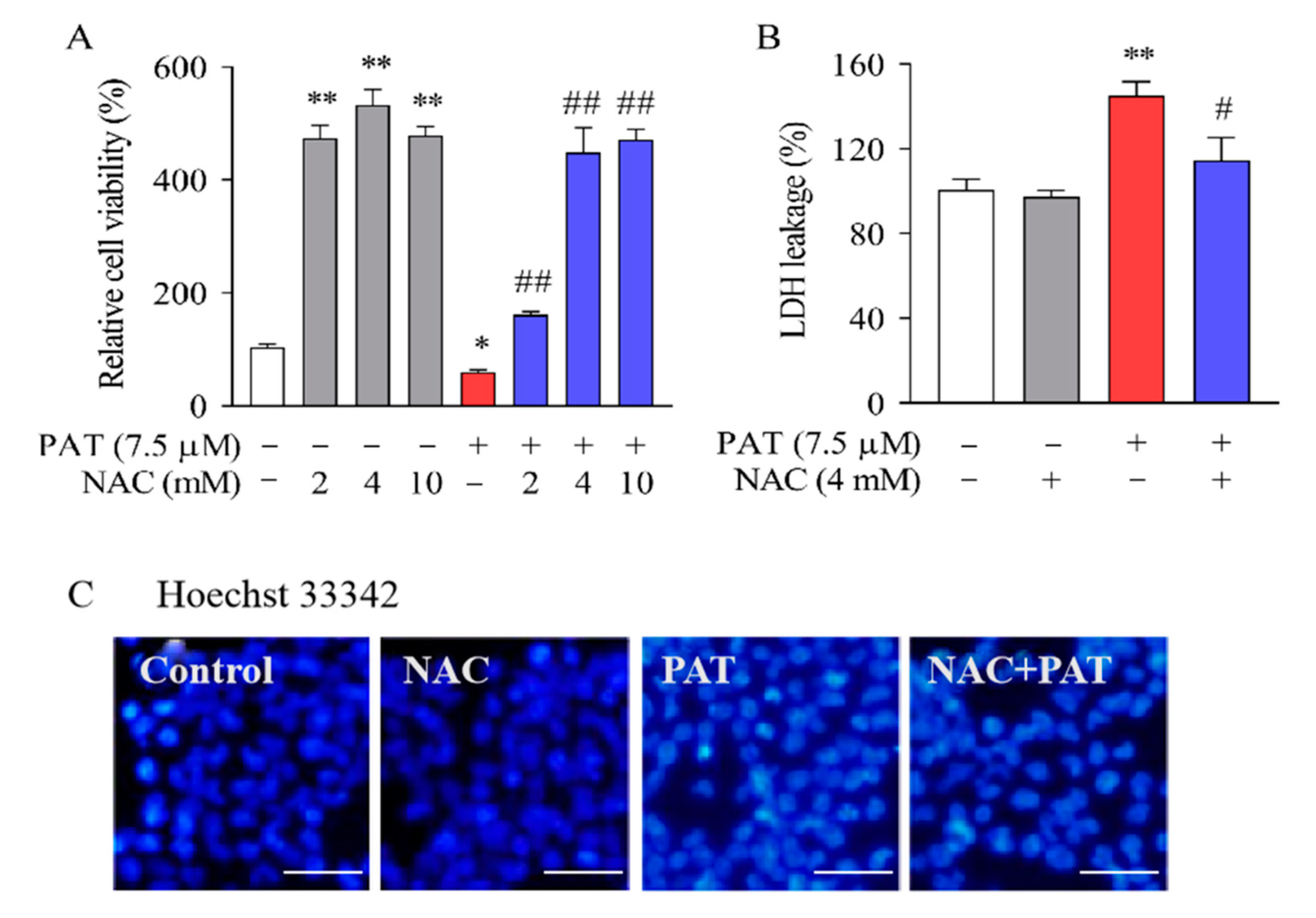
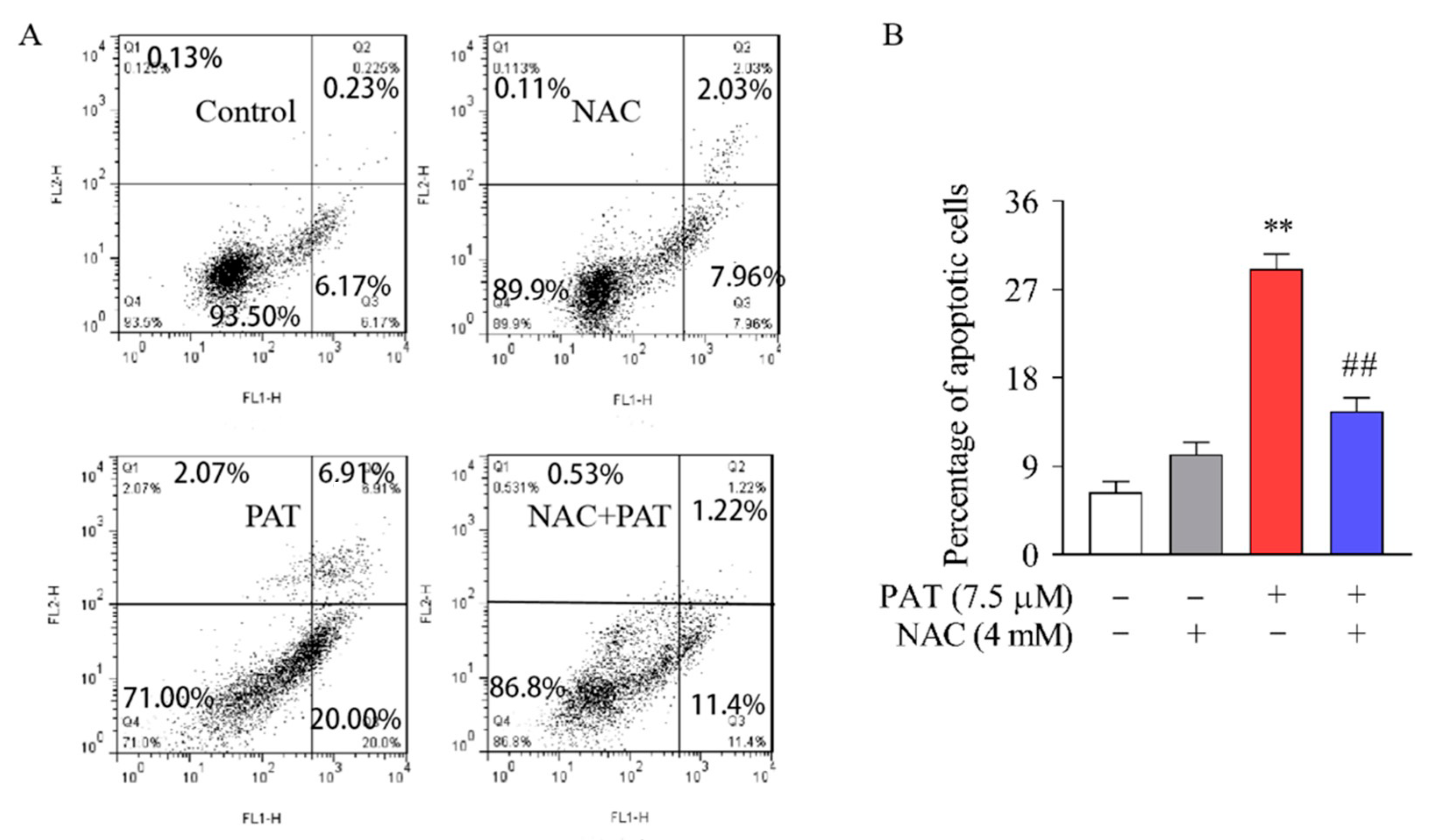
2.1. PAT Induced Cytotoxicity and Apoptosis in HEK293 Cells, Diminished by NAC
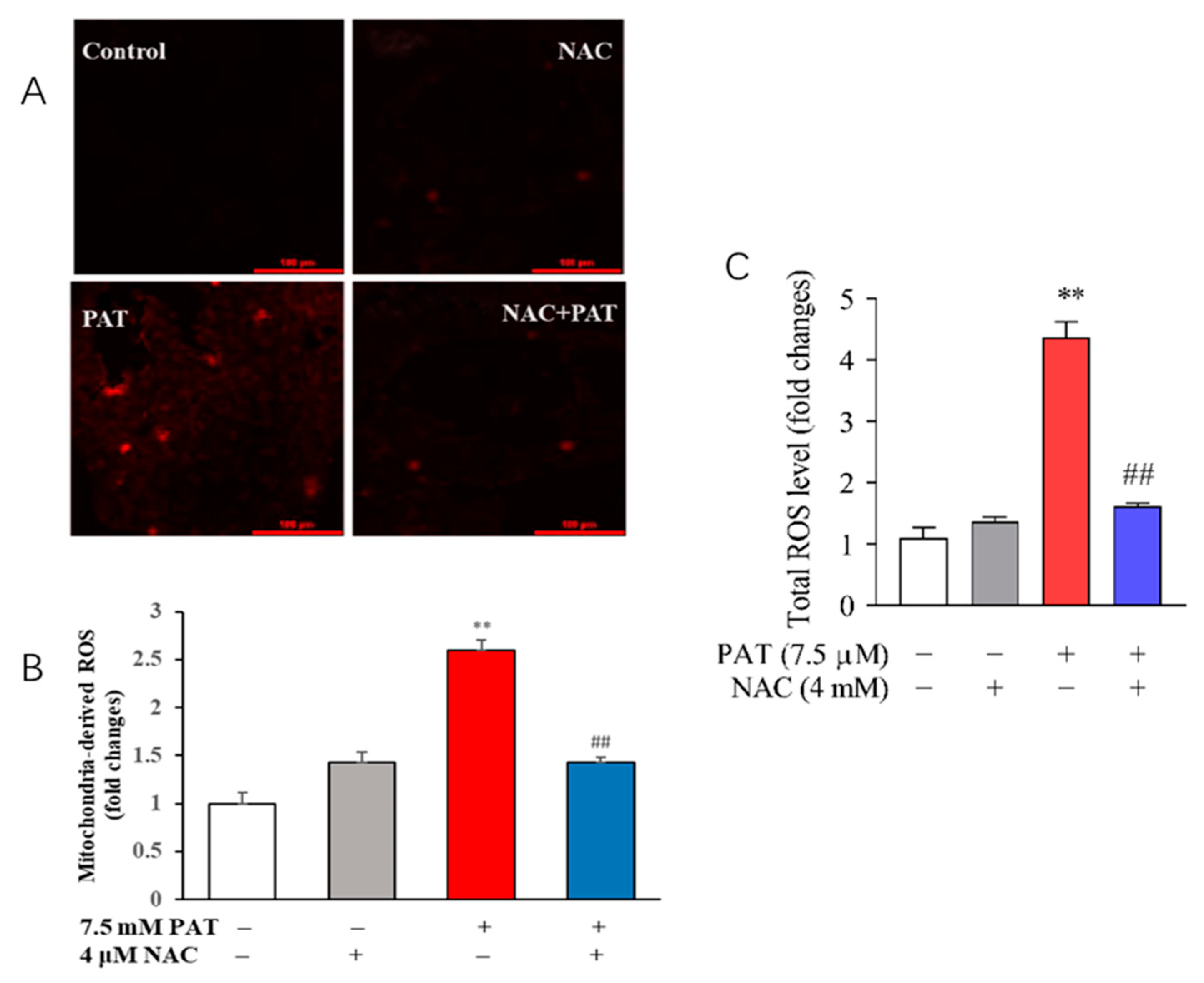
2.2. NAC Reduced ROS Generation in Response to PAT Exposure
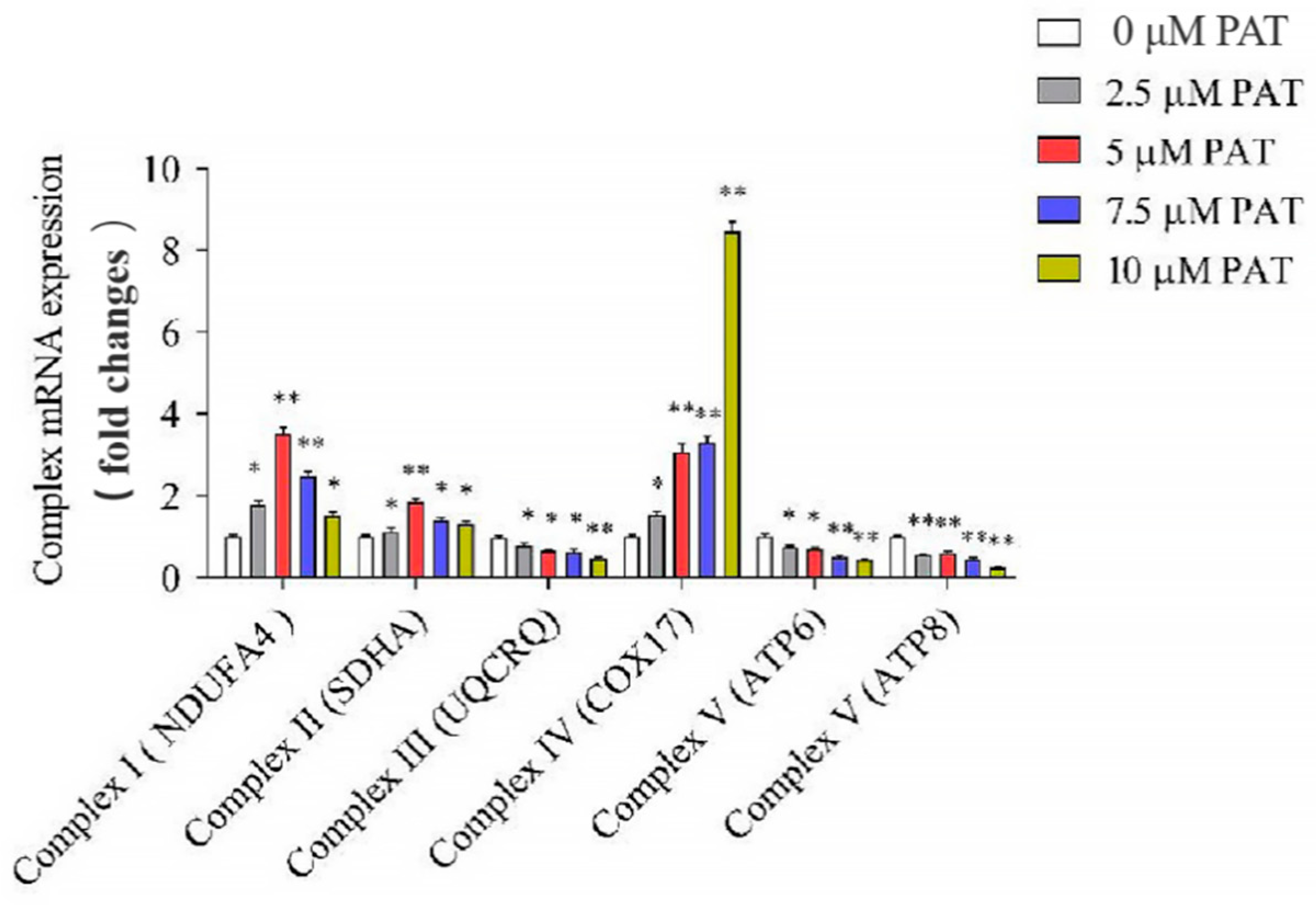
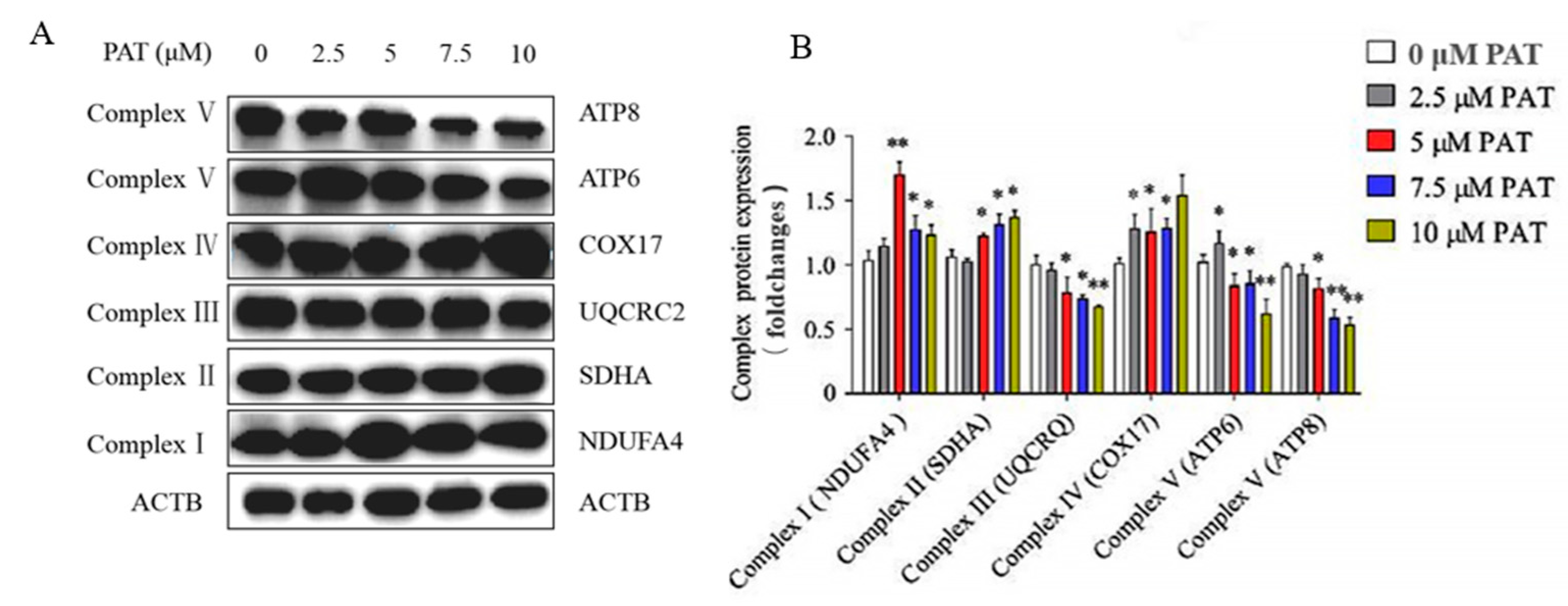
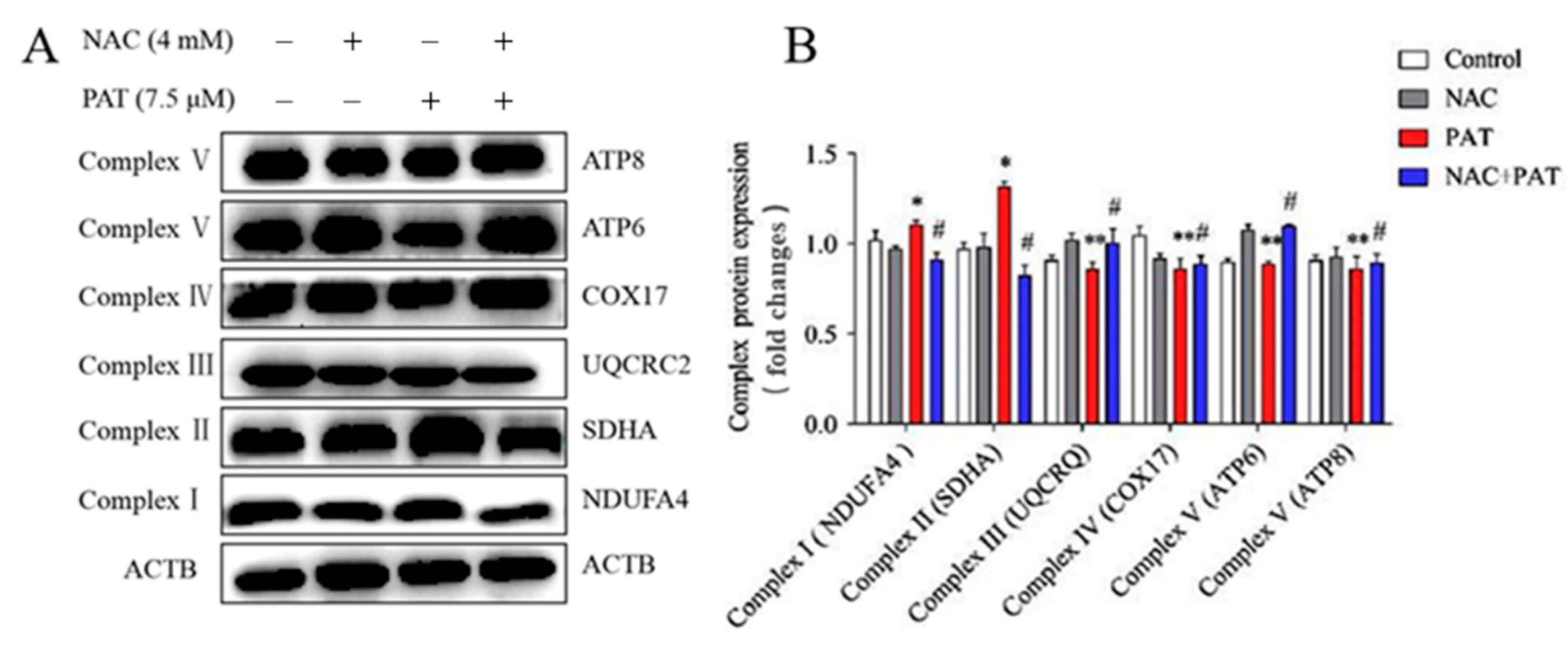
2.3. PAT Caused the Impairment of the Mitochondrial Respiratory Chain Complex Signaling Pathway
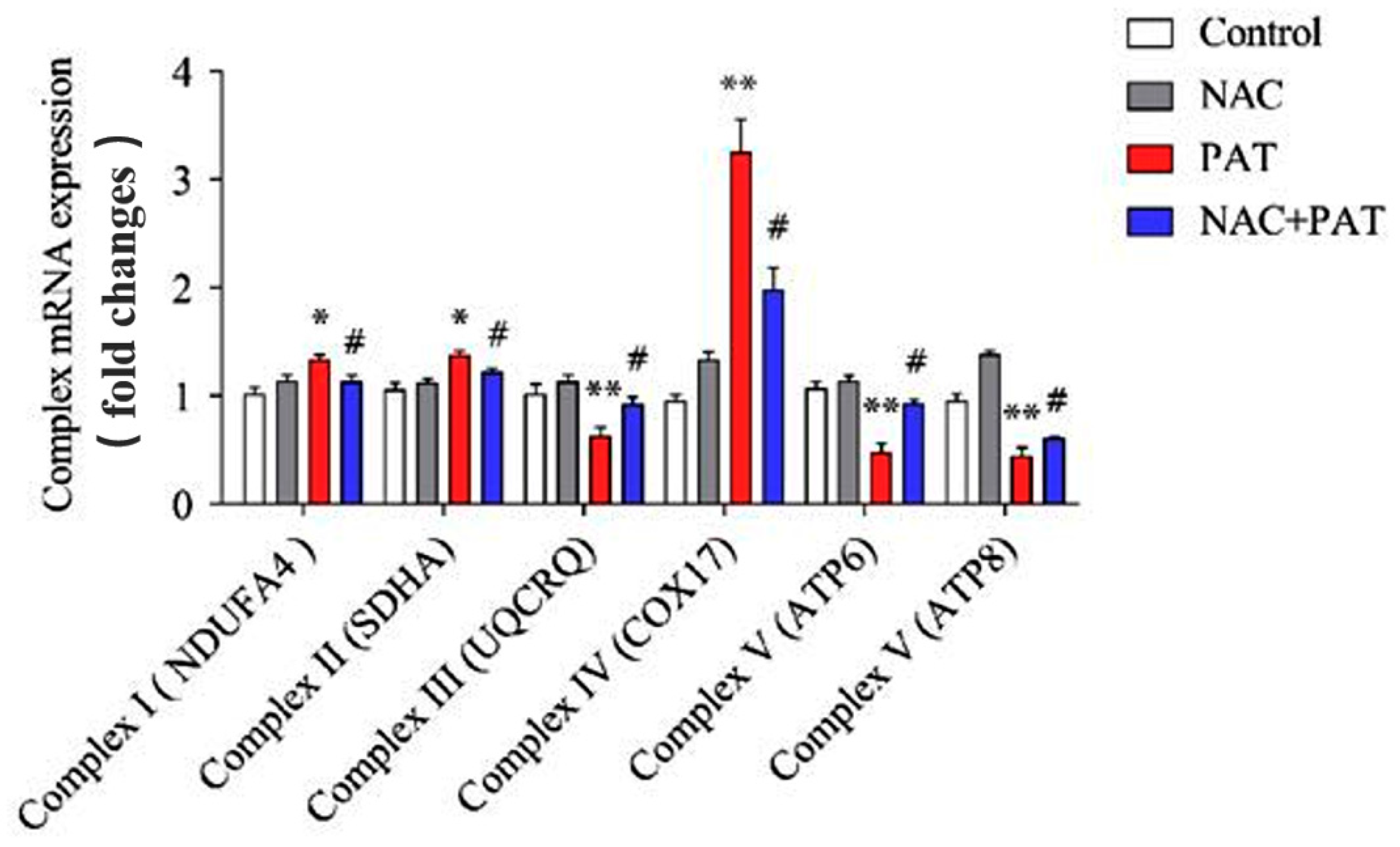
2.4. NAC Improved the Disorders of the Mitochondrial Respiratory Chain Complex Induced by PAT
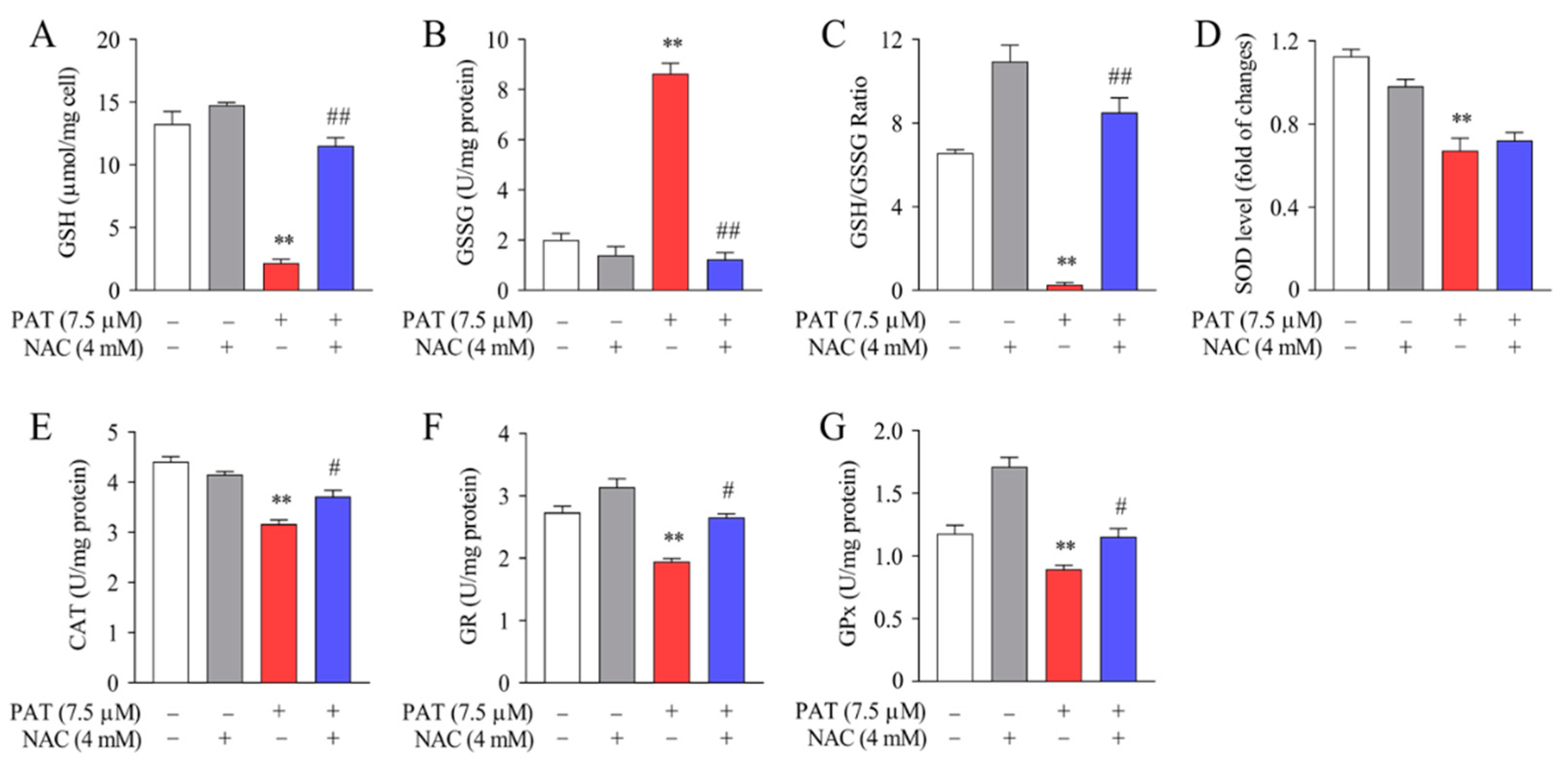
2.5. NAC Protected against PAT-Induced GSH Depletion and Improved Antioxidant Enzyme Activities
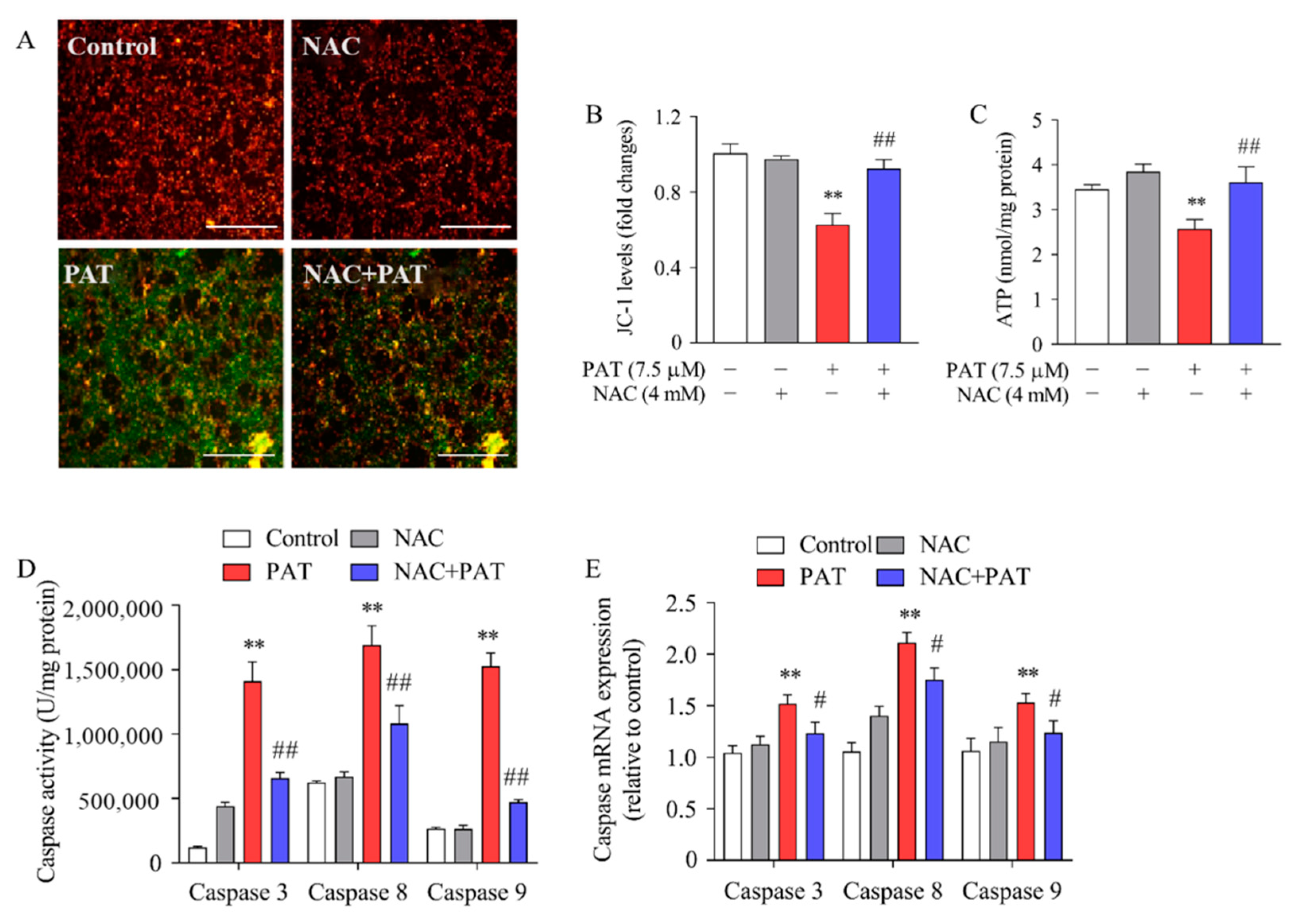
2.6. NAC Inhibited PAT-Induced Mitochondrial Dysfunction and Caspase-Dependent Apoptotic Pathway
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Cell Cultivation and Treatment
5.3. Cell Viability Assays
5.4. Hoechst 33342 Staining
5.5. Apoptosis Assessment
5.6. Measurement of Mitochondrial Membrane Potential
5.7. Measurement of ATP Level
5.8. Measurement of Total ROS and Mitochondrial ROS
5.9. Analysis of GSH and GSSG Contents
5.10. Activities of SOD, CAT, GR, and GPx
5.11. Caspase Activity Assay
5.12. Quantitative Real-Time Reverse Transcription PCR (qRT-PCR)
5.13. Western Blot Analysis
5.14. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer | Reverse Primer | Amplicon Size (bp) |
|---|---|---|---|
| ACTB | TCGTGCGTGACATTAAGGAG | AGGAAGGAAGGCTGGAAGAG | 178 |
| NDUFA4 | AGCTTGATCCCCCTCTTTGT | TCATTGGGACCCAGTTTGTT | 140 |
| SDHA | CCTACCAGGTCACACACTGT | AGTCCGATCAGCCACACAG | 162 |
| UQCRC2 | TCAAGAACTGGTCCCGAC | TGTAAGGTACCCAGTCCAGG | 102 |
| COX17 | TAGATTTGGCTGTCTCCGCT | CTCTCATGCATTCCTTGTGGG | 213 |
| ATP6 | GGACTCCTGCCTCACTCATT | ATAAGGGGTGTAGGTGTGCC | 158 |
| ATP8 | ATGGCCCACCATAATTACCC | GCAATGAATGAAGCGAACAG | 170 |
| Caspase 3 | TCCTGAGATGGGTTTATGT | ATGTTTCCCTGAGGTTTGC | 110 |
| Caspase 8 | CTCCAAATGCAAACTGGATG | TGTTGATTTGGGCACAGACT | 122 |
| Caspase 9 | CCGCATACTCCATCCTCAGT | GCTTCTTCATCCCAAAGTCG | 123 |
References
- Braun, H.P. The oxidative phosphorylation system of the mitochondria in plants. Mitochondrion 2020, 53, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.H.; Wu, T.S.; Yu, F.Y.; Su, C.C. Induction of oxidative stress response by the mycotoxin patulin in mammalian cells. Toxicol. Sci. 2007, 95, 340–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Zhang, E.; Yin, S.; Fan, L.; Hu, H. Methylseleninic acid prevents patulin-induced hepatotoxicity and nephrotoxicity via the inhibition of oxidative stress and inactivation of p53 and MAPKs. J. Agric. Food Chem. 2017, 65, 5299–5305. [Google Scholar] [CrossRef]
- Boussabbeh, M.; Ben Salem, I.; Prola, A.; Guilbert, A.; Bacha, H.; Abid-Essefi, S.; Lemaire, C. Patulin induces apoptosis through ROS-mediated endoplasmic reticulum stress pathway. Toxicol. Sci. 2015, 144, 328–337. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Yin, S.; Song, X.; Zhang, E.; Fan, L.; Hu, H. p53 activation contributes to patulin-induced nephrotoxicity via modulation of reactive oxygen species generation. Sci. Rep. 2016, 6, 24455. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.H.; Wu, T.S.; Yu, F.Y.; Wang, C.H. Mycotoxin patulin activates the p38 kinase and JNK signaling pathways in human embryonic kidney cells. Toxicol. Sci. 2006, 89, 423–430. [Google Scholar] [CrossRef]
- Guerra-Moreno, A.; Hanna, J. Induction of proteotoxic stress by the mycotoxin patulin. Toxicol. Lett. 2017, 276, 85–91. [Google Scholar] [CrossRef]
- Pillay, Y.; Phulukdaree, A.; Nagiah, S.; Chuturgoon, A.A. Patulin triggers NRF2-mediated survival mechanisms in kidney cells. Toxicon 2015, 99, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Puel, O.; Galtier, P.; Oswald, I.P. Biosynthesis and toxicological effects of patulin. Toxins 2010, 2, 613–631. [Google Scholar] [CrossRef] [Green Version]
- Kunio, D.; Koji, U. Mechanisms of mycotoxin-induced dermal toxicity and tumorigenesis through oxidative stress-related pathways. J. Toxicol. Pathol. 2014, 27, 1–10. [Google Scholar]
- Boussabbeh, M.; Prola, A.; Ben Salem, I.; Guilbert, A.; Bacha, H.; Lemaire, C.; Abis-Essefi, S. Crocin and quercetin prevent PAT-induced apoptosis in mammalian cells: Involvement of ROS-mediated ER stress pathway. Environ. Toxicol. 2016, 31, 1851–1858. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Peng, X.; Li, G.; Xu, Y.; Xia, X.; Wang, Q. Oxidative stress is involved in Patulin induced apoptosis in HEK293 cells. Toxicon 2015, 94, 1–7. [Google Scholar] [CrossRef]
- Grancara, S.; Zonta, F.; Ohkubo, S.; Brunati, A.M.; Agostinelli, E.; Toninello, A. Pathophysiological implications of mitochondrial oxidative stress mediated by mitochondriotropic agents and polyamines: The role of tyrosine phosphorylation. Amino Acids 2015, 47, 869–883. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, M.; Filosto, M.; Orsucci, D.; Siciliano, G. Mitochondrial DNA sequence variation and neurodegeneration. Hum. Genom. 2008, 3, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.; Cao, M.; Liu, J.; Yang, Q.; Miao, X.; Go, V.L.W.; Lee, P.W.N.; Xiao, G.G. Metabolic regulation in mitochondria and drug resistance. Mitochondrial DNA Dis. 2017, 1038, 149–171. [Google Scholar]
- Castellanos, E.; Lanning, N.J. Phosphorylation of OXPHOS machinery subunits: Functional implications in cell biology and disease. Yale J. Biol. Med. 2019, 92, 523–531. [Google Scholar] [PubMed]
- Xu, Q.; Vu, H.; Liu, L.; Wang, T.C.; Schaefer, W.H. Metabolic profiles show specific mitochondrial toxicities in vitro in myotube cells. J. Biomol. NMR 2011, 49, 207–219. [Google Scholar] [CrossRef]
- Yang, Y.; Karakhanova, S.; Hartwig, W.; D’Haese, J.G.; Philippov, P.P.; Werner, J.; Bazhin, A.V. Mitochondria and mitochondrial ROS in cancer: Novel targets for anticancer therapy. J. Cell. Physiol. 2016, 231, 2570–2581. [Google Scholar] [CrossRef]
- Cecarini, V.; Gee, J.; Fioretti, E.; Amici, M.; Angeletti, M.; Eleuteri, A.M.; Keller, J.N. Protein oxidation and cellular homeostasis: Emphasis on metabolism. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2007, 1773, 93–104. [Google Scholar] [CrossRef]
- Kietzmann, T. Cellular redox compartments. Antioxid. Redox Signal. 2019, 30, 1–4. [Google Scholar] [CrossRef]
- Qi, J.; Wang, J.; Gong, Z.; Zhou, J.M. Apoplastic ROS signaling in plant immunity. Curr. Opin. Plant Biol. 2017, 38, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Kala, M.; Shaikh, M.V.; Nivsarkar, M. Equilibrium between anti-oxidants and reactive oxygen species: A requisite for oocyte development and maturation. Reprod. Med. Biol. 2017, 16, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Gibb, Z.; Baker, M.A.; Drevet, J.; Gharagozloo, P. Causes and consequences of oxidative stress in spermatozoa. Reprod. Fertil. Dev. 2016, 28, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xiong, S.; Zhao, F.; Lu, X.; Wu, B.; Yang, B. Effect of magnesium on reducing the UV-induced oxidative damage in marrow mesenchymal stem cells. J. Biomed. Mater. Res. Part A 2019, 107, 1253–1263. [Google Scholar] [CrossRef] [PubMed]
- Sabeti, P.; Pourmasumi, S.; Rahiminia, T.; Akyash, F.; Talebi, A.R. Etiologies of sperm oxidative stress. Int. J. Reprod. BioMed. 2016, 14, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Kajarabille, N.; Latunde-Dada, G.O. Programmed cell-death by ferroptosis: Antioxidants as mitigators. Int. J. Mol. Sci. 2019, 20, 4968. [Google Scholar] [CrossRef] [Green Version]
- Cui, B.; Zhang, X.; Han, G.; Li, K. Antioxidant defense response and growth reaction of amorpha fruticosa seedlings in petroleum-contaminated soil. Water Air Soil Pollut. 2016, 227, 121. [Google Scholar] [CrossRef]
- Ferrareze, J.P.; Fugate, K.K.; Bolton, M.D.; Deckard, E.L.; Campbell, L.G.; Finger, F.L. Jasmonic acid does not increase oxidative defense mechanisms or common defense-related enzymes in postharvest sugarbeet roots. Postharvest Biol. Technol. 2013, 77, 11–18. [Google Scholar] [CrossRef]
- Krishnan, N.; Kodrik, D.; Turanli, F.; Sehnal, F. Stage-specific distribution of oxidative radicals and antioxidant enzymes in the midgut of Leptinotarsa decemlineata. J. Insect Physiol. 2007, 53, 67–74. [Google Scholar] [CrossRef]
- Akhtar, M.J.; Ahamed, M.; Alhadlaq, H.A.; Alshamsan, A. Mechanism of ROS scavenging and antioxidant signalling by redox metallic and fullerene nanomaterials: Potential implications in ROS associated degenerative disorders. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2017, 1861, 802–813. [Google Scholar] [CrossRef]
- Giancarlo, A.; Alessandra, A.; Giovanna, B.; Giulio, V.; Marina, C.; Luisa, B.; Francesco, S. N-Acetylcysteine as an antioxidant and disulphide breaking agent: The reasons why. Free Radic. Res. 2018, 52, 751–762. [Google Scholar]
- Xie, C.; Yi, J.; Lu, J.; Nie, M.; Huang, M.; Rong, J.; Zhu, Z.; Chen, J.; Zhou, X.; Li, B.; et al. N-Acetylcysteine reduces ROS-mediated oxidative DNA damage and PI3K/Akt pathway activation induced by helicobacter pylori Infection. Oxidative Med. Cell. Longev. 2018, 2018, 1874985. [Google Scholar] [CrossRef] [Green Version]
- Niraula, P.; Kim, M.S. N-Acetylcysteine extends lifespan of Drosophila via modulating ROS scavenger gene expression. Biogerontology 2019, 20, 533–543. [Google Scholar] [CrossRef]
- Jin, L.; Ni, J.; Tao, Y.; Weng, X.; Zhu, Y.; Yan, J.; Hu, B. N-acetylcysteine attenuates PM 2.5-induced apoptosis by ROS-mediated Nrf2 pathway in human embryonic stem cells. Sci. Total Environ. 2019, 666, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Lisa, C.C.; Brooke, J.M.; Adrian, P.S.; Nigel, J.W. Quantitation of apoptosis and necrosis by Annexin V binding, Propidium Iodide uptake, and flow cytometry. Cold Spring Harb. Protoc. 2016, 11, pdb-prot087288. [Google Scholar]
- Droese, S.; Brandt, U. Molecular mechanisms of superoxide production by the mitochondrial respiratory chain. Mitochondrial Oxidative Phosphorylation 2012, 748, 145–169. [Google Scholar]
- Carrillo-Vico, A.; Murillo-Cabezas, F.; Egea-Guerrero, J.J.; Rodriguez-Rodriguez, A. Oxidative stress in traumatic brain injury. Curr. Med. Chem. 2014, 21, 1201–1211. [Google Scholar]
- Kopeina, G.S.; Prokhorova, E.A.; Lavrik, I.N.; Zhivotovsky, B. Alterations in the nucleocytoplasmic transport in apoptosis: Caspases lead the way. Cell Prolif. 2018, 51, e12467–e12469. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Jin, C.; Zhong, Y.; Li, X.; Han, J.; Xue, W.; Wu, P.; Xia, X.; Peng, X. Glutathione reduction of patulin-evoked cytotoxicity in HEK293 cells by the prevention of oxidative damage and the mitochondrial apoptotic pathway. J. Agric. Food Chem. 2018, 66, 7775–7785. [Google Scholar] [CrossRef]
- Ramalingam, S.; Bahuguna, A.; Kim, M. The effects of mycotoxin patulin on cells and cellular components. Trends Food Sci. Technol. 2019, 83, 99–113. [Google Scholar] [CrossRef]
- Mai, N.; Chrzanowska-Lightowlers, Z.M.A.; Lightowlers, R.N. The process of mammalian mitochondrial protein synthesis. Cell Tissue Res. 2017, 367, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Signes, A.; Fernandez-Vizarra, E. Assembly of mammalian oxidative phosphorylation complexes I-V and supercomplexes. Essays Biochem. 2018, 62, 255–270. [Google Scholar] [PubMed]
- DiMauro, S.; Schon, E.A. Mechanisms of disease: Mitochondrial respiratory-chain diseases. N. Engl. J. Med. 2003, 348, 2656–2668. [Google Scholar] [CrossRef]
- Du, S.N.N.; Khajali, F.; Dawson, N.J.; Scott, G.R. Hybridization increases mitochondrial production of reactive oxygen species in sunfish. Evolution 2017, 71, 1643–1652. [Google Scholar] [CrossRef]
- Wojtala, A.; Karkucinska-Wieckowska, A.; Sardao, V.A.; Szczepanowska, J.; Kowalski, P.; Pronicki, M.; Duszynski, J.; Wieckowski, M.R. Modulation of mitochondrial dysfunction-related oxidative stress in fibroblasts of patients with Leigh syndrome by inhibition of prooxidative p66Shc pathway. Mitochondrion 2017, 37, 62–79. [Google Scholar] [CrossRef]
- Mokhtari, V.; Afsharian, P.; Shahhoseini, M.; Kalantar, S.M.; Moini, A. A review on various uses of N-acetyl cysteine. Cell J. 2017, 19, 11–17. [Google Scholar] [PubMed]
- Yamada, M.; Tsukimura, N.; Ikeda, T.; Sugita, Y.; Att, W.; Kojima, N.; Kubo, K.; Ueno, T.; Sakurai, K.; Ogawa, T. N-acetyl cysteine as an osteogenesis-enhancing molecule for bone regeneration. Biomaterials 2013, 34, 6147–6156. [Google Scholar] [CrossRef] [PubMed]
- Samarghandian, S.; Azimi-Nezhad, M.; Farkhondeh, T.; Samini, F. Anti-oxidative effects of curcumin on immobilization-induced oxidative stress in rat brain, liver and kidney. Biomed. Pharmacother. 2017, 87, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.A.; Roberts, K.N.; Scheff, S.W. Oxidative stress and modification of synaptic proteins in hippocampus after traumatic brain injury. Free. Radic. Biol. Med. 2008, 45, 443–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limon-Pacheco, J.; Gonsebatt, M.E. The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress. Mutat. Res./Genet. Toxicol. Environ. Mutagenesis 2009, 674, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem.-Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Girindrababu, V.J.; Krishnaswamy, K.; Puttasiddiah, R.; Farhath, K. Patulin induced oxidative stress mediated apoptotic damage in mice, and its modulation by green tea leaves. J. Clin. Exp. Hepatol. 2017, 7, 127–134. [Google Scholar]
- He, M.D.; Xu, S.C.; Lu, Y.H.; Li, L.; Zhong, M.; Zhang, Y.W.; Wang, Y.; Li, M.; Yang, J.; Zhang, G.B.; et al. L-carnitine protects against nickel-induced neurotoxicity by mitochondrial function in Neuro-2a cells. Toxicol. Appl. Pharmacol. 2011, 253, 38–44. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Liu, Q.; Han, J.; Feng, J.; Guo, T.; Li, Z.; Min, F.; Jin, R.; Peng, X. N-Acetylcysteine Inhibits Patulin-Induced Apoptosis by Affecting ROS-Mediated Oxidative Damage Pathway. Toxins 2021, 13, 595. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090595
Liu J, Liu Q, Han J, Feng J, Guo T, Li Z, Min F, Jin R, Peng X. N-Acetylcysteine Inhibits Patulin-Induced Apoptosis by Affecting ROS-Mediated Oxidative Damage Pathway. Toxins. 2021; 13(9):595. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090595
Chicago/Turabian StyleLiu, Jiayu, Qi Liu, Jiahui Han, Jiayu Feng, Tianmin Guo, Zhiman Li, Fenyi Min, Ruyi Jin, and Xiaoli Peng. 2021. "N-Acetylcysteine Inhibits Patulin-Induced Apoptosis by Affecting ROS-Mediated Oxidative Damage Pathway" Toxins 13, no. 9: 595. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090595