Solanum nigrum Extract and Solasonine Affected Hemolymph Metabolites and Ultrastructure of the Fat Body and the Midgut in Galleria mellonella

 , , ,
, , ,  , , ,
, , ,  , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. The Effects of S. nigrum Extract and Pure Glycoalkaloids on Sugars, Polyols, and Proline Concentrations in the Hemolymph
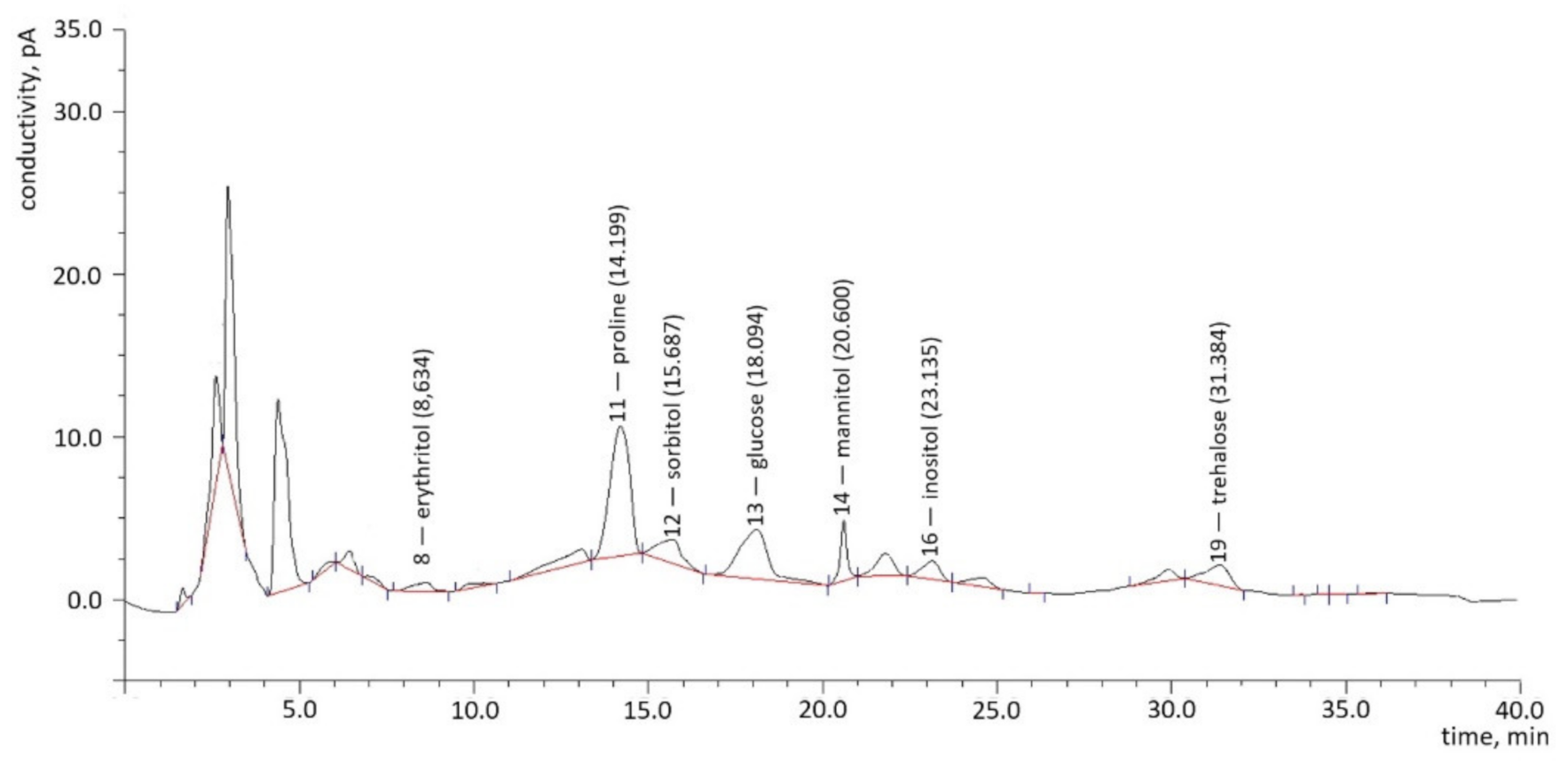
2.1.1. Chromatography
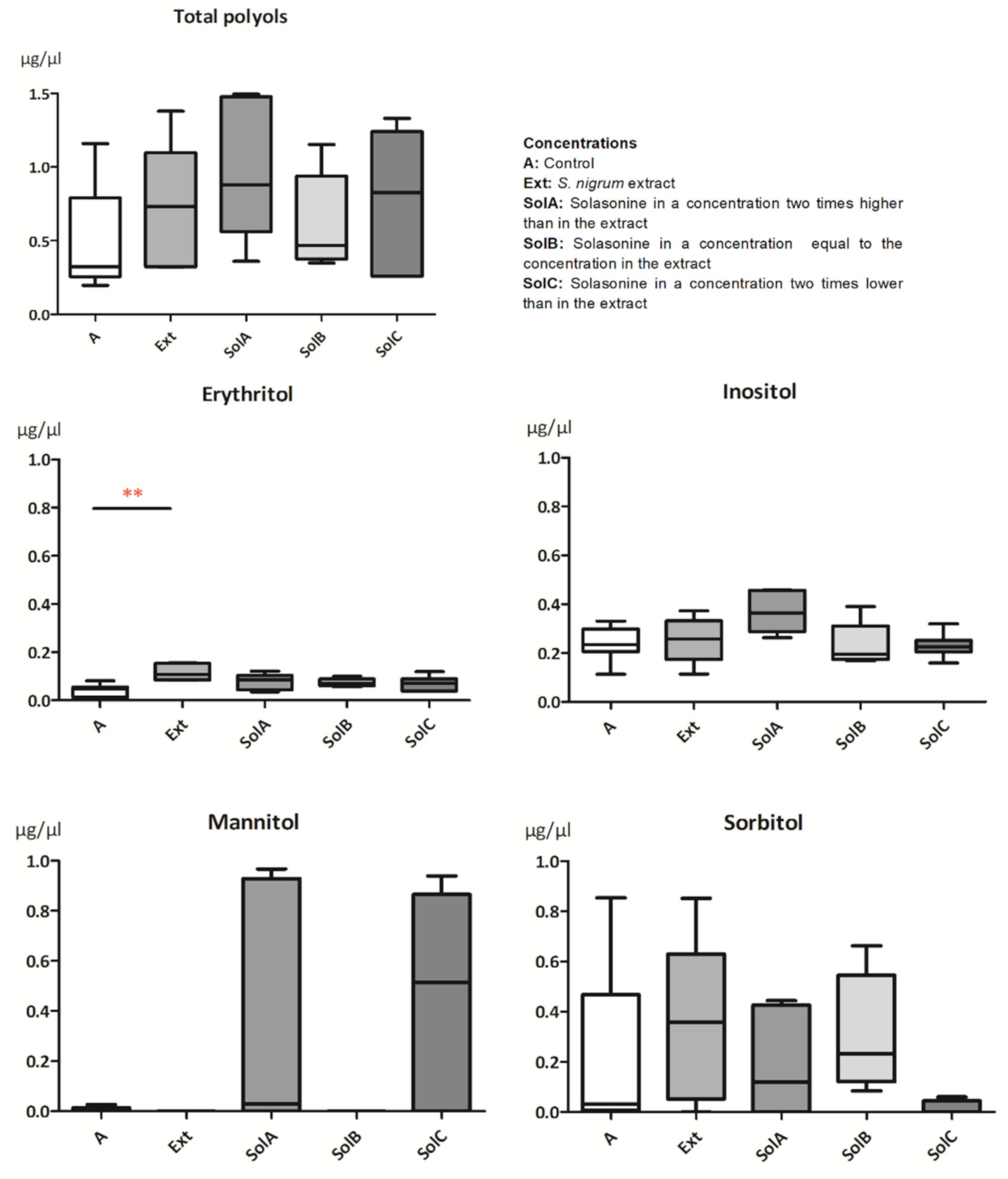
2.1.2. Polyols
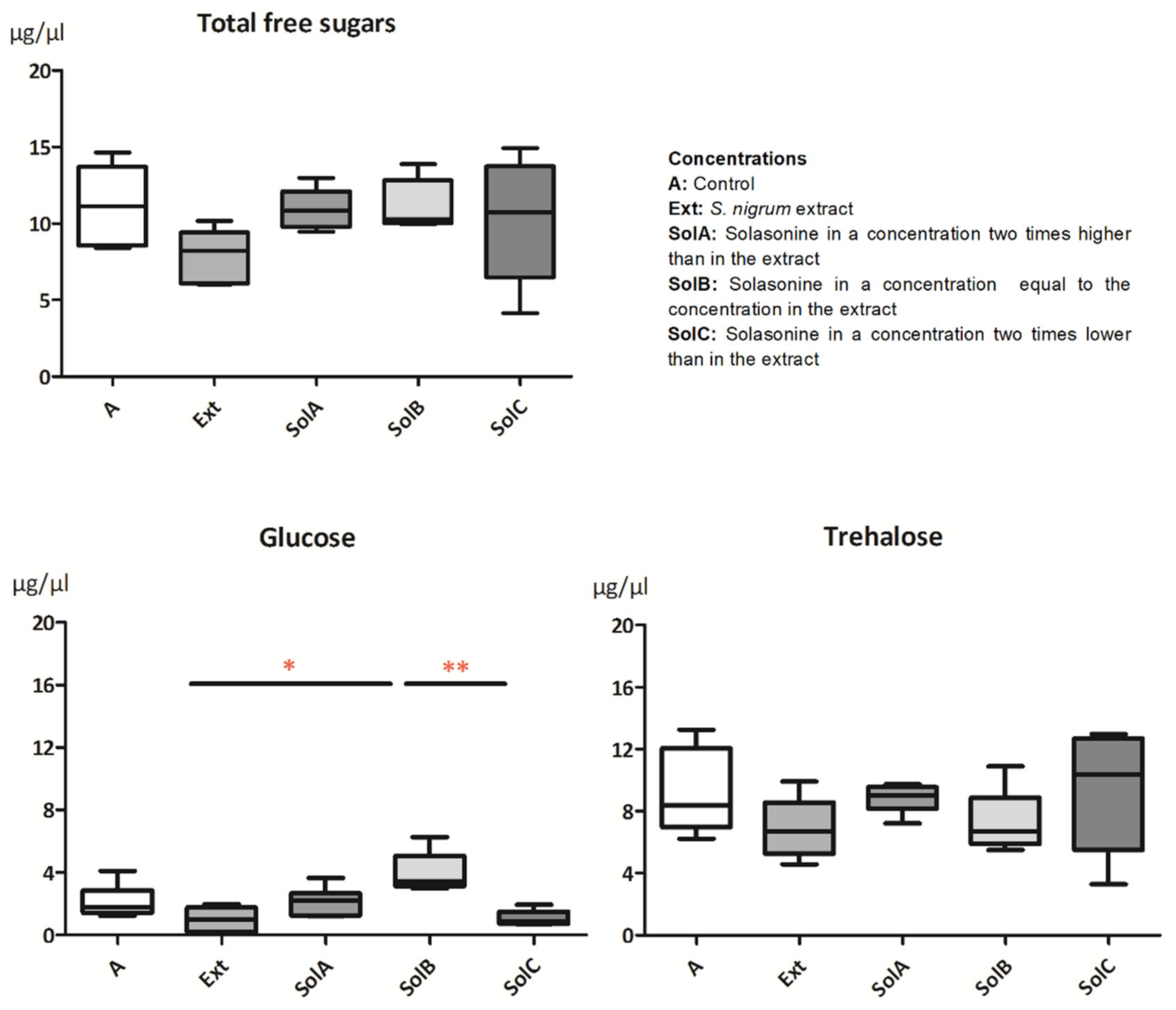
2.1.3. Sugars
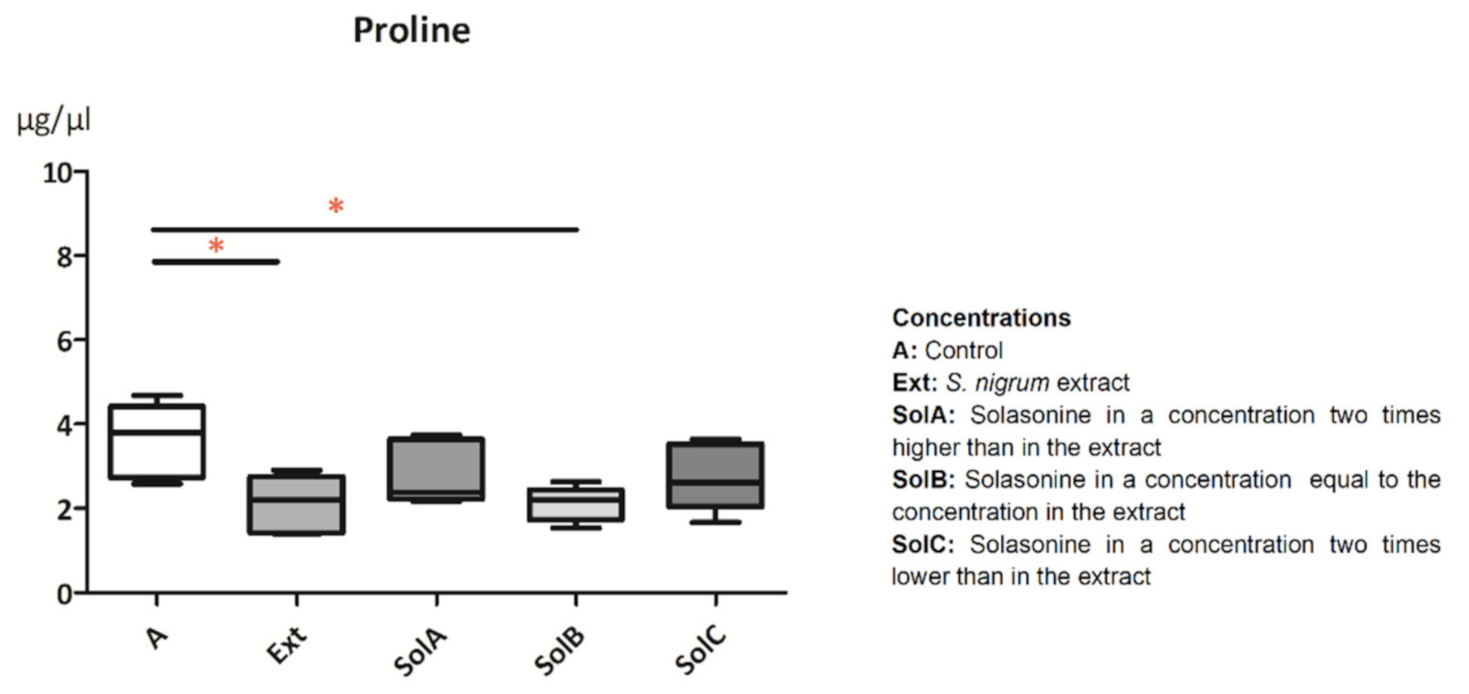
2.1.4. Proline
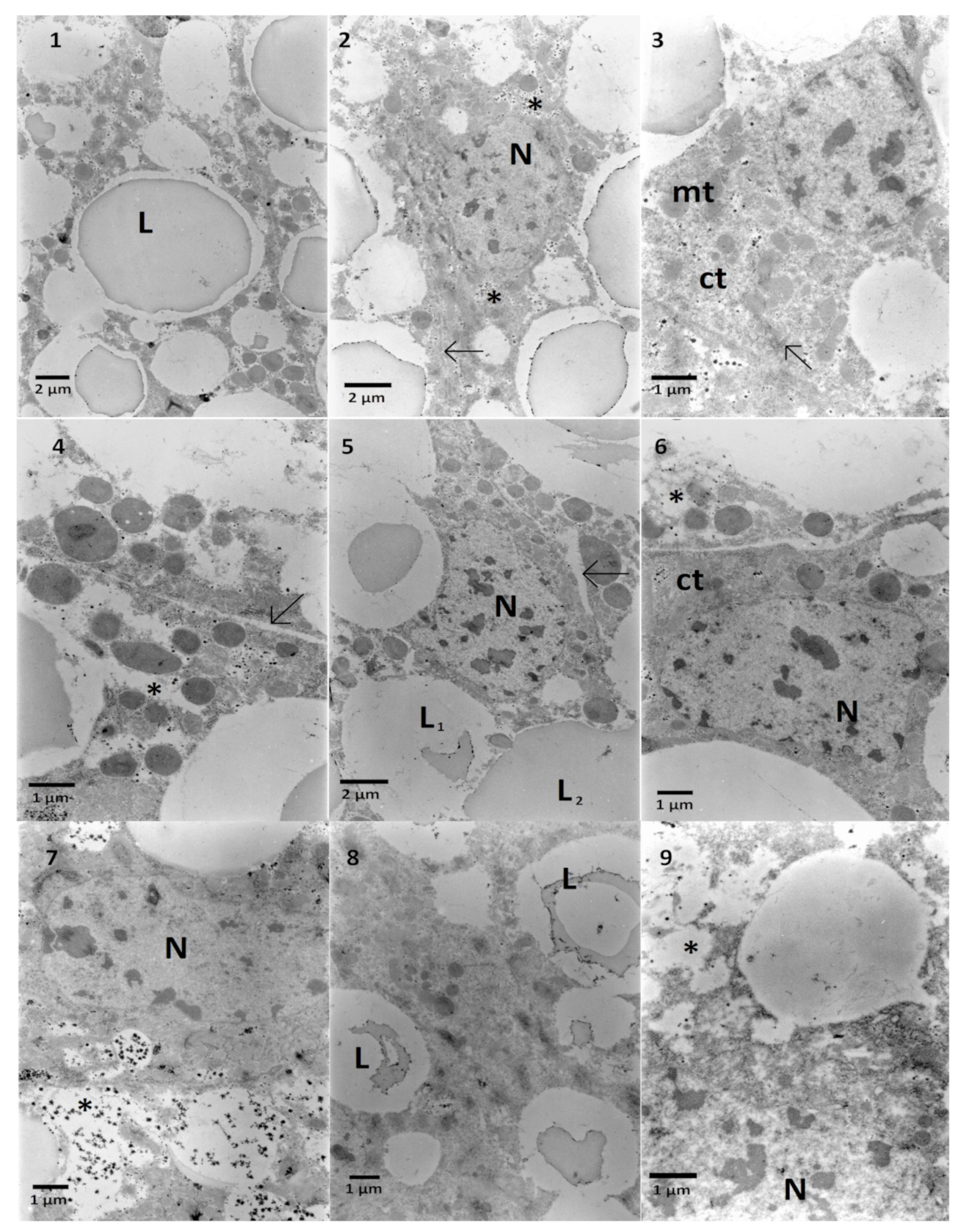
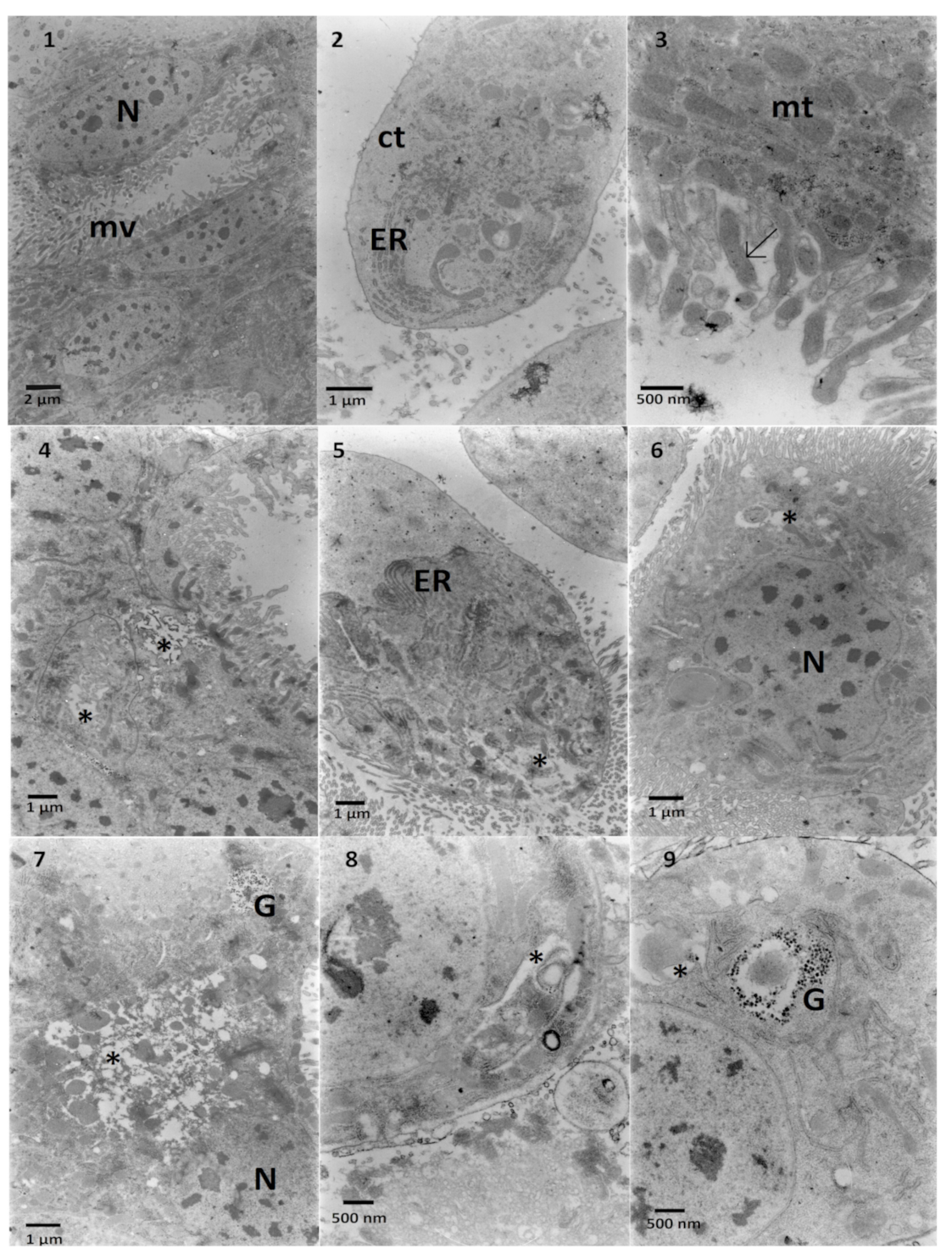
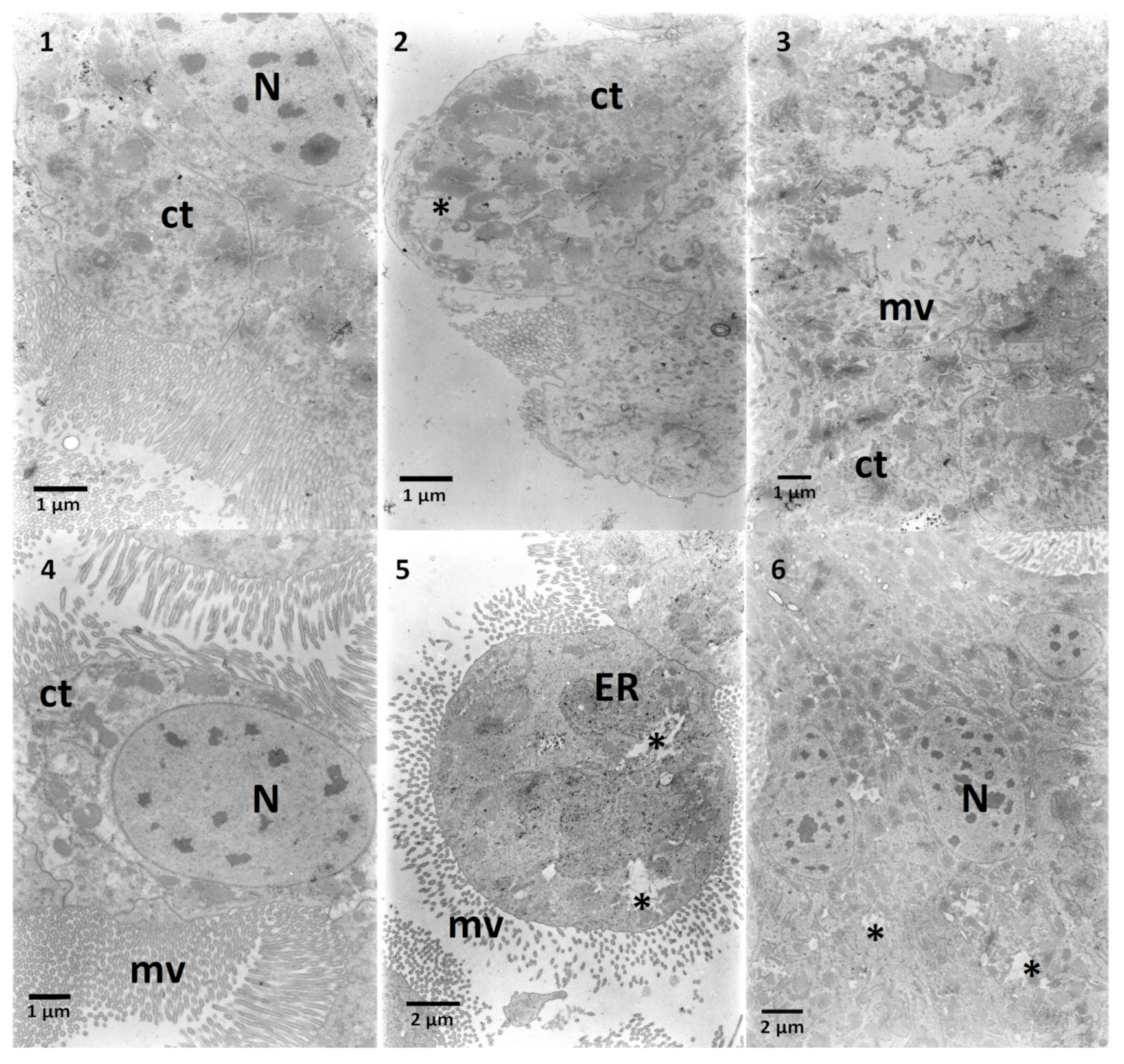
2.2. The Effects on the Ultrastructure
2.2.1. The Fat Body
2.2.2. The Midgut
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Larvae
5.2. Nourishment Preparation
5.3. Extract and Solasonine Preparation
- SolA: two times higher than the extract concentration (1.49 ×10−3 mg/1 g of the diet);
- SolB: equal to the extract concentration (7.465 × 10−4 mg/1 g of the diet);
- SolC: two times lower than the extract concentration (3.825 × 10−4 mg/1 g of the diet).
5.4. Experiment
5.5. Samples Collection for RP-HPLC
5.6. Qualitative and Quantitative Determination of Hemolymph Components
5.7. Samples Collection for TEM
5.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spochacz, M.; Chowański, S.; Walkowiak-Nowicka, K.; Szymczak, M.; Adamski, Z. Plant-derived substances used against beetles–pests of stored crops and food–and their mode of action: A review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1339–1366. [Google Scholar] [CrossRef] [Green Version]
- Spochacz, M.; Chowański, S.; Szymczak, M.; Lelario, F.; Bufo, S.A.; Adamski, Z. Sublethal Effects of Solanum nigrum Fruit Extract and Its Pure Glycoalkaloids on the Physiology of Tenebrio molitor (Mealworm). Toxins 2018, 10, 504. [Google Scholar] [CrossRef] [Green Version]
- Spochacz, M.; Szymczak, M.; Chowański, S.; Bufo, S.A.; Adamski, Z. Solanum nigrum Fruit Extract Increases Toxicity of Fenitrothion—A Synthetic Insecticide, in the Mealworm Beetle Tenebrio molitor Larvae. Toxins 2020, 12, 612. [Google Scholar] [CrossRef]
- Ntalli, N.G.; Menkissoglu-Spiroudi, U. Pesticides of botanical origin: A promising tool in plant protection. In Pesticides-Formulations, Effects, Fate; IntechOpen: London, UK, 2011; pp. 1–23. [Google Scholar]
- Marciniak, P.; Kolińska, A.; Spochacz, M.; Chowański, S.; Adamski, Z.; Scrano, L.; Falabella, P.; Bufo, S.A.; Rosiński, G. Differentiated Effects of Secondary Metabolites from Solanaceae and Brassicaceae Plant Families on the Heartbeat of Tenebrio molitor Pupae. Toxins 2019, 11, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventrella, E.; Marciniak, P.; Adamski, Z.; Rosiński, G.; Chowański, S.; Falabella, P.; Scrano, L.; Bufo, S.A. Cardioactive properties of Solanaceae plant extracts and pure glycoalkaloids on Zophobas atratus. Insect Sci. 2015, 22, 251–262. [Google Scholar] [CrossRef]
- Chowański, S.; Adamski, Z.; Marciniak, P.; Rosiński, G.; Büyükgüzel, E.; Büyükgüzel, K.; Falabella, P.; Scrano, L.; Ventrella, E.; Lelario, F. A review of bioinsecticidal activity of Solanaceae alkaloids. Toxins 2016, 8, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Büyükgüzel, E.; Büyükgüzel, K.; Snela, M.; Erdem, M.; Radtke, K.; Ziemnicki, K.; Adamski, Z. Effect of boric acid on antioxidant enzyme activity, lipid peroxidation, and ultrastructure of midgut and fat body of Galleria mellonella. Cell Biol. Toxicol. 2013, 29, 117–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamski, Z.; Marciniak, P.; Ziemnicki, K.; Büyükgüzel, E.; Erdem, M.; Büyükgüzel, K.; Ventrella, E.; Falabella, P.; Cristallo, M. Potato Leaf Extract and It’s Component, α-solanine, Exert Similar Impacts on Development and Oxidative Stress in Galleria mellonella L. Arch. Insect Biochem. Physiol. 2014, 87, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Mikulak, E.; Gliniewicz, A.; Przygodzka, M.; Solecka, J. Galleria mellonella L. as model organism used in biomedical and other studies. Przegl. Epidemiol. 2018, 72, 57–73. [Google Scholar]
- Kwadha, C.A.; Ong’amo, G.O.; Ndegwa, P.N.; Raina, S.K.; Fombong, A.T. The biology and control of the greater wax moth, Galleria mellonella. Insects 2017, 8, 61. [Google Scholar] [CrossRef] [Green Version]
- Auerswald, L.; Gäde, G. Metabolic changes in the African fruit beetle, Pachnoda sinuata, during starvation. J. Insect Physiol. 2000, 46, 343–351. [Google Scholar] [CrossRef]
- Mullins, D.E. Chemistry and Physiology of the Hemolymph. Compr. Insect Physiol. Biochem. Pharmacol. 1985, 3, 355–400. [Google Scholar]
- Słocińska, M.; Chowański, S.; Marciniak, P. Identification of sulfakinin receptors (SKR) in Tenebrio molitor beetle and the influence of sulfakinins on carbohydrates metabolism. J. Comp. Physiol. B 2020, 190, 669–679. [Google Scholar] [CrossRef]
- Martoja, R.; Ballan-Dufrançais, C. The Ultrastructure of the Digestive and Excretory Organs. Insect Ultrastruct. 1984, 2, 199–268. [Google Scholar]
- Killiny, N. Generous hosts: Why the larvae of greater wax moth, Galleria mellonella is a perfect infectious host model? Virulence 2018, 9, 860–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanzal, R.; Jegorov, A. Changes in free amino acid composition in haemolymph of larvae of the wax moth, Galleria mellonella L., during cold acclimation. Comp. Biochem. Physiol. A Physiol. 1991, 100, 957–962. [Google Scholar] [CrossRef]
- Chowanski, S.; Lubawy, J.; Spochacz, M.; Ewelina, P.; Grzegorz, S.; Rosinski, G.; Slocinska, M. Cold induced changes in lipid, protein and carbohydrate levels in the tropical insect Gromphadorhina coquereliana. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2015, 183, 57–63. [Google Scholar] [CrossRef]
- Kim, M.; Lee, S.; Chun, Y.S.; Na, J.; Kwon, H.; Kim, W.; Kim, Y. Heat tolerance induction of the Indian meal moth (Lepidoptera: Pyralidae) is accompanied by upregulation of heat shock proteins and polyols. Environ. Entomol. 2017, 46, 1005–1011. [Google Scholar] [CrossRef]
- Yang, S.; Zhao, Y.; Yu, J.; Fan, Z.; Gong, S.-T.; Tang, H.; Pan, L. Sugar alcohols of polyol pathway serve as alarmins to mediate local-systemic innate immune communication in Drosophila. Cell Host Microbe 2019, 26, 240–251.e8. [Google Scholar] [CrossRef]
- Munro, I.; Bernt, W.; Borzelleca, J.; Flamm, G.; Lynch, B.; Kennepohl, E.; Bär, E.; Modderman, J. Erythritol: An interpretive summary of biochemical, metabolic, toxicological and clinical data. Food Chem. Toxicol. 1998, 36, 1139–1174. [Google Scholar] [CrossRef]
- Tang, S.B.; Lee, J.C.; Jung, J.K.; Choi, M.-Y. Effect of erythritol formulation on the mortality, fecundity and physiological excretion in Drosophila suzukii. J. Insect Physiol. 2017, 101, 178–184. [Google Scholar] [CrossRef]
- Den Hartog, G.J.; Boots, A.W.; Adam-Perrot, A.; Brouns, F.; Verkooijen, I.W.; Weseler, A.R.; Haenen, G.R.; Bast, A. Erythritol is a sweet antioxidant. Nutrition 2010, 26, 449–458. [Google Scholar] [CrossRef]
- Roddick, J.G.; Rijnenberg, A.L.; Weissenberg, M. Membrane-disrupting properties of the steroidal glycoalkaloids solasonine and solamargine. Phytochemistry 1990, 29, 1513–1518. [Google Scholar] [CrossRef]
- Klowden, M.J. Physiological Systems in Insects, 2nd ed.; Academic Press: Cambridge, MA, USA, 2008; pp. 357–401. ISBN 978-012369493-5. [Google Scholar]
- Wyatt, G.; Kalf, G. The Chemistry of Insect Hemolymph II. Trehalose and Other Carbohydrates. J. Gen. Physiol. 1957, 40, 833–847. [Google Scholar] [CrossRef]
- Boctor, I. Carbohydrates in the Haemolymph of the Prepupal and Pupal Stages of Spodoptera littoralis Boisduval. J. Comp. Physiol. 1974, 94, 353–356. [Google Scholar] [CrossRef]
- Vanin, S.; Bubacco, L.; Beltramini, M. Seasonal variation of trehalose and glycerol concentrations in winter snow-active insects. CryoLetters 2008, 29, 485–491. [Google Scholar] [PubMed]
- Thompson, S.N. Trehalose—The Insect ‘Blood’ Sugar. Adv. Insect Physiol. 2003, 31, 205–285. [Google Scholar]
- Niimi, N.; Yako, H.; Takaku, S.; Chung, S.K.; Sango, K. Aldose Reductase and the Polyol Pathway in Schwann Cells: Old and New Problems. Int. J. Mol. Sci. 2021, 22, 1031. [Google Scholar] [CrossRef] [PubMed]
- Bursell, E. The Role of Proline in Energy Metabolism. In Energy Metabolism in Insects; Springer: Boston, MA, USA, 1981; pp. 135–154. [Google Scholar]
- Adamski, Z.; Radtke, K.; Kopiczko, A.; Chowański, S.; Marciniak, P.; Szymczak, M.; Spochacz, M.; Falabella, P.; Lelario, F.; Scrano, L. Ultrastructural and developmental toxicity of potato and tomato leaf extracts to beet armyworm, Spodoptera exigua (Lepidoptera: Noctuidae). Microsc. Res. Tech. 2016, 79, 948–958. [Google Scholar] [CrossRef]
- Siddique, M.A.B.; Brunton, N. Food Glycoalkaloids: Distribution, Structure, Cytotoxicity, Extraction, and Biological Activity. In Alkaloids-Their Importance in Nature and Human Life; IntechOpen: London, UK, 2019; pp. 13–15. [Google Scholar] [CrossRef] [Green Version]
- Hasanain, M.; Bhattacharjee, A.; Pandey, P.; Ashraf, R.; Singh, N.; Sharma, S.; Vishwakarma, A.; Datta, D.; Mitra, K.; Sarkar, J. α-Solanine induces ROS-mediated autophagy through activation of endoplasmic reticulum stress and inhibition of Akt/mTOR pathway. Cell Death Dis. 2015, 6, e1860. [Google Scholar] [CrossRef] [Green Version]
- Nepal, B.; Stine, K.J. Glycoalkaloids: Structure, Properties, and Interactions with Model Membrane Systems. Processes 2019, 7, 513. [Google Scholar] [CrossRef] [Green Version]
- Keukens, E.A.; de Vrije, T.; van den Boom, C.; de Waard, P.; Plasman, H.H.; Thiel, F.; Chupin, V.; Jongen, W.M.; de Kruijff, B. Molecular basis of glycoalkaloid induced membrane disruption. BBA-Biomembr. 1995, 1240, 216–228. [Google Scholar] [CrossRef] [Green Version]
- Kasiotis, K.M.; Evergetis, E.; Papachristos, D.; Vangelatou, O.; Antonatos, S.; Milonas, P.; Haroutounian, S.A.; Machera, K. An essay on ecosystem availability of Nicotiana glauca Graham alkaloids: The honeybees case study. BMC Ecol. 2020, 20, 57. [Google Scholar] [CrossRef] [PubMed]
- Du Rand, E.E.; Smit, S.; Beukes, M.; Apostolides, Z.; Pirk, C.W.W.; Nicolson, S.W. Detoxification mechanisms of honey bees (Apis mellifera) resulting in tolerance of dietary nicotine. Sci. Rep. 2015, 5, 11779. [Google Scholar] [CrossRef] [Green Version]
- Campadelli, G. Allevamento di Galleria mellonella L. (Lepidoptera: Galleriidae) con dieta semiartificiale. Boll. Dell’istituto Entomol. Univ. Studi Bologna 1973, 32, 11–25. [Google Scholar]
- Lelario, F.; De Maria, S.; Rivelli, A.R.; Russo, D.; Milella, L.; Bufo, S.A.; Scrano, L. A Complete Survey of Glycoalkaloids Using LC-FTICR-MS and IRMPD in a Commercial Variety and a Local Landrace of Eggplant (Solanum melongena L.) and their Anticholinesterase and Antioxidant Activities. Toxins 2019, 11, 230. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proline | Sorbitol | Mannitol | Erythritol | Inositol | Glucose | Trehalose | ||
|---|---|---|---|---|---|---|---|---|
| Ext | ↓ * | - | ↓ | ↑ * | - | ↓ |  | ↓ |
| SolA (2x↑Ext) | ↓ | - | ↑ | ↑ | ↑ | - | - | |
| SolB (-II- Ext) | ↓ * | - | ↓ | ↑ | - | ↑ | - | |
| SolC (2x↓Ext) | ↓ | ↓ | ↑ | ↑ | - | ↓ | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spochacz, M.; Chowański, S.; Szymczak-Cendlak, M.; Marciniak, P.; Lelario, F.; Salvia, R.; Nardiello, M.; Scieuzo, C.; Scrano, L.; Bufo, S.A.; et al. Solanum nigrum Extract and Solasonine Affected Hemolymph Metabolites and Ultrastructure of the Fat Body and the Midgut in Galleria mellonella. Toxins 2021, 13, 617. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090617
Spochacz M, Chowański S, Szymczak-Cendlak M, Marciniak P, Lelario F, Salvia R, Nardiello M, Scieuzo C, Scrano L, Bufo SA, et al. Solanum nigrum Extract and Solasonine Affected Hemolymph Metabolites and Ultrastructure of the Fat Body and the Midgut in Galleria mellonella. Toxins. 2021; 13(9):617. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090617
Chicago/Turabian StyleSpochacz, Marta, Szymon Chowański, Monika Szymczak-Cendlak, Paweł Marciniak, Filomena Lelario, Rosanna Salvia, Marisa Nardiello, Carmen Scieuzo, Laura Scrano, Sabino A. Bufo, and et al. 2021. "Solanum nigrum Extract and Solasonine Affected Hemolymph Metabolites and Ultrastructure of the Fat Body and the Midgut in Galleria mellonella" Toxins 13, no. 9: 617. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090617