ExlA Pore-Forming Toxin: Localization at the Bacterial Membrane, Regulation of Secretion by Cyclic-Di-GMP, and Detection In Vivo

 , , and
, , and
Abstract
:1. Introduction
2. Results
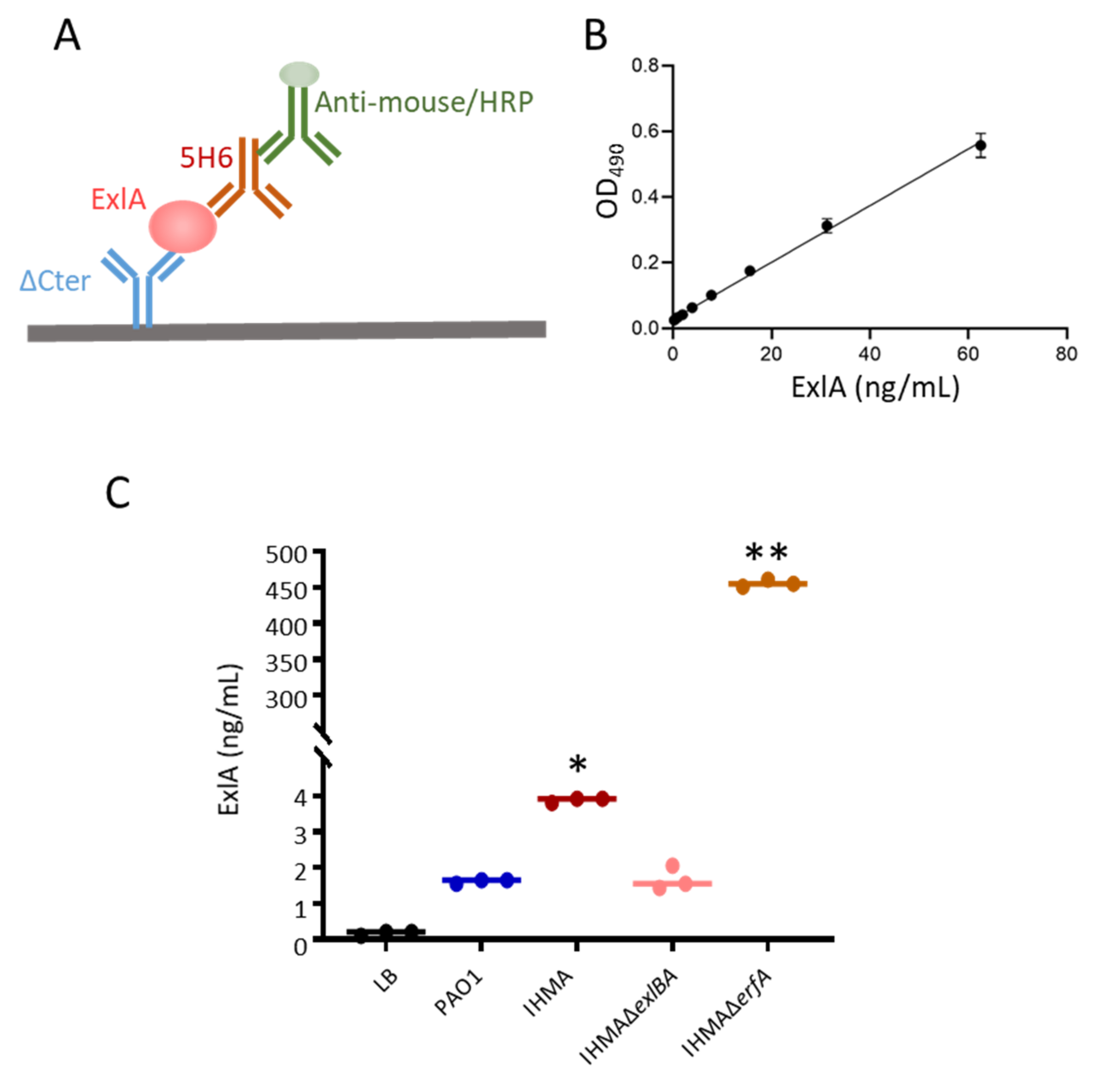
2.1. ExlA Quantification in Bacterial Secretomes
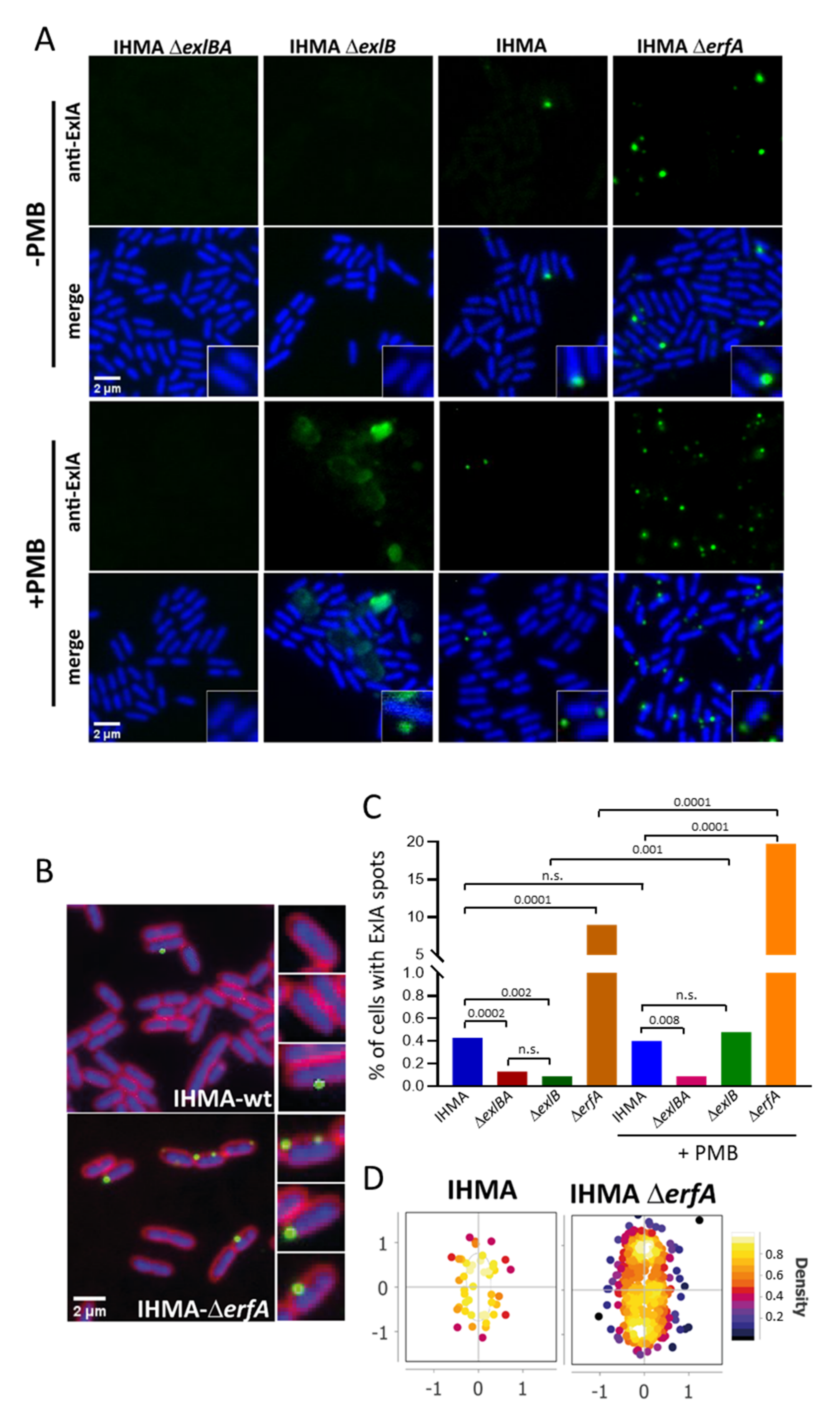
2.2. ExlA Localization at the Bacterial Membrane
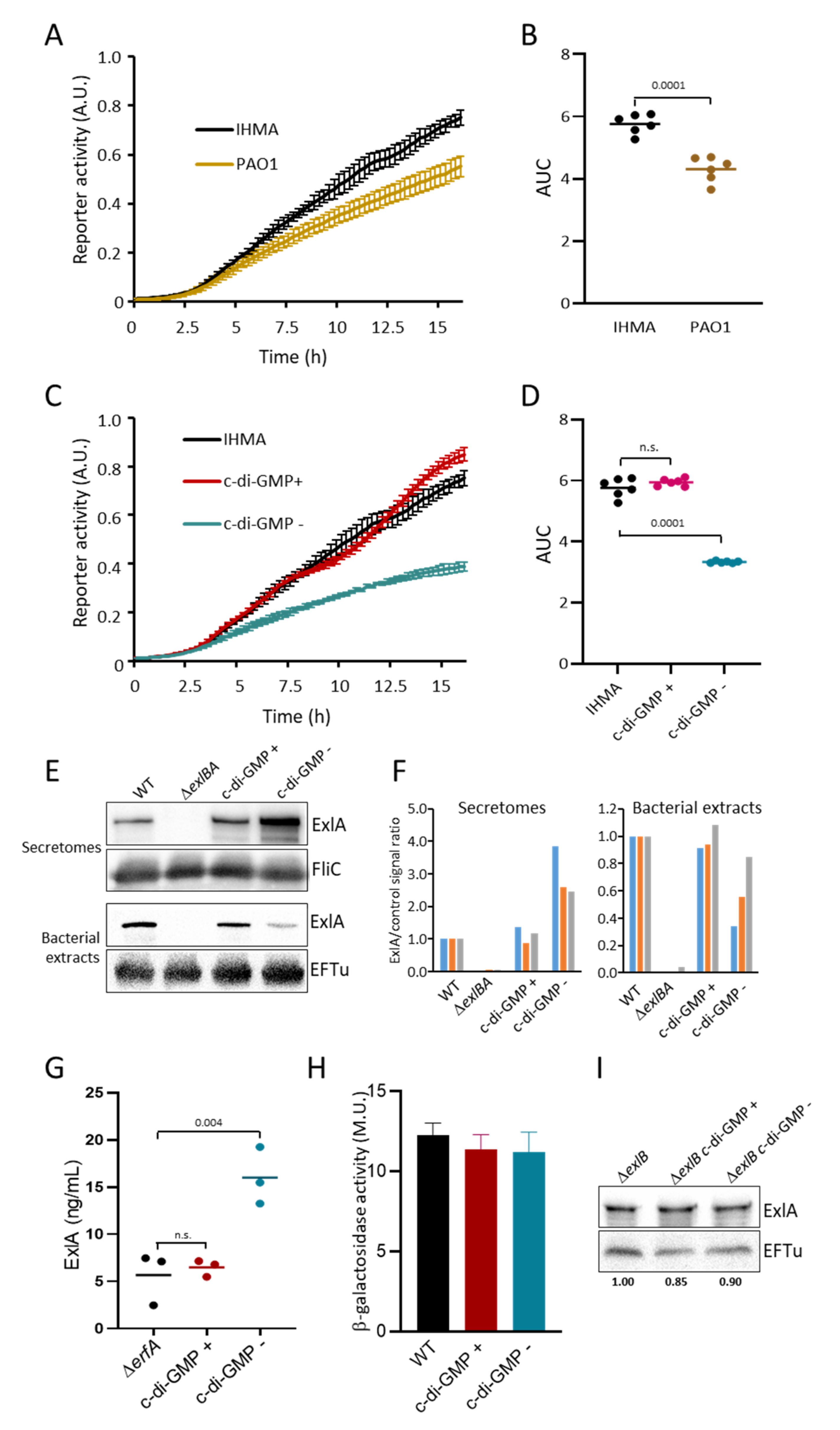
2.3. Cyclic-di-GMP Controls ExlA Secretion
2.4. ExlA Is Retained at the Outer Membrane
2.5. ExlA Detection in Broncho-Alveolar Lavage Fluids of Infected Mice
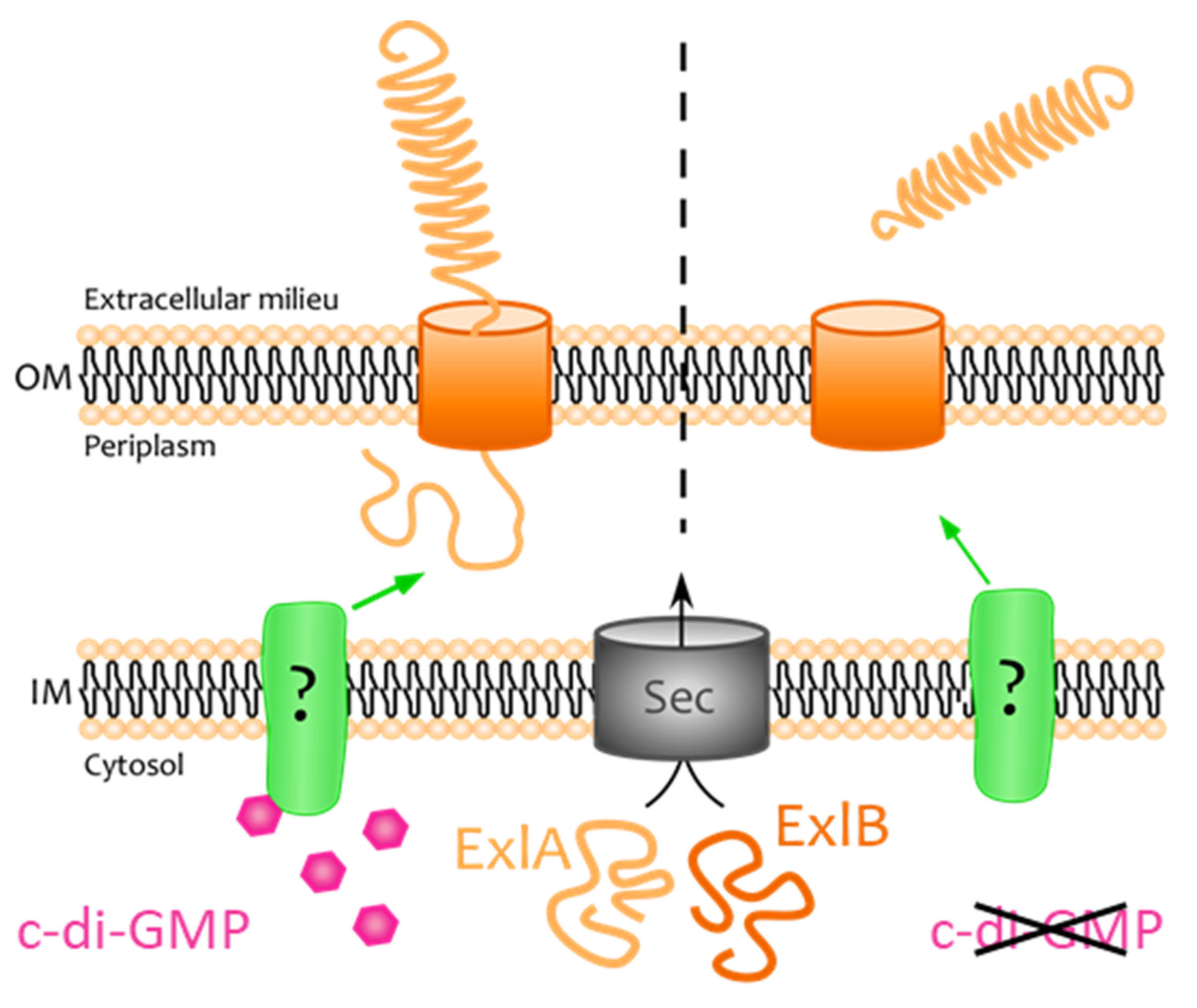
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. E. Coli and P. Aeruginosa Strains and Culture
4.3. Cloning
4.4. Production and Purification of ExlA and ExlA Truncated Derivatives
4.5. Production of Monoclonal Antibodies
4.6. Antibody Mapping on ExlA Sequence
4.7. ExlA ELISA
4.8. Production of Bacterial Secretomes
4.9. Immunofluorescence of Bacteria
4.10. Fluorescence of GFP Bacteria
4.11. Total Protein Extracts from Secretomes and Bacteria
4.12. Cellular Fractionation
4.13. Proteinase K Treatment
4.14. Western Blot
4.15. β-Galactosidase Activity
4.16. Mouse Pulmonary Infection
4.17. Analysis of BALs
4.18. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gellatly, S.L.; Hancock, R.E.W. Pseudomonas aeruginosa: New insights into pathogenesis and host defenses. Pathog. Dis. 2013, 67, 159–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, B.J.; Dehnbostel, J.; Blackwell, T.S. Pseudomonas aeruginosa: Host defence in lung diseases. Respirology 2010, 15, 1037–1056. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.R. The type III secretion system of Pseudomonas aeruginosa: Infection by injection. Nat. Rev. Genet. 2009, 7, 654–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsen, S.; Huber, P.; Bouillot, S.; Couté, Y.; Fournier, P.; Dubois, Y.; Timsit, J.-F.; Maurin, M.; Attrée, I. A Type III Secretion Negative Clinical Strain of Pseudomonas aeruginosa Employs a Two-Partner Secreted Exolysin to Induce Hemorrhagic Pneumonia. Cell Host Microbe 2014, 15, 164–176. [Google Scholar] [CrossRef] [Green Version]
- Huber, P.; Basso, P.; Reboud, E.; Attrée, I. Pseudomonas aeruginosa renews its virulence factors. Environ. Microbiol. Rep. 2016, 8, 564–571. [Google Scholar] [CrossRef]
- Reboud, E.; Elsen, S.; Bouillot, S.; Golovkine, G.; Basso, P.; Jeannot, K.; Attree, I.; Huber, P. Phenotype and toxicity of the recently discovered exlA -positive Pseudomonas aeruginosa strains collected worldwide. Environ. Microbiol. Rep. 2016, 18, 3425–3439. [Google Scholar] [CrossRef]
- Roy, P.H.; Tetu, S.; Larouche, A.; Elbourne, L.; Tremblay, S.; Ren, Q.; Dodson, R.; Harkins, D.; Shay, R.; Watkins, K.; et al. Complete Genome Sequence of the Multiresistant Taxonomic Outlier Pseudomonas aeruginosa PA7. PLoS ONE 2010, 5, e8842. [Google Scholar] [CrossRef]
- Basso, P.; Ragno, M.; Elsen, S.; Reboud, E.; Golovkine, G.; Bouillot, S.; Huber, P.; Lory, S.; Faudry, E.; Attrée, I. Pseudomonas aeruginosa Pore-Forming Exolysin and Type IV Pili Cooperate To Induce Host Cell Lysis. mBio 2017, 8, e02250-16. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, Q.; Job, V.; Maillard, A.P.; Imbert, L.; Teulon, J.-M.; Favier, A.; Pellequer, J.-L.; Huber, P.; Attrée, I.; Dessen, A. Exolysin (ExlA) from Pseudomonas aeruginosa Punctures Holes into Target Membranes Using a Molten Globule Domain. J. Mol. Biol. 2020, 432, 4466–4480. [Google Scholar] [CrossRef]
- Basso, P.; Wallet, P.; Elsen, S.; Soleilhac, E.; Henry, T.; Faudry, E.; Attrée, I. Multiple Pseudomonas species secrete exolysin-like toxins and provoke Caspase-1-dependent macrophage death. Environ. Microbiol. Rep. 2017, 19, 4045–4064. [Google Scholar] [CrossRef]
- Vesga, P.; Flury, P.; Vacheron, J.; Keel, C.; Croll, D.; Maurhofer, M. Transcriptome plasticity underlying plant root colonization and insect invasion by Pseudomonas protegens. ISME J. 2020, 14, 2766–2782. [Google Scholar] [CrossRef] [PubMed]
- Guérin, J.; Bigot, S.; Schneider, R.; Buchanan, S.K.; Jacob-Dubuisson, F. Two-Partner Secretion: Combining Efficiency and Simplicity in the Secretion of Large Proteins for Bacteria-Host and Bacteria-Bacteria Interactions. Front. Cell. Infect. Microbiol. 2017, 7, 148. [Google Scholar] [CrossRef] [PubMed]
- Reboud, E.; Bouillot, S.; Patot, S.; Béganton, B.; Attrée, I.; Huber, P. Pseudomonas aeruginosa ExlA and Serratia marcescens ShlA trigger cadherin cleavage by promoting calcium influx and ADAM10 activation. PLoS Pathog. 2017, 13, e1006579. [Google Scholar] [CrossRef] [PubMed]
- Berry, A.; Han, K.; Trouillon, J.; Robert-Genthon, M.; Ragno, M.; Lory, S.; Attrée, I.; Elsen, S. cAMP and Vfr Control Exolysin Expression and Cytotoxicity of Pseudomonas aeruginosa Taxonomic Outliers. J. Bacteriol. 2018, 200, e00135-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trouillon, J.; Sentausa, E.; Ragno, M.; Robert-Genthon, M.; Lory, S.; Attrée, I.; Elsen, S. Species-specific recruitment of transcription factors dictates toxin expression. Nucleic Acids. Res. 2020, 48, 2388–2400. [Google Scholar] [CrossRef] [Green Version]
- Bouillot, S.; Munro, P.; Gallet, B.; Reboud, E.; Cretin, F.; Golovkine, G.; Schoehn, G.; Attrée, I.; Lemichez, E.; Huber, P. Pseudomonas aeruginosa Exolysin promotes bacterial growth in lungs, alveolar damage and bacterial dissemination. Sci. Rep. 2017, 7, 2120. [Google Scholar] [CrossRef]
- Bouillot, S.; Pont, S.; Gallet, B.; Moriscot, C.; Deruelle, V.; Attrée, I.; Huber, P. Inflammasome activation by Pseudomonas aeruginosa’s ExlA pore-forming toxin is detrimental for the host. Cell. Microbiol. 2020, 22, 13251. [Google Scholar] [CrossRef]
- Freschi, L.; Vincent, A.T.; Jeukens, J.; Emond-Rheault, J.-G.; Kukavica-Ibrulj, I.; Dupont, M.-J.; Charette, S.J.; Boyle, B.; Levesque, R.C. The Pseudomonas aeruginosa Pan-Genome Provides New Insights on Its Population Structure, Horizontal Gene Transfer, and Pathogenicity. Genome Biol. Evol. 2019, 11, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Medina-Rojas, M.; Stribling, W.; Snesrud, E.; Garry, B.I.; Li, Y.; Mc Gann, P.; Demons, S.T.; Tyner, S.D.; Zurawski, D.V.; Antonic, V. Comparison of Pseudomonas aeruginosa strains reveals that Exolysin A toxin plays an additive role in virulence. Pathog. Dis. 2020, 78, ftaa010. [Google Scholar] [CrossRef]
- Ruiz-Roldán, L.; Rojo-Bezares, B.; De Toro, M.; López, M.; Toledano, P.; Lozano, C.; Chichón, G.; Alvarez-Erviti, L.; Torres, C.; Sáenz, Y. Antimicrobial resistance and virulence of Pseudomonas spp. among healthy animals: Concern about exolysin ExlA detection. Sci. Rep. 2020, 10, 11667. [Google Scholar] [CrossRef]
- Ruhe, Z.C.; Subramanian, P.; Song, K.; Nguyen, J.Y.; Stevens, T.A.; Low, D.A.; Jensen, G.J.; Hayes, C.S. Programmed Secretion Arrest and Receptor-Triggered Toxin Export during Antibacterial Contact-Dependent Growth Inhibition. Cell 2018, 175, 921–933.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Argenio, D.A.; Miller, S.I. Cyclic di-GMP as a bacterial second messenger. Microbiology 2004, 150, 2497–2502. [Google Scholar] [CrossRef] [Green Version]
- Rybtke, M.T.; Borlee, B.R.; Murakami, K.; Irie, Y.; Hentzer, M.; Nielsen, T.E.; Givskov, M.; Parsek, M.R.; Tolker-Nielsen, T. Fluorescence-Based Reporter for Gauging Cyclic Di-GMP Levels in Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2012, 78, 5060–5069. [Google Scholar] [CrossRef] [Green Version]
- Baker, A.E.; Diepold, A.; Kuchma, S.L.; Scott, J.E.; Ha, D.G.; Orazi, G.; Armitage, J.P.; O’Toole, G.A. PilZ Domain Protein FlgZ Mediates Cyclic Di-GMP-Dependent Swarming Motility Control in Pseudomonas aeruginosa. J. Bacteriol. 2016, 198, 1837–1846. [Google Scholar] [CrossRef] [Green Version]
- Valentini, M.; Filloux, A. Biofilms and Cyclic di-GMP (C-di-GMP) Signaling: Lessons from Pseudomonas aeruginosa and Other Bacteria. J. Biol. Chem. 2016, 291, 12547–12555. [Google Scholar] [CrossRef] [Green Version]
- Moscoso, J.A.; Mikkelsen, H.; Heeb, S.; Williams, P.; Filloux, A. The Pseudomonas aeruginosa sensor RetS switches Type III and Type VI secretion via c-di-GMP signalling. Environ. Microbiol. Rep. 2011, 13, 3128–3138. [Google Scholar] [CrossRef] [PubMed]
- Borlee, B.R.; Goldman, A.D.; Murakami, K.; Samudrala, R.; Wozniak, D.J.; Parsek, M.R. Pseudomonas aeruginosa uses a cyclic-di-GMP-regulated adhesin to reinforce the biofilm extracellular matrix. Mol. Microbiol. 2010, 75, 827–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dal Peraro, M.; van der Goot, F.G. Pore-forming toxins: Ancient, but never really out of fashion. Nat. Rev. Microbiol. 2016, 14, 77–92. [Google Scholar] [CrossRef]
- Di Venanzio, G.; Stepanenko, T.M.; Véscovi, E.G. Serratia marcescens ShlA Pore-Forming Toxin Is Responsible for Early Induction of Autophagy in Host Cells and Is Transcriptionally Regulated by RcsB. Infect. Immun. 2014, 82, 3542–3554. [Google Scholar] [CrossRef] [Green Version]
- Grass, S.; Rempe, K.A.; Geme, J.W.S. Structural Determinants of the Interaction between the TpsA and TpsB Proteins in the Haemophilus influenzae HMW1 Two-Partner Secretion System. J. Bacteriol. 2015, 197, 1769–1780. [Google Scholar] [CrossRef] [Green Version]
- Cooley, R.B.; Smith, T.J.; Leung, W.; Tierney, V.; Borlee, B.R.; O’Toole, G.A.; Sondermann, H. Cyclic Di-GMP-Regulated Periplasmic Proteolysis of a Pseudomonas aeruginosa Type Vb Secretion System Substrate. J. Bacteriol. 2016, 198, 66–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nash, Z.M.; Cotter, P.A. Bordetella Filamentous Hemagglutinin, a Model for the Two-Partner Secretion Pathway. Microbiol. Spectr. 2019, 7, 3542–3554. [Google Scholar] [CrossRef]
- Kos, V.N.; Déraspe, M.; McLaughlin, R.E.; Whiteaker, J.D.; Roy, P.H.; Alm, R.A.; Corbeil, J.; Gardner, H. The Resistome of Pseudomonas aeruginosa in Relationship to Phenotypic Susceptibility. Antimicrob. Agents Chemother. 2015, 59, 427–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, B.; de Lorenzo, V.; Timmis, K.N. A general system to integrate lacZ fusions into the chromosomes of gram-negative eubacteria: Regulation of the Pm promoter of the TOL plasmid studied with all controlling elements in monocopy. Mol. Gen. Genet. 1992, 233, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Baynham, P.J.; Ramsey, D.M.; Gvozdyev, B.V.; Cordonnier, E.M.; Wozniak, D.J. The Pseudomonas aeruginosa Ribbon-Helix-Helix DNA-Binding Protein AlgZ (AmrZ) Controls Twitching Motility and Biogenesis of Type IV Pili. J. Bacteriol. 2006, 188, 132–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairman, J.W.; Dautin, N.; Wojtowicz, D.; Liu, W.; Noinaj, N.; Barnard, T.J.; Udho, E.; Przytycka, T.M.; Cherezov, V.; Buchanan, S.K. Crystal Structures of the Outer Membrane Domain of Intimin and Invasin from Enterohemorrhagic E. coli and Enteropathogenic Y. pseudotuberculosis. Structure 2012, 20, 1233–1243. [Google Scholar] [CrossRef] [Green Version]
- Ducret, A.; Quardokus, E.; Brun, Y.V. MicrobeJ, a tool for high throughput bacterial cell detection and quantitative analysis. Nat. Microbiol. 2016, 1, 16077. [Google Scholar] [CrossRef] [Green Version]
- Faure, L.; Garvis, S.; De Bentzmann, S.; Bigot, S. Characterization of a novel two-partner secretion system implicated in the virulence of Pseudomonas aeruginosa. Microbiology 2014, 160, 1940–1952. [Google Scholar] [CrossRef]
- Sall, K.M.; Casabona, M.G.; Bordi, C.; Huber, P.; De Bentzmann, S.; Attree, I.; Elsen, S. A gacS Deletion in Pseudomonas aeruginosa Cystic Fibrosis Isolate CHA Shapes Its Virulence. PLoS ONE 2014, 9, e95936. [Google Scholar] [CrossRef]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1972. [Google Scholar]
- Thibault, J.; Faudry, E.; Ebel, C.; Attree, I.; Elsen, S. Anti-activator ExsD Forms a 1:1 Complex with ExsA to Inhibit Transcription of Type III Secretion Operons. J. Biol. Chem. 2009, 284, 15762–15770. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Names | Relevant Characteristics | Reference/Source |
|---|---|---|
| E. coli strain | ||
| BL21 Star™ (DE3) | IPTG-inducible production of T7 RNA polymerase (prophage DE3), increased mRNA stability (rne131) and increased protein stability (lacks the Lon and OmpT proteases) | Life Technologies 3537305 10411735 |
| P. aeruginosa strains | ||
| IHMA879472 (“IHMA”) | exlA positive | [22] |
| IHMAΔexlBA | IHMA isogenic mutant deleted in exlB and exlA | [15] |
| IHMAΔerfA | IHMA isogenic mutant deficient in erfA | [15] |
| IHMA::pSW196 | IHMA with empty pSW196 vector integrated at chromosomal attB site | This study |
| IHMA::pSW196-wspR* (“c-di-GMP +”) | IHMA expressing WspR* | This study |
| IHMA::pSW196-PA2133 (“c-di-GMP −”) | IHMA expressing PA2133 | This study |
| IHMAΔerfA::pSW196 | IHMA isogenic mutant deficient in erfA with empty pSW196 vector integrated at chromosomal attB site | This study |
| IHMAΔerfA::pSW196-wspR* | IHMA isogenic mutant deficient in erfA expressing WspR* | This Study |
| IHMAΔerfA::pSW196-PA2133 | IHMA isogenic mutant deficient in erfA expressing PA2133 | This Study |
| IHMA::pSW196:Tn7-pcdrA-gfp(ASV)c | IHMA::pSW196 tagged with miniTn7 harboring PcdrA-gfp reporter fusion | This study |
| IHMA::pSW196-wspR*: Tn7-pcdrA-gfp(ASV)c | IHMA-c-di-GMP + with chromosomal PcdrA-gfp fusion | This study |
| IHMA::pSW196-PA2133: Tn7-pcdrA-gfp(ASV)c | IHMA-c-di-GMP − with chromosomal PcdrA-gfp fusion | This study |
| IHMA exlA::lacZ | IHMA87 with promoterless lacZ in exlA | [15] |
| IHMA exlA::lacZ:pSW196 | IHMA exlA::lacZ with integrated pSW196 | This study |
| IHMA exlA::lacZ:pSW196-wspR* | IHMA exlA::lacZ expressing wspR* | This study |
| IHMA exlA::lacZ:pSW196-PA2133 | IHMA exlA::lacZ expressing PA2133 | This study |
| IHMA exlB-mut | IHMA with suicide pEXG2 plasmid inserted in exlB | [8] |
| IHMAexlB-mut::pSW196-wspR* | IHMA exlB-mut expressing WspR* | This study |
| IHMAexlB-mut::pSW196-PA2133 | IHMA exlB-mut expressing PA2133 | This study |
| PAO1 | exlA negative, T3SS positive | Reference strain |
| PAO1:: pSW196:Tn7-pcdrA-gfp(ASV)c | PAO1::pSW196 tagged with miniTn7 harboring PcdrA-gfp reporter fusion | This study |
| Bacterial plasmids | ||
| pACYC-ompA-exlB | IPTG-inducible expression of codon-optimized exlB (E. coli) with signal peptide coding sequence replaced by that of E. coli ompA | This study |
| pET28-pelB-exlA | IPTG-inducible expression of codon-optimized exlA (E. coli) with signal peptide coding sequence replaced by that of D. dadantii pelB | [9] |
| pBBR1MCS4- R246A-wspR | Replicative pBBR1MCS-4 producing the highly active diguanylate cyclase WspRR246A (WspR*) | [23] |
| pBBR1MCS4-PA2133 | Replicative pBBR1MCS-4 producing the phosphodiesterase cyclase PA2133 | [23] |
| pRK600 | Helper plasmid with conjugative properties | [24] |
| pSW196 | Site-specific integrative plasmid (attB site), araC- PBAD cassette | [25] |
| pSW196-wspR* | pSW196 containing PBAD fused to wspR* sequence subcloned from pBBR1MCS4-R246A-wspR | This study |
| pSW196-PA2133 | pSW196 containing PBAD fused to PA2133 sequence subcloned from pBBR1MCS4-PA2133 | This study |
| pTn7 CdrA-gfp(ASV)c | Suicide vector for delivery of miniTn7 harboring cdrA promoter fused to unstable GFP-encoding sequence | [26] |
| pUX-BF13 | Plasmid providing Tn7 transposase genes in trans | [26] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deruelle, V.; Berry, A.; Bouillot, S.; Job, V.; Maillard, A.P.; Elsen, S.; Huber, P. ExlA Pore-Forming Toxin: Localization at the Bacterial Membrane, Regulation of Secretion by Cyclic-Di-GMP, and Detection In Vivo. Toxins 2021, 13, 645. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090645
Deruelle V, Berry A, Bouillot S, Job V, Maillard AP, Elsen S, Huber P. ExlA Pore-Forming Toxin: Localization at the Bacterial Membrane, Regulation of Secretion by Cyclic-Di-GMP, and Detection In Vivo. Toxins. 2021; 13(9):645. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090645
Chicago/Turabian StyleDeruelle, Vincent, Alice Berry, Stéphanie Bouillot, Viviana Job, Antoine P. Maillard, Sylvie Elsen, and Philippe Huber. 2021. "ExlA Pore-Forming Toxin: Localization at the Bacterial Membrane, Regulation of Secretion by Cyclic-Di-GMP, and Detection In Vivo" Toxins 13, no. 9: 645. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090645