Neutralizing Concentrations of Anti-Botulinum Toxin Antibodies Positively Correlate with Mouse Neutralization Assay Results in a Guinea Pig Model

 , and
, and
Abstract
:1. Introduction
2. Results
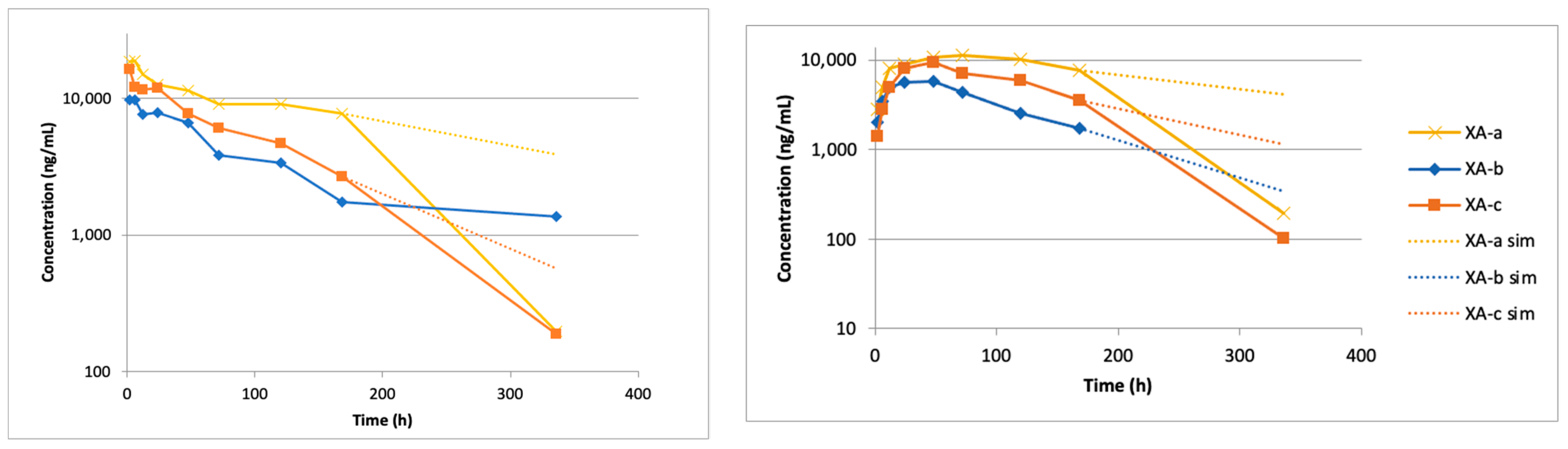
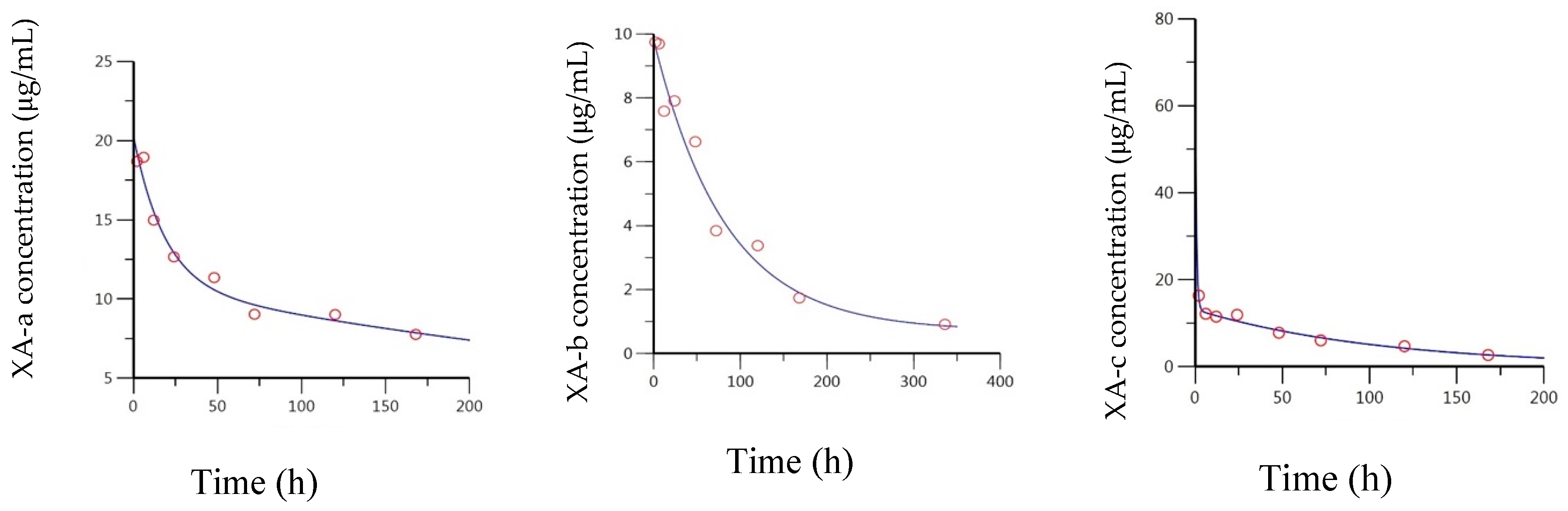
2.1. Intravenous (IV) Administration of Anti-BoNT/A Antibodies
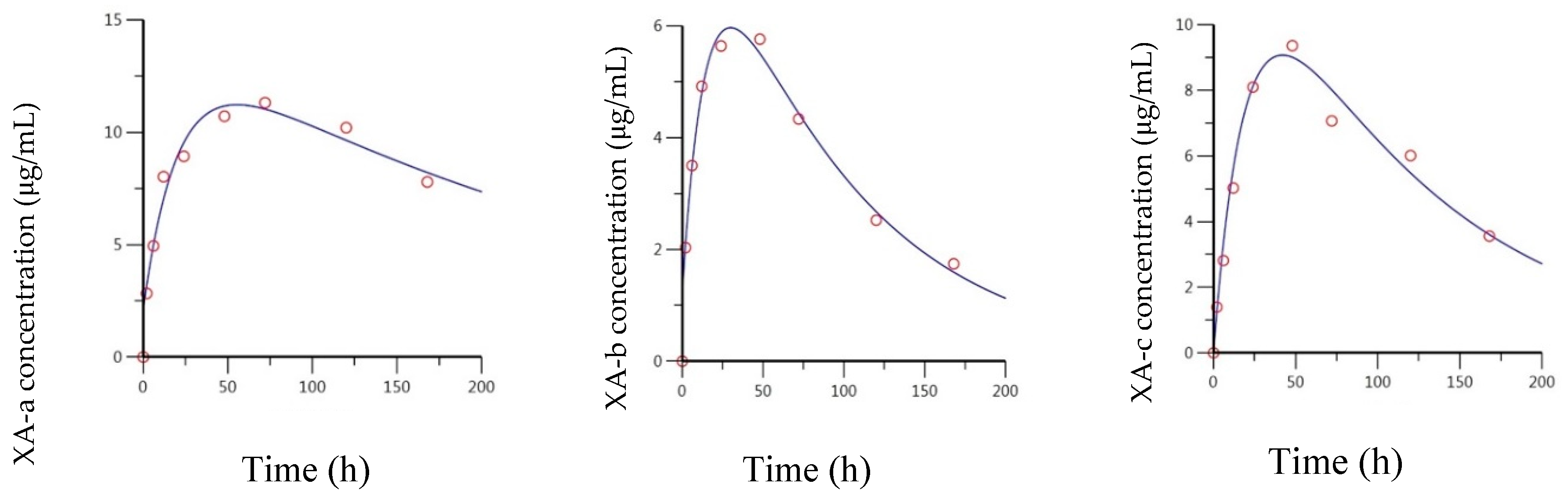
2.2. Pharmacokentics Following Intramuscular (IM) Administration of Anti-BoNT/A mAbs
2.3. Pharmacokentics Following Intravenous (IV) Administration of Anti-BoNT/B mAbs
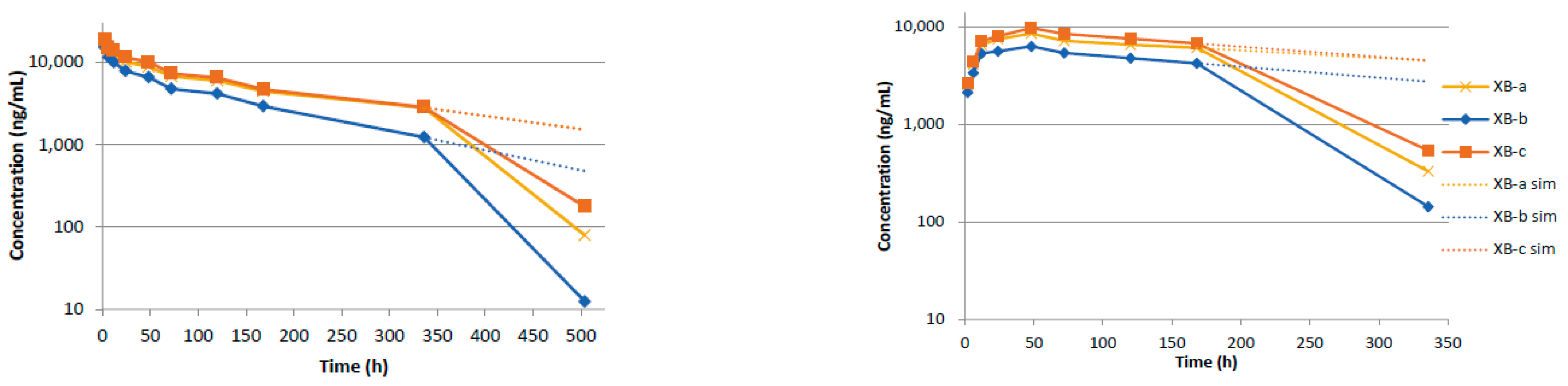
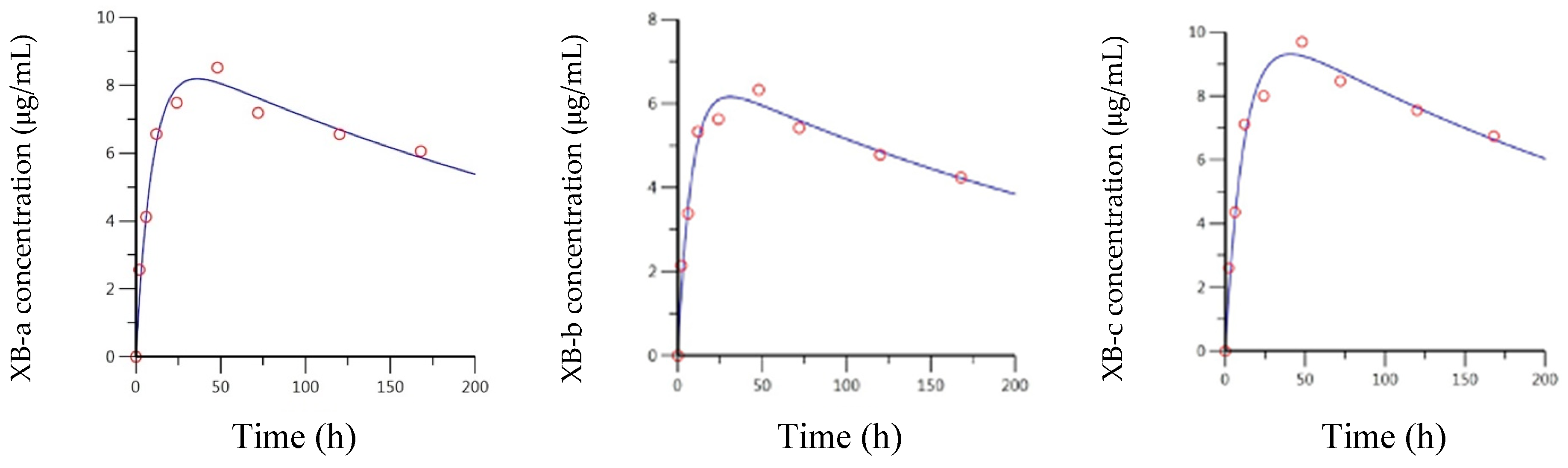
2.4. Pharmacokentics Following Intramuscular (IM) Administration of XB mAbs
2.5. Correlation of Pharmacokinetics and MNA for Anti-BoNT/A and /B mAbs
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Antibodies
5.2. Animals and Animal Welfare
5.3. Pharmacokinetics (PK)
5.4. Electrochemiluminescence (ECL) Assay for Anti-BoNT/B mAbs
5.5. ELISA Assay for Anti-BoNT/A mAbs
5.6. Measurement of Neutralizing Antibody Concentration (NAC) and Mouse Neutralization Assay (MNA)
5.7. Statistical Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ADA | Anti-drug antibody |
| AUC | Area under the curve |
| AUCinf | Area under the curve from zero timepoint extrapolated to infinity |
| AUClast | Area under the curve from zero timepoint to last measured timepoint |
| BAT® | Trade name of equine heptavalent Botulinum antitoxin |
| BIG | botulinum immune globulin |
| BoNT | Botulinum neurotoxin |
| BoNT/A | BoNT serotype A |
| BoNT/B | BoNT serotype B |
| BoNT/C | BoNT serotype C |
| BoNT/D | BoNT serotype D |
| BoNT/E | BoNT serotype E |
| BoNT/F | BoNT serotype F |
| C0 | Concentration at zero timepoint (immediately after injection) |
| Cmax | Maximal concentration |
| CI | Confidence interval |
| ECL | Electrochemiluminescence |
| ED50 | Effective dose at which 50% of the animals survived |
| ELISA | Enzyme-linked immunosorbent assay |
| F abs | Fraction absorbed (bioavailability) |
| FDA | Food and Drug Administration |
| ICCVAM | Interagency Coordinating Committee on the Validation of Alternative Methods |
| IM | Intramuscular |
| IP | Intraperitoneal |
| IV | Intravenous |
| LLOQ | Lower limit of quantitation |
| mAb | Monoclonal antibody |
| MNA | Mouse neutralization assay |
| MSD | Meso Scale Discovery |
| NAC | Neutralizing antibody concentration |
| NICEATM | National Toxicology Program Interagency Center for the Evaluation of Alternative Methods |
| NIH | National Institute of Health |
| PBS | Phosphate buffered saline |
| PK | Pharmacokinetic |
| QC | Quality control |
| t½ abs | Absorption rate half life |
| t½ elim | Elimination rate half-life |
| TMB | 3,3′,5,5′-Tetramethylbenzidine |
| XA-a | monoclonal antibody that binds BoNT/A component of G03-52-01 |
| XA-b | monoclonal antibody that binds BoNT/A component of G03-52-01 |
| XA-c | monoclonal antibody that binds BoNT/A component of G03-52-01 |
| XB-a | monoclonal antibody that binds BoNT/B component of G03-52-01 |
| XB-b | monoclonal antibody that binds BoNT/B component of G03-52-01 |
| XB-c | monoclonal antibody that binds BoNT/B component of G03-52-01 |
References
- Gupta, A.; Sumner, C.J.; Castor, M.; Maslanka, S.; Sobel, J. Adult botulism type F in the United States, 1981–2002. Neurology 2005, 65, 1694–1700. [Google Scholar] [CrossRef] [PubMed]
- Hibbs, R.G.; Weber, J.T.; Corwin, A.; Allos, B.M.; El Rehim, M.S.A.; Sharkawy, S.E.; Sarn, J.E.; McKee, K.T., Jr. Experience with the use of an investigational F(ab’)2 heptavalent botulism immune globulin of equine origin during an outbreak of type E botulism in Egypt. Clin. Infect. Dis. 1996, 23, 337–340. [Google Scholar] [CrossRef]
- Jalava, K.; Selby, K.; Pihlajasaari, A.; Kolho, E.; Dahlsten, E.; Forss, N.; Backlund, T.; Korkeala, H.; Honkanen-Buzalski, T.; Hulkko, T.; et al. Two cases of food-borne botulism in Finland caused by conserved olives, October 2011. Eurosurveillance 2011, 16, 20034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pingeon, J.M.; Vanbockstael, C.; Popoff, M.R.; King, L.A.; Deschamps, B.; Pradel, G.; Dupont, H.; Spanjaard, A.; Houdard, A.; Mazuet, C.; et al. Two outbreaks of botulism associated with consumption of green olive paste, France, September 2011. Eurosurveillance 2011, 16, 20035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, D.M. Bacterial toxins: A table of lethal amounts. Microbiol. Rev. 1982, 46, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Schechter, R.; Arnon, S.S. Extreme potency of botulinum toxin. Lancet 2000, 355, 237–238. [Google Scholar] [CrossRef]
- Lacy, D.B.; Stevens, R.C. Sequence homology and structural analysis of the clostridial neurotoxins. J. Mol. Biol. 1999, 291, 1091–1104. [Google Scholar] [CrossRef]
- Hatheway, C.L. Toxigenic clostridia. Clin. Microbiol. Rev. 1990, 3, 66–98. [Google Scholar] [CrossRef] [PubMed]
- Hatheway, C.L. Clostridium botulinum and other clostridia that produce botulinum neurotoxin. In Ecology and Control in Foods; Hauschild, A.H.W.D., Karen, L., Eds.; Marcel Dekker: New York, NY, USA, 1993; pp. 3–20. [Google Scholar]
- Barash, J.R.; Arnon, S.S. A novel strain of Clostridium botulinum that produces type B and type H botulinum toxins. J. Infect. Dis. 2014, 209, 183–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maslanka, S.E.; Lúquez, C.; Dykes, J.K.; Tepp, W.H.; Pier, C.L.; Pellett, S.; Raphael, B.H.; Kalb, S.R.; Barr, J.R.; Rao, A. A novel botulinum neurotoxin, previously reported as serotype H, has a hybrid-like structure with regions of similarity to the structures of serotypes A and F and is neutralized with serotype A antitoxin. J. Infect. Dis. 2016, 213, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Moller, V.; Scheibel, I. Preliminary report on the isolation of an apparently new type of CI. botulinum. Acta Pathol. Microbiol. Scand. 1960, 48, 80. [Google Scholar] [CrossRef]
- Arnon, S.S.; Schechter, R.; Inglesby, T.V.; Henderson, D.A.; Bartlett, J.G.; Ascher, M.S.; Eitzen, E.; Fine, A.D.; Hauer, J.; Layton, M.; et al. Botulinum toxin as a biological weapon: Medical and public health management. JAMA 2001, 285, 1059–1070. [Google Scholar] [CrossRef]
- Dembek, Z.F.; Smith, L.A.; Rusnak, J.M. Botulism: Cause, effects, diagnosis, clinical and laboratory identification, and treatment modalities. Disaster Med. Public Health Prep. 2007, 1, 122–134. [Google Scholar] [CrossRef]
- Sobel, J. Botulism. Clin. Infect. Dis. 2005, 41, 1167–1173. [Google Scholar] [CrossRef]
- Khouri, J.M.; Motter, R.N.; Arnon, S.S. Safety and immunogenicity of investigational recombinant botulinum vaccine, rBV A/B, in volunteers with pre-existing botulinum toxoid immunity. Vaccine 2018, 36, 2041–2048. [Google Scholar] [CrossRef]
- Nowakowski, A.; Wang, C.; Powers, D.B.; Amersdorfer, P.; Smith, T.J.; Montgomery, V.A.; Sheridan, R.; Blake, R.; Smith, L.A.; Marks, J.D. Potent neutralization of botulinum neurotoxin by recombinant oligoclonal antibody. Proc. Natl. Acad. Sci. USA 2002, 99, 11346–11350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Dong, J.; Lou, J.; Wen, W.; Conrad, F.; Geren, I.N.; Garcia-Rodriguez, C.; Smith, T.J.; Smith, L.A.; Ho, M. Monoclonal antibodies that inhibit the proteolytic activity of botulinum neurotoxin serotype/B. Toxins 2015, 7, 3405–3423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Rodriguez, C.; Razai, A.; Geren, I.N.; Lou, J.; Conrad, F.; Wen, W.H.; Farr-Jones, S.; Smith, T.J.; Brown, J.L.; Skerry, J.C.; et al. A Three Monoclonal Antibody Combination Potently Neutralizes Multiple Botulinum Neurotoxin Serotype E Subtypes. Toxins 2018, 10, 105. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Garcia-Rodriguez, C.; Lou, J.; Wen, W.; Conrad, F.; Zhai, W.; Smith, T.J.; Smith, L.A.; Marks, J.D. A three monoclonal antibody combination potently neutralizes multiple botulinum neurotoxin serotype F subtypes. PLoS ONE 2017, 12, e0174187. [Google Scholar] [CrossRef] [PubMed]
- Marks, J.D. Deciphering antibody properties that lead to potent botulinum neurotoxin neutralization. Mov. Disord. Off. J. Mov. Disord. Soc. 2004, 19, S101–S108. [Google Scholar] [CrossRef]
- Rasetti-Escargueil, C.; Avril, A.; Miethe, S.; Mazuet, C.; Derman, Y.; Selby, K.; Thullier, P.; Pelat, T.; Urbain, R.; Fontayne, A. The european AntibotABE framework program and its update: Development of innovative botulinum antibodies. Toxins 2017, 9, 309. [Google Scholar] [CrossRef] [Green Version]
- Casadevall, A. Passive antibody administration (immediate immunity) as a specific defense against biological weapons. Emerg. Infect. Dis. 2002, 8, 833. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Garcia-Rodriguez, C.; Manzanarez, G.; Silberg, M.A.; Conrad, F.; Bettencourt, J.; Pan, X.; Breece, T.; To, R.; Li, M.; et al. Engineered domain-based assays to identify individual antibodies in oligoclonal combinations targeting the same protein. Anal. Biochem. 2012, 430, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Q.; Li, M.; Silberg, M.A.; Conrad, F.; Bettencourt, J.; To, R.; Huang, C.; Ma, J.; Meyer, K.; Shimizu, R.; et al. Domain-based assays of individual antibody concentrations in an oligoclonal combination targeting a single protein. Anal. Biochem. 2012, 421, 351–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomic, M.T.; Espinoza, Y.; Martinez, Z.; Pham, K.; Cobb, R.R.; Snow, D.M.; Earnhart, C.G.; Pals, T.; Syar, E.E.; Niemuth, N.; et al. Monclonal antibody combinations prevent serotype A and serotype B inhalational botulism in a guinea pig model. Toxins 2019, 11, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayak, S.U.; Griffiss, J.M.; McKenzie, R.; Fuchs, E.J.; Jurao, R.A.; An, A.T.; Ahene, A.; Tomic, M.; Hendrix, C.W.; Zenilman, J.M. Safety and pharmacokinetics of XOMA 3AB, a novel mixture of three monoclonal antibodies against botulinum toxin A. Antimicrob. Agents Chemother. 2014, 58, 5047–5053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guptill, J.; Raja, S.; Juel, V.; Walter, E.; Cohen-Wolkowiez, M.; Hill, H.; Sendra, E.; Hauser, B.; Jackson, P.; Swamy, G. Safety, Tolerability and Pharmacokinetics of NTM-1632, a Novel Mixture of Three Monoclonal Antibodies against Botulinum Toxin B. Antimicrob. Agents Chemother. 2021, 65, e0232920. [Google Scholar] [CrossRef]
- Snow, D.M.; Riling, K.; Kimbler, A.; Espinoza, Y.; Wong, D.; Pham, K.; Martinez, Z.; Kraus, C.N.; Conrad, F.; Garcia-Rodriguez, C.; et al. Safety and Pharmacokinetics of a Four Monoclonal Antibody Combination Against Botulinum C and D Neurotoxins. Antimicrob. Agents Chemother. 2019, 63, e01270-19. [Google Scholar] [CrossRef] [Green Version]
- Franz, D.R.; Pitt, L.M.; Clayton, M.A.; Hanes, M.A.; Rose, K.J. Efficacy of prophylactic and therapeutic administration of antitoxin for inhalation botulism. In Botulinum and Tetanus Neurotoxins; Springer: New York, NY, USA, 1993; pp. 473–476. [Google Scholar]
- Gelzleichter, T.; Myers, M.; Menton, R.; Niemuth, N.; Matthews, M.; Langford, M. Protection against botulinum toxins provided by passive immunization with botulinum human immune globulin: Evaluation using an inhalation model. J. Appl. Toxicol. 1999, 19, S35–S38. [Google Scholar] [CrossRef]
- Interagency Coordinating Committee on the Validation of Alternative Methods. Report on the ICCVAM–NICEATM/ECVAM Scientific Workshop on Alternative Methods to Refine, Reduce, or Replace the Mouse LD50 Assay for Botulinum Toxin Testing. Available online: https://ntp.niehs.nih.gov/iccvam/docs/biologics-docs/bontwkshprept.pdf (accessed on 7 June 2021).
- Törnqvist, E.; Annas, A.; Granath, B.; Jalkesten, E.; Cotgreave, I.; Öberg, M. Strategic focus on 3R principles reveals major reductions in the use of animals in pharmaceutical toxicity testing. PLoS ONE 2014, 9, e101638. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, R.L.; Hatheway, C.; Swerdlow, D.L. Botulism in the United States: A clinical and epidemiologic review. Ann. Intern. Med. 1998, 129, 221–228. [Google Scholar] [CrossRef]
- Barker, D.; Gillum, K.T.; Niemuth, N.A.; Kodihalli, S. Therapeutic efficacy of equine botulism heptavalent antitoxin against all seven botulinum neurotoxins in symptomatic guinea pigs. PLoS ONE 2019, 14, e0222670. [Google Scholar] [CrossRef]
- Emanuel, A.; Qiu, H.; Barker, D.; Takla, T.; Gillum, K.; Neimuth, N.; Kodihalli, S. Efficacy of equine botulism antitoxin in botulism poisoning in a guinea pig model. PLoS ONE 2019, 14, e0209019. [Google Scholar] [CrossRef] [PubMed]
- Kodihalli, S.; Emanuel, A.; Takla, T.; Hua, Y.; Hobbs, C.; LeClaire, R.; O’Donnell, D.C. Therapeutic efficacy of equine botulism antitoxin in Rhesus macaques. PLoS ONE 2017, 12, e0186892. [Google Scholar] [CrossRef] [PubMed]
- Arnon, S.S.; Schechter, R.; Maslanka, S.E.; Jewell, N.P.; Hatheway, C.L. Human Botulism Immune Globulin for the Treatment of Infant Botulism. N. Engl. J. Med. 2006, 354, 462–471. [Google Scholar] [CrossRef]
- Cangene Corp. BAT® [Botulism Antitoxin Heptavalent (A, B, C, D, E, F, G)—(Equine)] Sterile Solution for Injection. Available online: https://www.fda.gov/vaccines-blood-biologics/approved-blood-products/bat-botulism-antitoxin-heptavalent-b-c-d-e-f-g-equine (accessed on 16 September 2021).
- Jankovic, J.; Brin, M.F. Therapeutic uses of botulinum toxin. N. Engl. J. Med. 1991, 324, 1186–1194. [Google Scholar] [PubMed]
- Yu, P.A.; Lin, N.H.; Mahon, B.E.; Sobel, J.; Yu, Y.; Mody, R.K.; Gu, W.; Clements, J.; Kim, H.J.; Rao, A.K. Safety and Improved Clinical Outcomes in Patients Treated With New Equine-Derived Heptavalent Botulinum Antitoxin. Clin. Infect. Dis. 2017, 66, S57–S64. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention, Botulism in the United States. In 1899–1996: Handbook for Epidemiologists, Clinicians, and Laboratory Workers; Centers for Disease Control and Prevention: Atlanta, GA, USA, 1998.
- Espinoza, Y.; Wong, D.; Ahene, A.; Der, K.; Martinez, Z.; Pham, J.; Cobb, R.R.; Farr-Jones, S.; Marks, J.; Tomic, M.T. Pharmacokinetics of human recombinant anti-botulinum toxin antibodies in rats. Toxins 2019, 11, 345. [Google Scholar] [CrossRef] [Green Version]
- Snow, D.M.; Cobb, R.R.; Martinez, J.; Finger-Baker, I.; Collins, L.; Terpening, S.; Syar, E.S.; Niemuth, N.; Kobs, D.; Barnewall, R.; et al. A monoclonal antibody combination against both serotypes A and B botulinum toxin prevents inhalational botulism in a guinea pig model. Toxins 2021, 13, 31. [Google Scholar] [CrossRef]
- USDA. Available online: https://www.nal.usda.gov/awic/institutional-animal-care-and-use-committees (accessed on 16 November 2020).
- Cardella, M.A. Botulism: Proceedings of a Symposium; U.S Department of Health, Education, Welfare, Public Service: Cincinnati, OH, USA, 1964.
- Hatheway, C.; Dang, C. Immunogenicity of the neurotoxins of Clostridium botulinum. In Therapy with Botulinum Toxin; Jankovic, J., Hallett, M., Eds.; Marcel Dekker: New York, NY, USA, 1994; Volume 25, pp. 93–107. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mAb | Dose (mg/kg), Route | Model | C0 (μg/mL) | Tmax (h) | β-Phase t1/2 (h) | t1/2 Abs (h) | t1/2 Elim (h) | AUCinf (h*μg/mL) |
|---|---|---|---|---|---|---|---|---|
| XA-a | 3, IV | 2-compartment | 20.3 | 0 | 365 | 201 | 5880 | |
| XA-b | 2-compartment | 9.83 | 0 | 5200 | 447 | 6330 | ||
| XA-c | 2-compartment | 66.9 | 0 | 73.5 | 14.8 | 1430 | ||
| XA-a | 3, IM | 1-compartment | 11.2 | 55.4 | 14.3 | 202 | 4000 | |
| XA-b | 1-compartment | 5.97 | 29.9 | 10.2 | 64.1 | 781 | ||
| XA-c | 1-compartment | 9.07 | 42.0 | 13.8 | 78.2 | 1480 |
| mAb | Dose (mg/kg), Route | Cmax (μg/mL) | Tmax (h) | t1/2 Elim (h) | AUClast (h*μg/mL) | F | Simulated Concentration at 336 h Post-dose (g/mL) |
|---|---|---|---|---|---|---|---|
| XA-a | 3, IV | 18.9 | 6 | 35.2 | 2410 | 3880 | |
| XA-b | 9.75 | 2 | 93.9 | 1020 | - | ||
| XA-c | 16.4 | 2 | 34.4 | 1390 | 569 | ||
| XA-a | 3, IM | 11.3 | 72 | 24.5 | 2260 | 94% | 4170 |
| XA-b | 5.76 | 48 | 73 | 627 | 83% | 344 | |
| XA-c | 9.36 | 48 | 30.4 | 1370 | 99% | 1140 |
| Administration Route | mAb | Dose (mg/kg) | Model | C0 (μg/mL) | Tmax (h) | β-Phase t1/2 (h) | t1/2 Abs (h) | t1/2 Elim (h) | AUCinf (h*μg/mL) |
|---|---|---|---|---|---|---|---|---|---|
| IV | XB-a | 3 | 2-compartment | 20.7 | 0 | 144 | 77.1 | 2300 | |
| XB-b | 2-compartment | 17.2 | 0 | 115 | 59.4 | 1470 | |||
| XB-c | 2-compartment | 20.7 | 0 | 133 | 81.1 | 2420 | |||
| IM | XB-a | 3 | 1-compartment | 8.19 | 36.1 | 6.71 | 254 | 3310 | |
| XB-b | 1-compartment | 6.16 | 31.0 | 5.59 | 237 | 2300 | |||
| XB-c | 1-compartment | 9.3 | 40.7 | 8.08 | 236 | 3570 |
| Administration Route | mAb | Dose (mg/kg) | Cmax (μg/mL) | Tmax (h) | t1/2 Elim (h) | AUClast (h*μg/mL) | F (%) | Simulated C336 (μg/mL) | Simulated C504 (μg/mL) |
|---|---|---|---|---|---|---|---|---|---|
| IV | XB-a | 3 | 18.2 | 2 | 69.5 | 2060 | 1040 | ||
| XB-b | 15.1 | 2 | 49.6 | 1380 | 361 | ||||
| XB-c | 18.8 | 2 | 77.2 | 2220 | 1000 | ||||
| IM | XB-a | 3 | 8.52 | 48 | 38.1 | 1670 | 81 | 4480 | |
| XB-b | 6.33 | 48 | 26.5 | 1210 | 88 | 2760 | |||
| XB-c | 9.69 | 48 | 46.3 | 1890 | 85 | 4520 |
| XA-a | XA-b | XA-c | All 3 Anti-A mAbs | XB-a | XB-b | XB-c | All 3 Anti-B mAbs | |
|---|---|---|---|---|---|---|---|---|
| Best fit Slope (95% CI) | 815.7 (614.0–1017) | 453.8 (335.9–571.8) | 618.9 461.4–776.5) | 1888 (1428–2349) | 324.8 (302.8–346.9) | 253.6 (234–273.1) | 358.3 (335.3–381.3) | 880.3 (694.8–1066) |
| Goodness of fit | 3855 | 2289 | 3011 | 8064 | 2169 | 1922 | 2261 | 6605 |
| R2 | 0.6934 | 0.5765 | 0.6795 | 0.6949 | 0.8012 | 0.7659 | 0.8210 | 0.7436 |
| p value | 0.0028 | 0.0176 | 0.0036 | 0.0027 | 0.0005 | 0.0009 | 0.0003 | 0.0013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomic, M.T.; Farr-Jones, S.; Syar, E.S.; Niemuth, N.; Kobs, D.; Hackett, M.J.; Espinoza, Y.; Martinez, Z.; Pham, K.; Snow, D.M.; et al. Neutralizing Concentrations of Anti-Botulinum Toxin Antibodies Positively Correlate with Mouse Neutralization Assay Results in a Guinea Pig Model. Toxins 2021, 13, 671. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090671
Tomic MT, Farr-Jones S, Syar ES, Niemuth N, Kobs D, Hackett MJ, Espinoza Y, Martinez Z, Pham K, Snow DM, et al. Neutralizing Concentrations of Anti-Botulinum Toxin Antibodies Positively Correlate with Mouse Neutralization Assay Results in a Guinea Pig Model. Toxins. 2021; 13(9):671. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090671
Chicago/Turabian StyleTomic, Milan T., Shauna Farr-Jones, Emily S. Syar, Nancy Niemuth, Dean Kobs, Michael J. Hackett, Yero Espinoza, Zacchary Martinez, Khanh Pham, Doris M. Snow, and et al. 2021. "Neutralizing Concentrations of Anti-Botulinum Toxin Antibodies Positively Correlate with Mouse Neutralization Assay Results in a Guinea Pig Model" Toxins 13, no. 9: 671. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13090671