Tetrodotoxin/Saxitoxins Selectivity of the Euryhaline Freshwater Pufferfish Dichotomyctere fluviatilis

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
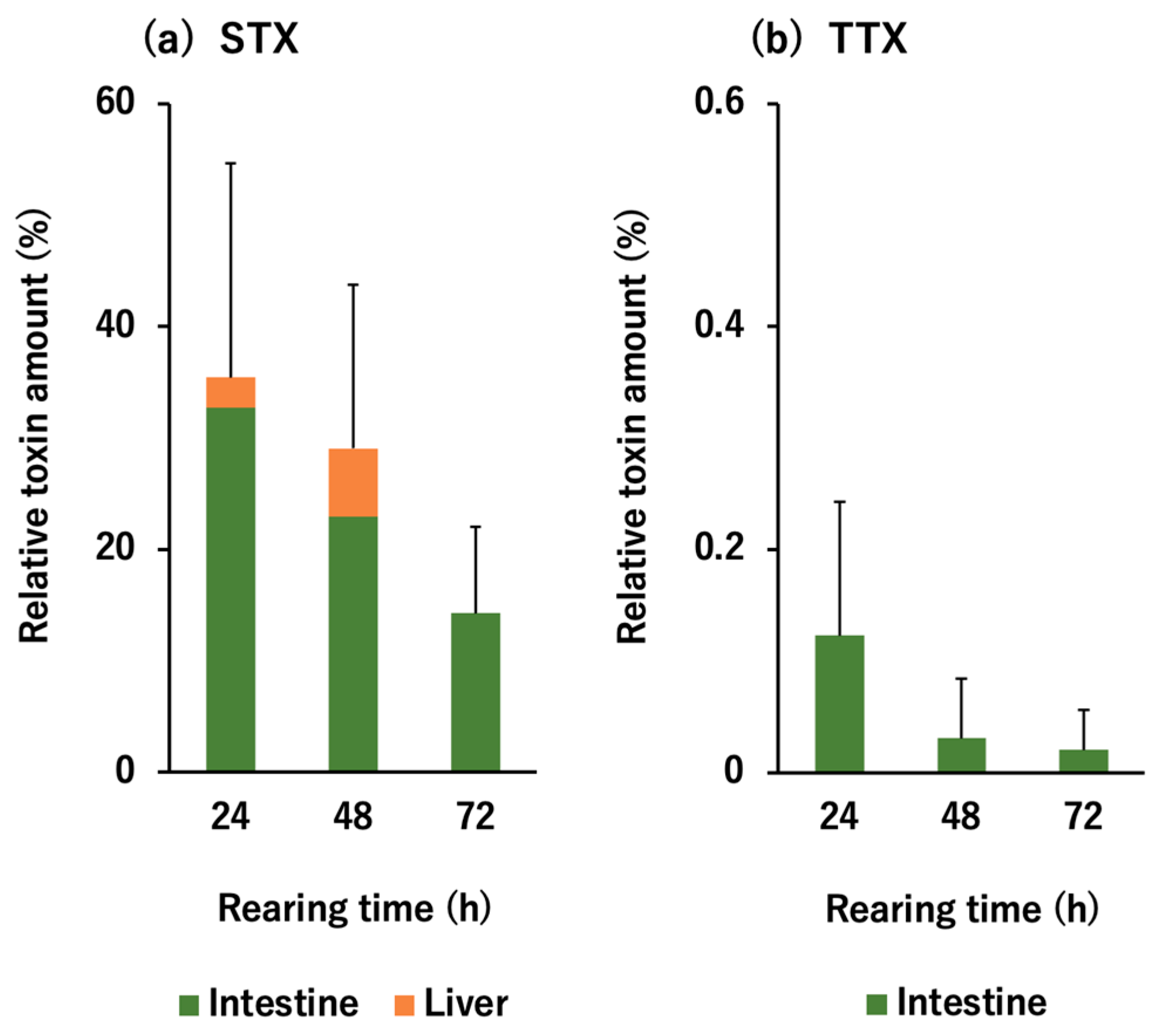
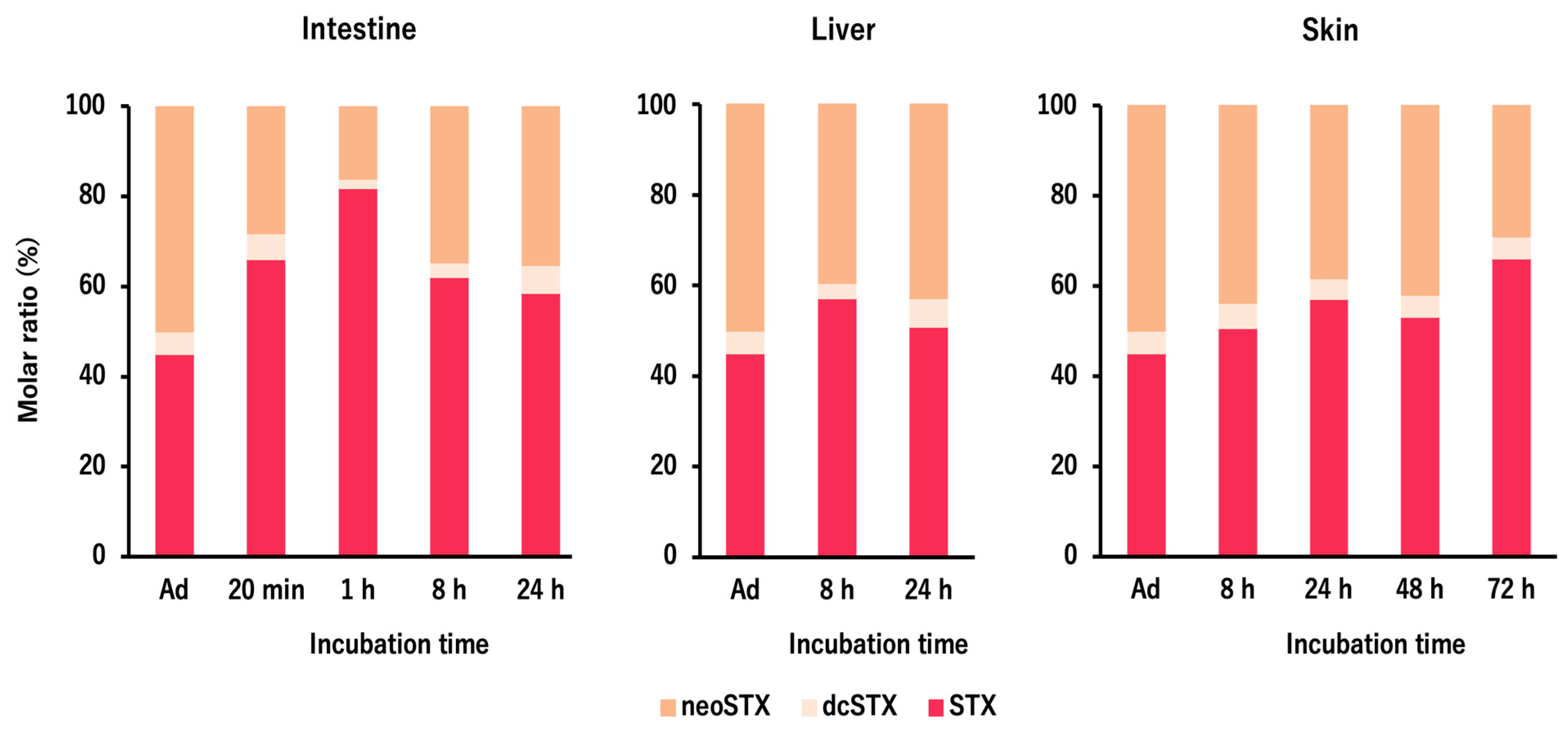
2.1. In Vivo Toxin Administration Experiment with D. fluviatilis
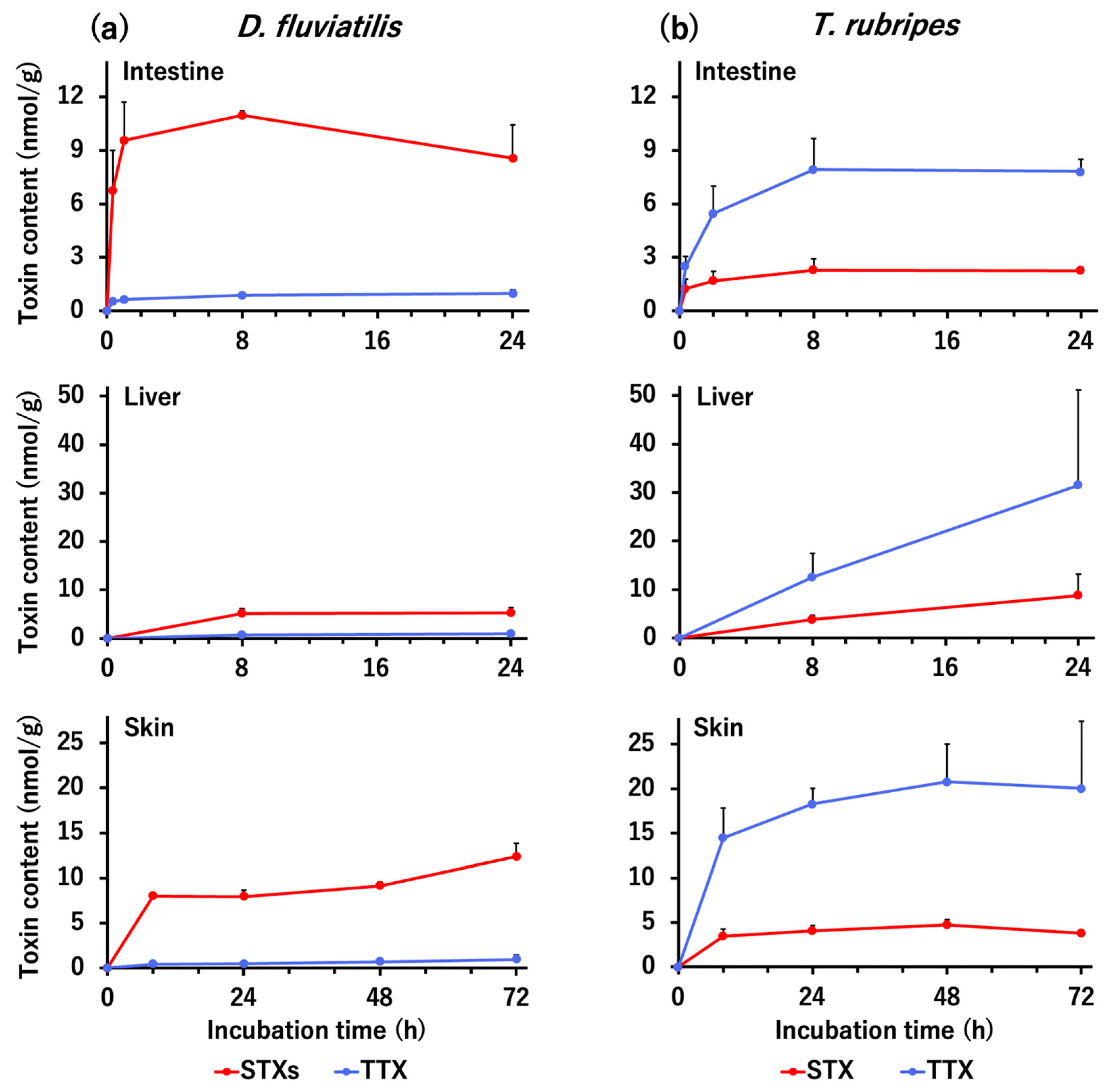
2.2. In Vitro Tissue Slice Incubation Experiment with D. fluviatilis and T. rubripes
3. Discussion
3.1. Selective Toxin Accumulation Ability of D. fluviatilis
3.2. Selective Toxin Uptake Ability of The Tissues in D. fluviatilis and T. rubripes
3.3. Toxin Profile and Ecology of D. fluviatilis in Nature
3.4. Future Perspective
4. Conclusions
5. Materials and Methods
5.1. Pufferfish Specimens
5.2. Toxin Preparation
5.3. In Vivo Toxin Administration Experiment
5.4. In Vitro Tissue Slice Incubation Experiment
5.5. Toxin Quantification
5.6. Data Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Noguchi, T.; Arakawa, O.; Takatani, T. TTX accumulation in pufferfish. Comp. Biochem. Physiol. Part D 2006, 1, 145–152. [Google Scholar] [CrossRef]
- Noguchi, T.; Arakawa, O. Tetrodotoxin—distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakawa, O.; Takatani, T.; Taniyama, S.; Tatsuno, R. Toxins of pufferfish—distribution, accumulation mechanism, and physiologic functions. Aqua BioScience Monogr. 2017, 10, 41–80. [Google Scholar] [CrossRef] [Green Version]
- Tani, T. Nihonsan Fugu no Chudokugakuteki Kenkyu (Toxicological Studies on Japanese Puffer); Teikoku-tosho: Tokyo, Japan, 1945. [Google Scholar]
- Noguchi, T.; Ebesu, J.S.M. Puffer poisoning: Epidemiology and treatment. J. Toxicol. Toxin Rev. 2001, 20, 1–10. [Google Scholar] [CrossRef]
- Sato, S.; Kodama, M.; Ogata, T.; Saitanu, K.; Furuya, M.; Hirayama, K.; Kamimura, K. Saxitoxin as a toxic principle of a freshwater puffer, Tetraodon fangi, in Thailand. Toxicon 1997, 35, 137–140. [Google Scholar] [CrossRef]
- Zaman, L.; Arakawa, O.; Shimosu, A.; Onoue, Y. Occurrence of paralytic shellfish poison in Bangladeshi freshwater puffers. Toxicon 1997, 35, 423–431. [Google Scholar] [CrossRef]
- Kungsuwan, A.; Arakawa, O.; Promdet, M.; Onoue, Y. Occurrence of paralytic shellfish poisons in Thai freshwater puffers. Toxicon 1997, 35, 1341–1346. [Google Scholar] [CrossRef]
- Ngy, L.; Tada, K.; Yu, C.F.; Takatani, T.; Arakawa, O. Occurrence of paralytic shellfish toxins in Cambodian Mekong pufferfish Tetraodon turgidus: Selective toxin accumulation in the skin. Toxicon 2008, 51, 280–288. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Yamada, A.; Goto, Y.; Horn, L.; Ngy, L.; Wada, M.; Doi, H.; Lee, J.S.; Takatani, T.; Arakawa, O. Phylogeny and toxin profile of freshwater pufferfish (genus Pao) collected from 2 different regions in Cambodia. Toxins 2020, 12, 689. [Google Scholar] [CrossRef] [PubMed]
- Saitanu, K.; Laobhripatr, S.; Limpakarnjanarat, K.; Sangwanloy, O.; Sudhasaneya, S.; Anuchatvorakul, B.; Leelasitorn, S. Toxicity of the freshwater puffer fish Tetraodon fungi and T. palembangensis from Thailand. Toxicon 1991, 29, 895–897. [Google Scholar] [CrossRef]
- Cusick, K.D.; Sayler, G.S. An overview on the marine neurotoxin, saxitoxin: Genetics, molecular targets, methods of detection and ecological functions. Mar. Drugs 2013, 11, 991–1018. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Ogata, T.; Borja, V.; Gonzales, C.; Fukuyo, Y.; Kodama, M. Frequent occurrence of paralytic shellfish poisoning toxins as dominant toxins in marine puffer from tropical water. Toxicon 2000, 38, 1101–1109. [Google Scholar] [CrossRef]
- Nakashima, K.; Arakawa, O.; Taniyama, S.; Nonaka, M.; Takatani, T.; Yamamori, K.; Fuchi, Y.; Noguchi, T. Occurrence of saxitoxins as a major toxin in the ovary of a marine puffer Arothron firmamentum. Toxicon 2004, 43, 207–212. [Google Scholar] [CrossRef]
- Landsberg, J.H.; Hall, S.; Johannessen, J.N.; White, K.D.; Conrad, S.M. Saxitoxin puffer fish poisoning in the United States, with the first report of Pyrodinium bahamense as the putative toxin source. Environ. Health Perspect. 2006, 114, 1502–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrientos, R.G.; Hernández-Mora, G.; Alegre, F.; Field, T.; Flewelling, L.; McGrath, S.; Deeds, J.; Chacón, Y.S.; Arrieta, K.R.; Vargas, E.C.; et al. Saxitoxin poisoning in green turtles (Chelonia mydas) linked to scavenging on mass mortality of Caribbean sharpnose puffer fish (Canthigaster rostrata-Tetraodontidae). Front. Vet. Sci. 2019, 6, 466. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Sonoyama, T.; Yamada, M.; Gao, W.; Tatsuno, R.; Takatani, T.; Arakawa, O. Co-occurrence of tetrodotoxin and saxitoxins and their intra-body distribution in the pufferfish Canthigaster valentini. Toxins 2020, 12, 436. [Google Scholar] [CrossRef] [PubMed]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Persson, K.M. Quick detection method for paralytic shellfish toxins (PSTs) monitoring in freshwater—A review. Chemosphere 2021, 265, 128591. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Arakawa, O.; Takatani, T. Toxicity of pufferfish Takifugu rubripes cultured in netcages at sea or aquaria on land. Comp. Biochem. Physiol. Part D 2006, 1, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Murakami, Y.; Emoto, Y.; Ngy, L.; Taniyama, S.; Yagi, M.; Takatani, T.; Arakawa, O. Transfer profile of intramuscularly administered tetrodotoxin to non-toxic cultured specimens of the pufferfish Takifugu rubripes. Toxicon 2009, 53, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Araki, T.; Tatsuno, R.; Nina, S.; Ikeda, K.; Hamasaki, M.; Sakakura, Y.; Takatani, T.; Arakawa, O. Transfer profile of intramuscularly administered tetrodotoxin to artificial hybrid specimens of pufferfish, Takifugu rubripes and Takifugu niphobles. Toxicon 2011, 58, 565–569. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Araki, T.; Tatsuno, R.; Nina, S.; Ikeda, K.; Takatani, T.; Arakawa, O. Transfer profile of orally and intramuscularly administered tetrodotoxin to artificial hybrid specimens of the pufferfish Takifugu rubripes and Takifugu porphyreus. J. Food Hyg. Soc. Jpn. 2012, 55, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Tatsuno, R.; Shikina, M.; Shirai, Y.; Wang, J.; Soyano, K.; Nishihara, G.N.; Takatani, T.; Arakawa, O. Change in the transfer profile of orally administered tetrodotoxin to non-toxic cultured pufferfish Takifugu rubripes depending of its development stage. Toxicon 2013, 65, 76–80. [Google Scholar] [CrossRef]
- Tatsuno, R.; Gao, W.; Ibi, K.; Mine, T.; Okita, K.; Nishihara, G.N.; Takatani, T.; Arakawa, O. Profile differences in tetrodotoxin transfer to skin and liver in the pufferfish Takifugu rubripes. Toxicon 2017, 130, 73–78. [Google Scholar] [CrossRef]
- Gao, W.; Kanahara, Y.; Yamada, M.; Tatsuno, R.; Yoshikawa, H.; Doi, H.; Takatani, T.; Arakawa, O. Contrasting toxin selectivity between the marine pufferfish Takifugu pardalis and the freshwater pufferfish Pao suvattii. Toxins 2019, 11, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagashima, Y.; Toyoda, M.; Hasobe, M.; Shimakura, K.; Shiomi, K. In vitro accumulation of tetrodotoxin in pufferfish liver tissue slices. Toxicon 2003, 41, 569–574. [Google Scholar] [CrossRef]
- Nagashima, Y.; Ohta, A.; Yin, X.; Ishizaki, S.; Matsumoto, T.; Doi, H.; Ishibashi, T. Difference in uptake of tetrodotoxin and saxitoxins into liver tissue slices among pufferfish, boxfish and porcupinefish. Mar. Drugs 2018, 16, 17. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, T.; Nagashima, Y.; Takayama, K.; Shimakura, K.; Shiomi, K. Difference between tetrodotoxin and saxitoxins in accumulation in puffer fish Takifugu rubripes liver tissue slices. Fish Physiol. Biochem. 2005, 31, 95–100. [Google Scholar] [CrossRef]
- Matsumoto, T.; Nagashima, Y.; Kusuhara, H.; Sugiyama, Y.; Ishizaki, S.; Shimakura, K.; Shiomi, K. Involvement of carrier-mediated transport system in uptake of tetrodotoxin into liver tissue slices of puffer fish Takifugu rubripes. Toxicon 2007, 50, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Yamada, M.; Ohki, R.; Nagashima, Y.; Tatsuno, R.; Ikeda, K.; Kawatsu, K.; Takatani, T.; Arakawa, O. Evaluation of the tetrodotoxin uptake ability of pufferfish Takifugu rubripes tissues according to age using an in vitro tissue slice incubation method. Toxicon 2020, 174, 8–12. [Google Scholar] [CrossRef]
- Kottelat, M. The fishes of the inland waters of Southeast Asia: A catalogue and core bibiography of the fishes known to occur in freshwaters, mangroves and estuaries. Raffles Bull. Zool. 2013, 27, 469–480. [Google Scholar]
- Rainboth, W.J. Fishes of the Cambodian Mekong; Food and Agriculture Organization of The United Nations: Rome, Italy, 1996; p. 226. [Google Scholar]
- Pethiyagoda, R. Freshwater fishes of Sri Lanka; The Wildlife Heritage Trust of Sri Lanka: Colombo, Sri Lanka, 1991; pp. 1–362. [Google Scholar]
- Riede, K. Global Register of Migratory Species: From Global to Regional Scales: Final Report of the R&D-Projekt 808 05 081; Federal Agency for Nature Conservation: Bonn, Germany, 2004; pp. 1–330. [Google Scholar]
- Talwar, P.K.; Jhingran, A.G. Inland Fishes of India and Adjacent Countries; A.A. Balkema: Rotterdam, The Netherlands, 1992; Volume 2, pp. 1055–1056. [Google Scholar]
- Schaefer, F. Brackish-water fishes: All About Species, Care and Breeding; Hollywood Import & Export, Inc.: Gainesville, FL, USA, 2005; pp. 1–80. [Google Scholar]
- Rakshit, A.; Chanda, I. On some fish diversity pattern Sundarban delta, West Bengal. J. Environ. Sociobiol. 2018, 15, 75–82. [Google Scholar]
- Shamsuzzaman, M.M.; Rashid, A.H.A.; Mamun, M.A.A.; Mazumder, S.K.; Haque, M.A. Present status of marine puffer fishes in Bangladesh. J. Aquac. Res. Dev. 2015, 6, 370. [Google Scholar] [CrossRef] [Green Version]
- Crnogorac-Jurcevic, T.; Brown, J.R.; Lehrach, H.; Schalkwyk, L.C. Tetraodon fluviatilis, a new puffer fish model for genome studies. Genomics 1997, 41, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, Y.; Doi, H.; Yamanoue, Y.; Kinoshita, S.; Ishibashi, T.; Ushio, H.; Asakawa, S.; Nishida, M.; Watabe, S. Molecular phylogenetic relationship of Tetraodon pufferfish based on mitochondrial DNA analysis. Fish. Sci. 2013, 79, 243–250. [Google Scholar] [CrossRef]
- Mahmud, Y.; Yamamori, K.; Noguchi, T. Occurrence of TTX in a brackish water puffer ‘midorifugu’, Tetraodon nigroviridis, collected from Thailand. J. Food Hyg. Soc. Jpn. 1999, 40, 363–367. [Google Scholar] [CrossRef] [Green Version]
- Mahmud, Y.; Yamamori, K.; Noguchi, T. Toxicity and tetrodotoxin as the toxic principle of a brackish water puffer, Tetraodon steindachneri, collected from Thailand. J. Food Hyg. Soc. Jpn. 1999, 40, 391–395. [Google Scholar] [CrossRef]
- Chulanetra, M.; Sookrung, N.; Srimanote, P.; Indrawattana, N.; Thanongsaksrikul, J.; Sakolvaree, Y.; Chongsa-Nguan, M.; Kurazono, H.; Chaicumpa, W. Toxic marine puffer fish in Thailand seas and tetrodotoxin they contained. Toxins 2011, 3, 1249–1262. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, K.; Emoto, Y.; Tatsuno, R.; Wang, J.J.; Ngy, L.; Taniyama, S.; Takatani, T.; Arakawa, O. Maturation-associated changes in toxicity of the pufferfish Takifugu poecilonotus. Toxicon 2010, 55, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Kanahara, Y.; Tatsuno, R.; Soyano, K.; Nishihara, G.N.; Urata, C.; Takatani, T.; Arakawa, O. Maturation-associated changes in internal distribution and intra-ovarian microdistribution of tetrodotoxin in the pufferfish Takifugu pardalis. Fish. Sci. 2018, 84, 723–732. [Google Scholar] [CrossRef]
- Shimizu, Y.; Yoshioka, M. Transformation of paralytic shellfish toxins as demonstrated in scallop homogenates. Science 1981, 212, 547–549. [Google Scholar] [CrossRef]
- Oshima, Y. Chemical and enzymatic transformation of paralytic shellfish toxins in marine organisms. In Harmful Marine Algal Blooms; Lassus, P., Arzul, G., Erard, E., Gentien, P., Marcaillou, C., Eds.; Lavoisier: Paris, France, 1995; pp. 475–480. [Google Scholar]
- Arakawa, O.; Noguchi, T.; Onoue, Y. Transformation of gonyautoxins in the xanthid crab Atergatis floridus. Fish. Sci. 1998, 64, 334–337. [Google Scholar] [CrossRef]
- Untario, N.; Dewi, T.C.; Widodo, M.A.; Rahaju, P. Effect of tetrodotoxin from crude puffer fish (Tetraodon fluviatilis) liver extract on intracellular calcium level and apoptosis of HeLa cell culture. J. Trop. Life. Sci. 2017, 7, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Yotsu-Yamashita, M.; Sugimoto, A.; Terakawa, T.; Shoji, Y.; Miyazawa, T.; Yasumoto, T. Purification, characterization, and cDNA cloning of a novel soluble saxitoxin and tetrodotoxin binding protein from plasma of the puffer fish, Fugu pardalis. Eur. J. Biochem. 2001, 268, 5937–5946. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Okoshi, N.; Watanabe, K.; Araki, N.; Yamaki, H.; Shoji, Y.; Terakawa, T. Localization of pufferfish saxitoxin and tetrodotoxin binding protein (PSPBP) in the tissues of the pufferfish, Takifugu pardalis, analyzed by immunohistochemical staining. Toxicon 2013, 72, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Hashiguchi, Y.; Lee, J.M.; Shiraishi, M.; Komatsu, S.; Miki, S.; Shimasaki, Y.; Mochioka, N.; Kusakabe, T.; Oshima, Y. Characterization and evolutionary analysis of tributyltin-binding protein and pufferfish saxitoxin and tetrodotoxin-binding protein genes in toxic and nontoxic pufferfishes. J. Evol. Biol. 2015, 28, 1103–1118. [Google Scholar] [CrossRef] [Green Version]
- Arakawa, O.; Noguchi, T.; Shida, Y.; Onoue, Y. Occurrence of 11-oxotetrodotoxin and 11-nortetrodotoxin-6(R)-ol in a xanthid crab Atergatis floridus collected at Kojima, Ishigaki Island. Fish. Sci. 1994, 60, 769–771. [Google Scholar] [CrossRef]
- Arakawa, O.; Noguchi, T.; Shida, Y.; Onoue, Y. Occurrence of carbamoyl-N-hydroxy derivatives of saxitoxin and neosaxitoxin in a xanthid crab Zosimus aeneus. Toxicon 1994, 32, 175–183. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, H.; Sakai, T.; Nagashima, Y.; Doi, H.; Takatani, T.; Arakawa, O. Tetrodotoxin/Saxitoxins Selectivity of the Euryhaline Freshwater Pufferfish Dichotomyctere fluviatilis. Toxins 2021, 13, 731. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100731
Zhu H, Sakai T, Nagashima Y, Doi H, Takatani T, Arakawa O. Tetrodotoxin/Saxitoxins Selectivity of the Euryhaline Freshwater Pufferfish Dichotomyctere fluviatilis. Toxins. 2021; 13(10):731. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100731
Chicago/Turabian StyleZhu, Hongchen, Towa Sakai, Yuji Nagashima, Hiroyuki Doi, Tomohiro Takatani, and Osamu Arakawa. 2021. "Tetrodotoxin/Saxitoxins Selectivity of the Euryhaline Freshwater Pufferfish Dichotomyctere fluviatilis" Toxins 13, no. 10: 731. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins13100731