Variation of Microbial Community and Fermentation Quality in Corn Silage Treated with Lactic Acid Bacteria and Artemisia argyi during Aerobic Exposure


Abstract
:1. Introduction
2. Results
2.1. Characteristics of Whole Plant Corn and Artemisia Argyi before Ensiling
2.2. Fermentation Quality, Chemical Composition and Microbial Population of Silage
2.2.1. Fermentation Quality
2.2.2. Chemical Composition
2.2.3. Microbial Population
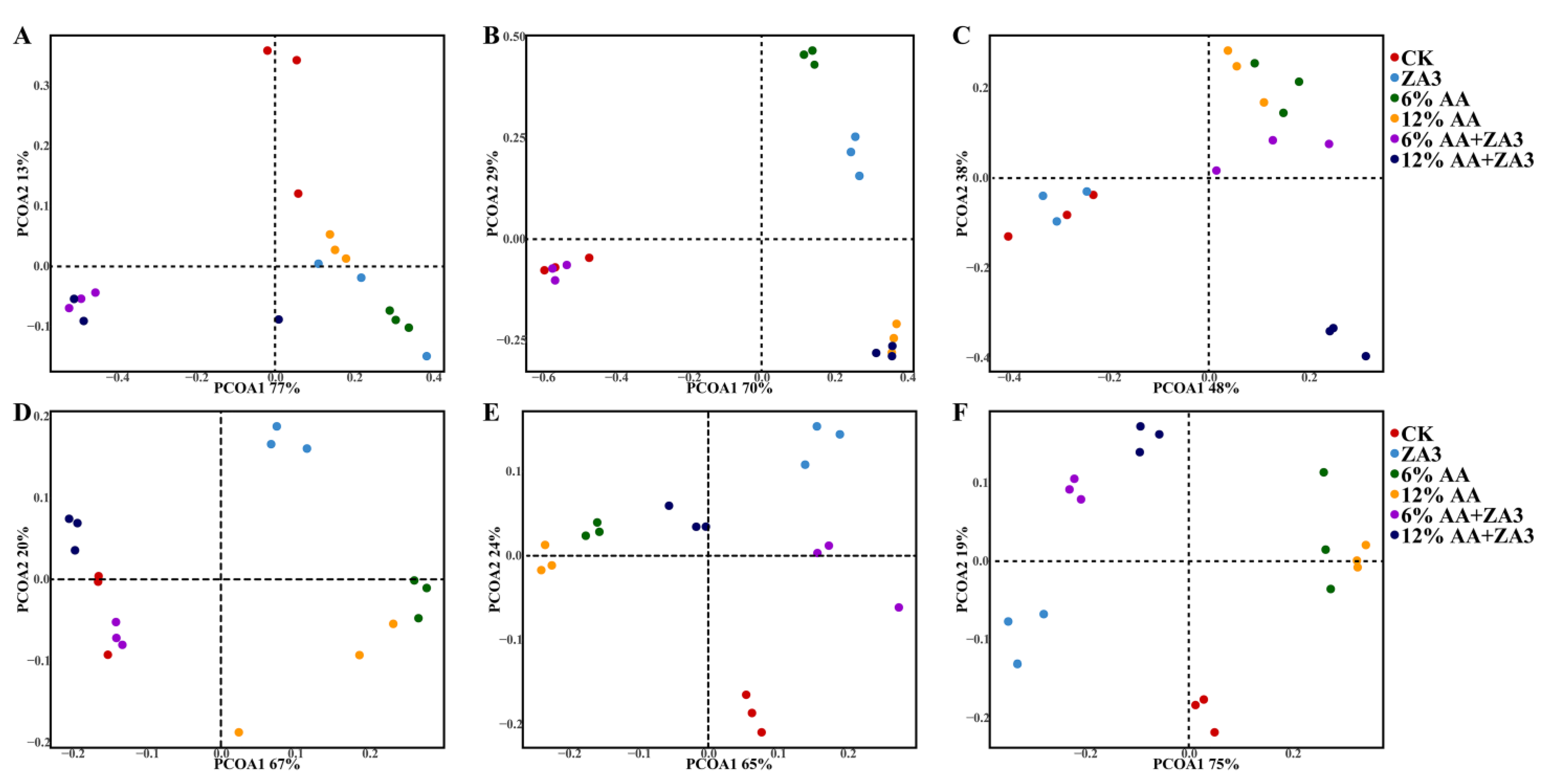
2.3. Diversity of Bacteria and Fungi
2.4. Relative Abundances of Bacterial and Fungi
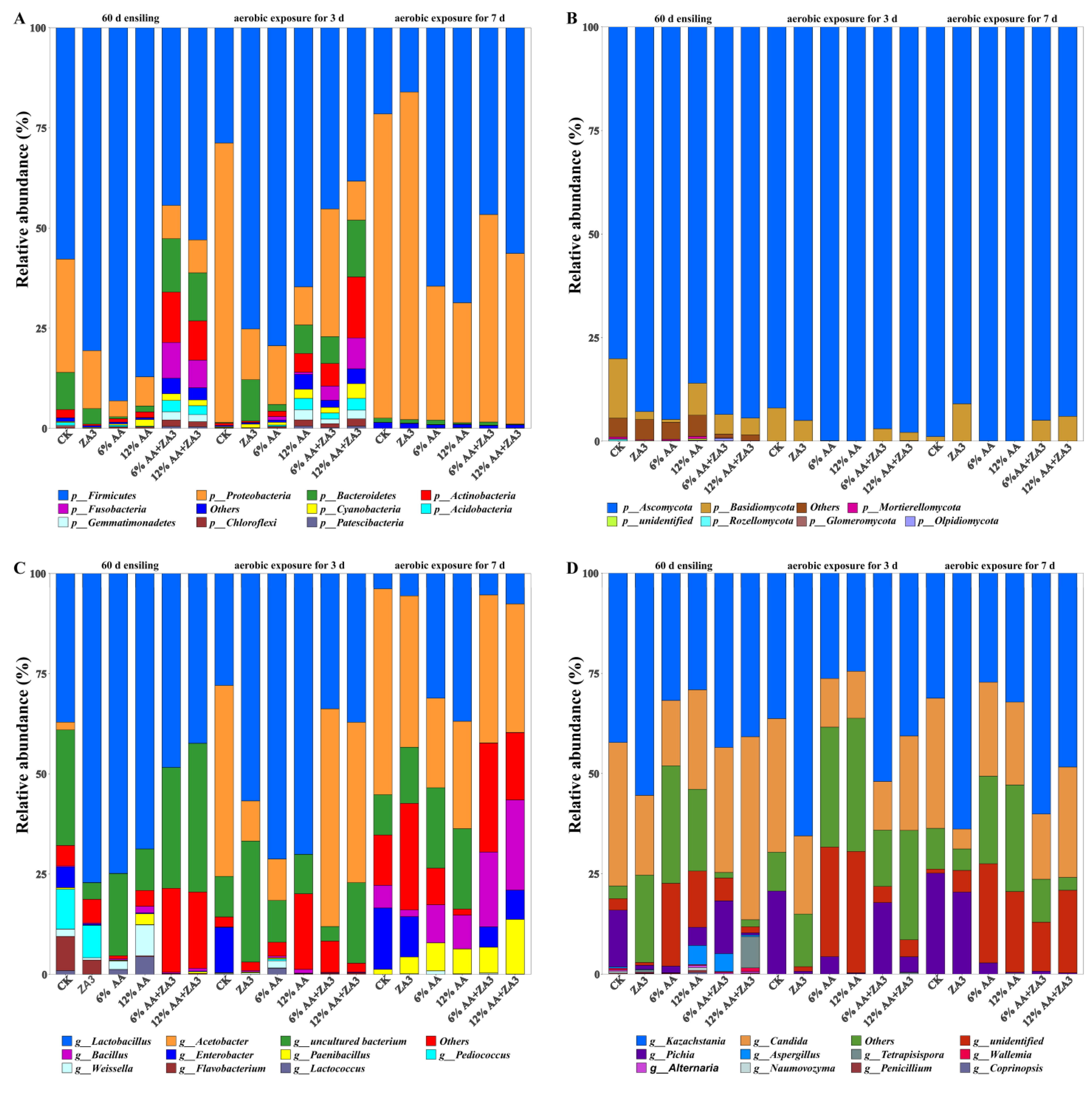
2.4.1. Phylum Level
2.4.2. Genus Level
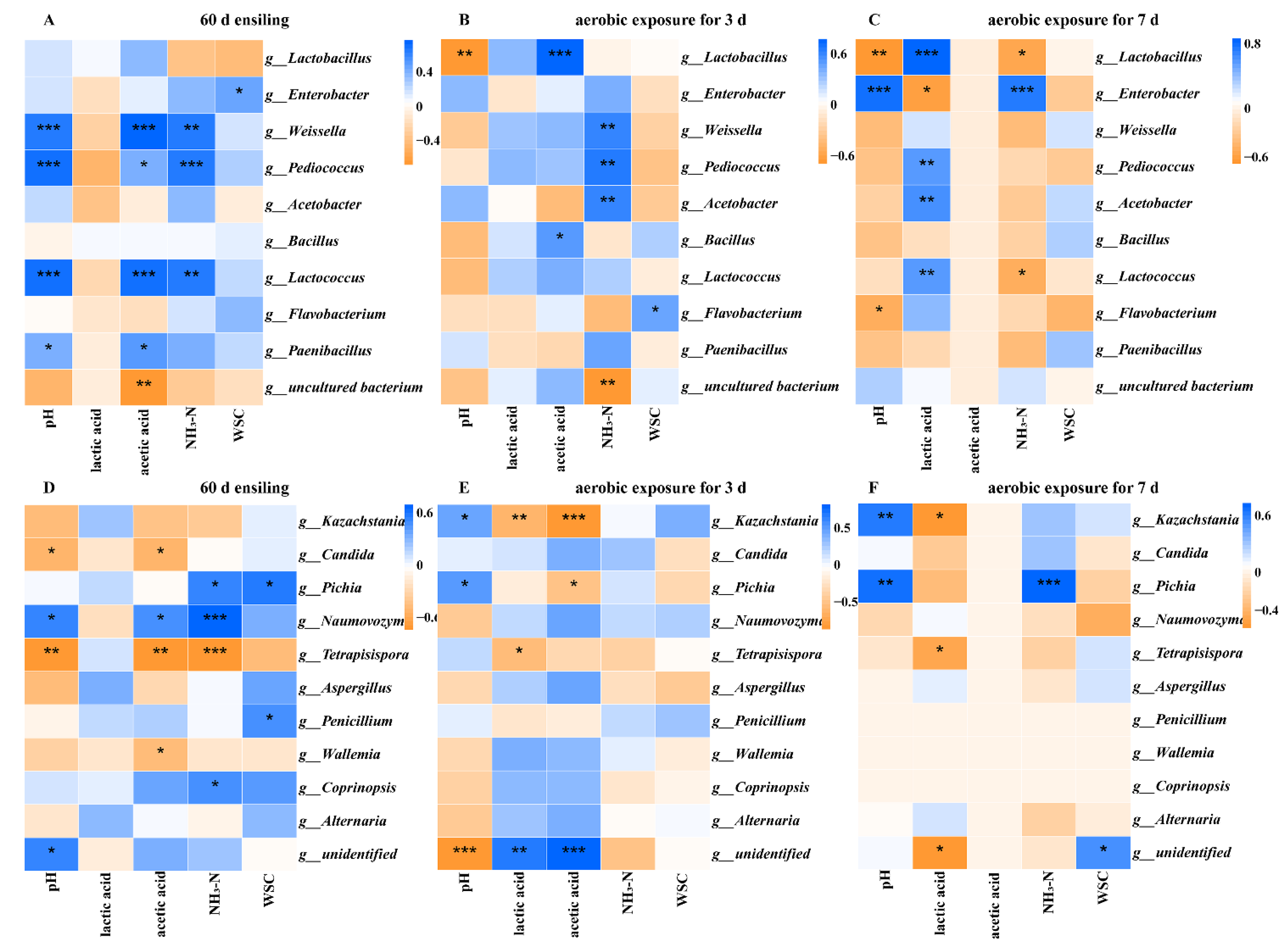
2.5. Correlation Analyses of the Bacterial and Fungi Community with Fermentation Properties
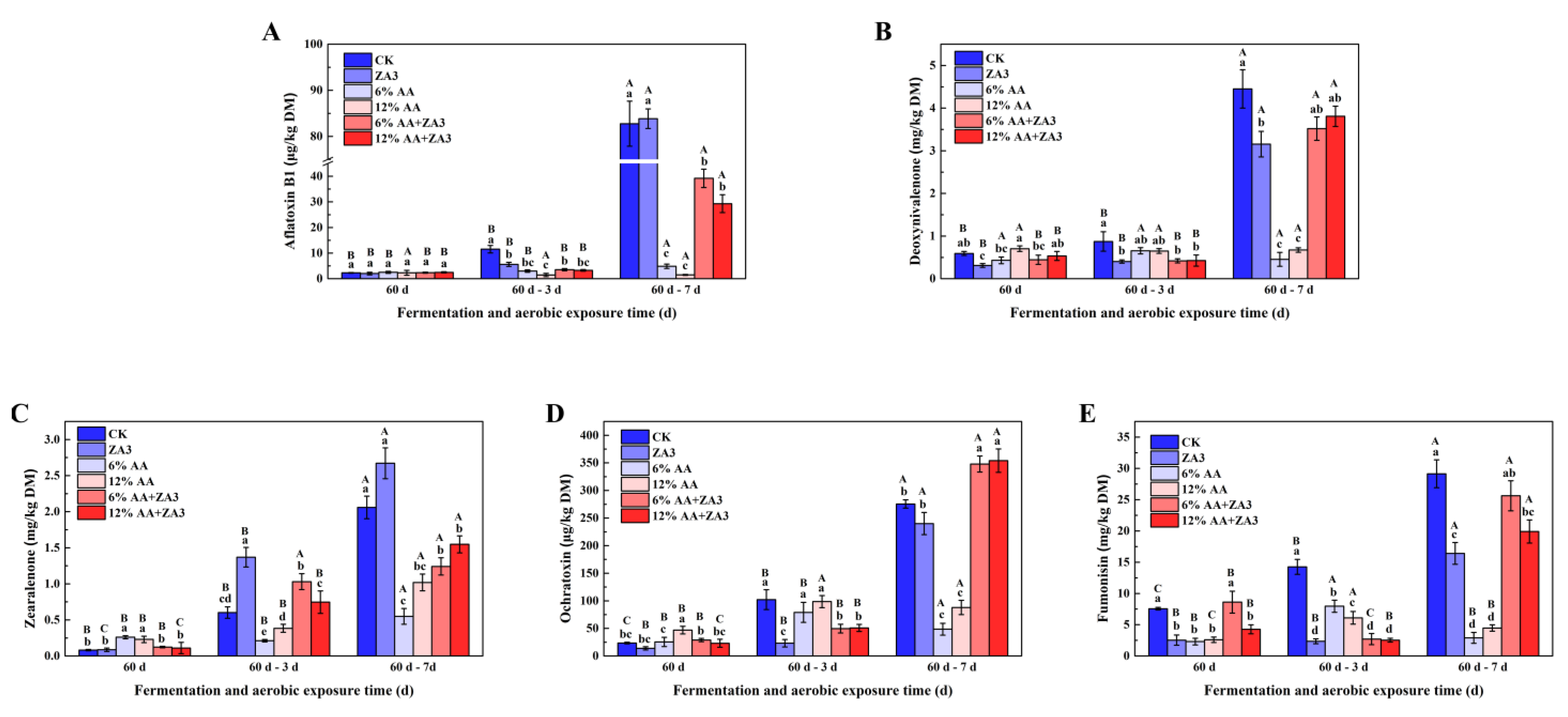
2.6. Toxin Content
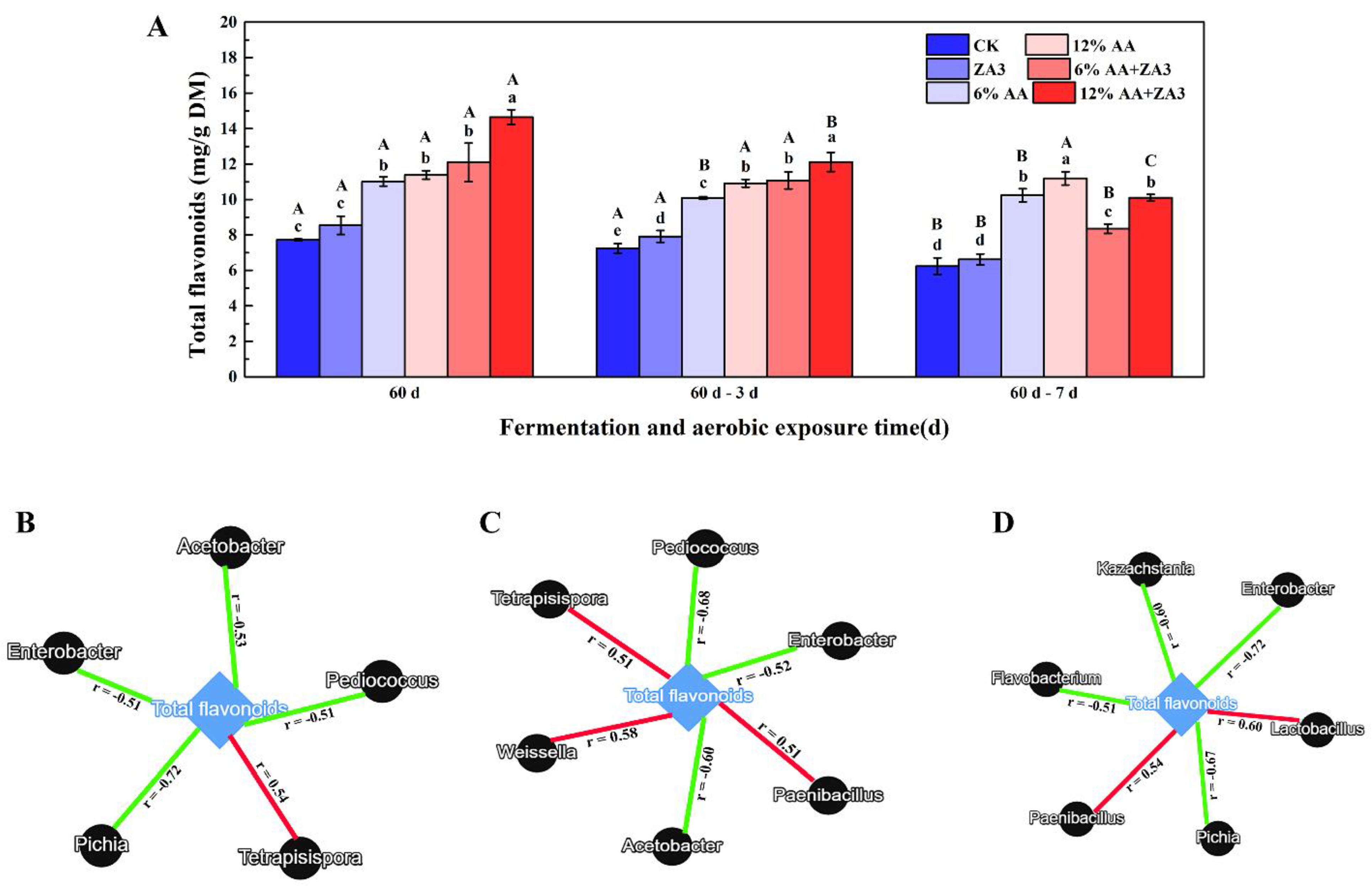
2.7. Total Flavonoid and Linkages with Microbial
3. Discussion
4. Materials and Methods
4.1. Materials Collection and Analysis
4.2. Lactic Acid Bacteria and Silage Preparation
4.3. Fermentation Quality, Microbial Population, and Chemical Composition of Silage
4.4. Bacterial and Fungi Community Analyses
4.5. Mycotoxin Determination
4.6. Total Flavonoid Measurement
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Borreani, G.; Tabacco, E.; Schmidt, R.J.; Holmes, B.J.; Muck, R.E. Silage review: Factors affecting dry matter and quality losses in silages. J. Dairy Sci. 2018, 101, 3952–3979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, T.W.; Adesogan, A.T.; Kim, S.C.; Lee, S.S. Effects of an esterase-producing inoculant on fermentation, aerobic stability, and neutral detergent fiber digestibility of corn silage. J. Dairy Sci. 2009, 92, 732–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrero, F.; Prencipe, S.; Spadaro, D.; Gullino, M.L.; Cavallarin, L.; Piano, S.; Tabacco, E.; Borreani, G. Increase in aflatoxins due to Aspergillus section Flavi multiplication during the aerobic deterioration of corn silage treated with different bacteria inocula. J. Dairy Sci. 2019, 102, 1176–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zychowski, K.E.; Pohlenz, C.; Mays, T.; Romoser, A.; Hume, M.; Buentello, A.; Gatlin, D.M.; Phillips, T.D. The effect of NovaSil dietary supplementation on the growth and health performance of Nile tilapia (Oreochromis niloticus) fed aflatoxin-B1 contaminated feed. Aquaculture 2013, 376–379, 117–123. [Google Scholar] [CrossRef]
- Kamala, A.; Kimanya, M.; Haesaert, G.; Tiisekwa, B.; Madege, R.; Degraeve, S.; Cyprian, C.; De Meulenaer, B. Local post-harvest practices associated with aflatoxin and fumonisin contamination of maize in three agro ecological zones of Tanzania. Food Addit. Contam. A 2016, 33, 551–559. [Google Scholar] [CrossRef]
- Athmanathan, A.; Emery, I.R.; Kuczek, T.; Mosier, N.S. Impact of temperature, moisture, and storage duration on the chemical composition of switchgrass, corn stover, and sweet sorghum bagasse. Bioenergy Res. 2014, 8, 843–856. [Google Scholar] [CrossRef]
- Muck, R. Silage microbiology and its control through additives. Rev. Bras. Zootecn. 2010, 39, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Ogunade, I.M.; Jiang, Y.; Kim, D.H.; Cervantes, A.A.P.; Arriola, K.G.; Vyas, D.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Fate of Escherichia coli O157:H7 and bacterial diversity in corn silage contaminated with the pathogen and treated with chemical or microbial additives. J. Dairy Sci. 2017, 100, 1780–1794. [Google Scholar] [CrossRef]
- Zhao, P.; Kim, I.H. Effect of direct-fed microbial on growth performance, nutrient digestibility, fecal noxious gas emission, fecal microbial flora and diarrhea score in weanling pigs. Anim. Feed Sci. Technol. 2015, 200, 86–92. [Google Scholar] [CrossRef]
- Fabiszewska, A.U.; Zielinska, K.J.; Wrobel, B. Trends in designing microbial silage quality by biotechnological methods using lactic acid bacteria inoculants: A minireview. World J. Microb. Biot. 2019, 35, 76. [Google Scholar] [CrossRef] [Green Version]
- Ni, K.; Minh, T.T.; Tu, T.T.; Tsuruta, T.; Pang, H.; Nishino, N. Comparative microbiota assessment of wilted Italian ryegrass, whole crop corn, and wilted alfalfa silage using denaturing gradient gel electrophoresis and next-generation sequencing. Appl. Microbiol. Biot. 2017, 101, 1385–1394. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ke, W.; Vyas, D.; Adesogan, A.T.; Franco, M.; Li, F.; Bai, J.; Guo, X. Antioxidant status, chemical composition and fermentation profile of alfalfa silage ensiled at two dry matter contents with a novel Lactobacillus plantarum strain with high-antioxidant activity. Anim. Feed Sci. Technol. 2021, 272, 114751. [Google Scholar] [CrossRef]
- Lee, D.; Kim, C.E.; Park, S.Y.; Kim, K.O.; Hiep, N.T.; Lee, D.; Jang, H.; Lee, J.; Kang, K. Protective effect of Artemisia argyi and its flavonoid constituents against contrast-induced cytotoxicity by iodixanol in LLC-PK1 Cells. Int. J. Mol. Sci. 2018, 19, 1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalaji, S.; Zaghari, M.; Hatami, K.; Hedari-Dastjerdi, S.; Lotfi, L.; Nazarian, H. Black cumin seeds, Artemisia leaves (Artemisia sieberi), and Camellia L. plant extract as phytogenic products in broiler diets and their effects on performance, blood constituents, immunity, and cecal microbial population. Poult. Sci. 2011, 90, 2500–2510. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Tan, Z.; Gu, L.; Ma, H.; Wang, Z.; Wang, L.; Wu, G.; Qin, G.; Wang, Y.; Pang, H. Dynamics changes of microorganisms community and fermentation quality in soybean meal prepared with lactic acid bacteria and Artemisia argyi through fermentation and aerobic exposure processes. Foods 2022, 11, 795. [Google Scholar] [CrossRef]
- He, L.; Zhou, W.; Wang, C.; Yang, F.; Chen, X.; Zhang, Q. Effect of cellulase and Lactobacillus casei on ensiling characteristics, chemical composition, antioxidant activity, and digestibility of mulberry leaf silage. J. Dairy Sci. 2019, 102, 9919–9931. [Google Scholar] [CrossRef]
- Mariotti, M.; Andreuccetti, V.; Nuvoloni, R.; Gatta, D.; Liponi, G.B.; Pedonese, F.; Fratini, F.; Turchi, B.; Saccomanni, G.; Torracca, B. Rutin and quercetin content in the forage of common buckwheat as affected by maturity and conservation method. Grassl. Sci. 2017, 63, 169–176. [Google Scholar] [CrossRef]
- Ehsan, O.; Norhani, A.; Rudi, H. Bioactive compounds, antioxidant, xanthine oxidase inhibitory, tyrosinase inhibitory and anti-inflammatory activities of selected agro-industrial by products. Int. J. Mol. Sci. 2011, 12, 8610–8625. [Google Scholar] [CrossRef]
- Martínez, R.G.; Porres, J.M.; Melesio, A.M.; Heras, L. Medicago sativa L., a functional food to relieve hypertension and metabolic disorders in a spontaneously hypertensive rat model. J. Funct. Foods 2016, 26, 470–484. [Google Scholar] [CrossRef]
- Guan, X.; Ge, D.; Li, S.; Huang, K.; Liu, J.; Li, F. Chemical composition and antimicrobial activities of Artemisia argyi Levl. et Vant essential oils extracted by simultaneous distillation-extraction, subcritical extraction and hydrodistillation. Molecules 2019, 24, 483. [Google Scholar] [CrossRef] [Green Version]
- Xiang, F.; Bai, J.; Tan, X.; Chen, T.; Yang, W.; He, F. Antimicrobial activities and mechanism of the essential oil from Artemisia argyi Levl. et Van. var. argyi cv. Qiai. Ind. Crop Prod. 2018, 125, 582–587. [Google Scholar] [CrossRef]
- Zhang, P.; Chen, H.; Shi, B.; Zhao, F.; Guo, X.; Jin, X.; Yan, S. In vitro antioxidant activity of Artemisia argyi powder and the effect on hepatic and intestinal antioxidant indices in broiler chickens. Ann. Anim. Sci. 2020, 20, 1085–1099. [Google Scholar] [CrossRef]
- Liu, L.; Zuo, W.; Li, F. Dietary addition of Artemisia argyi reduces diarrhea and modulates the gut immune function without affecting growth performances of rabbits after weaning. J. Anim. Sci. 2019, 97, 1693–1700. [Google Scholar] [CrossRef] [PubMed]
- Beigh, Y.A.; Ganai, A.M. Potential of wormwood (Artemisia absinthium Linn.) herb for use as additive in livestock feeding: A review. Pharma. Innov. 2017, 6, 176–187. [Google Scholar]
- Wilkins, R.J. The biochemistry of silage. Anim. Feed Sci. Technol. 1982, 7, 317–318. [Google Scholar] [CrossRef]
- Lin, C.; Bolsen, K.K.; Brent, B.E.; Hart, R.A.; Aimutis, W.R. Epiphytic microflora on alfalfa and whole-plant corn. J. Dairy Sci. 1992, 75, 2484–2493. [Google Scholar] [CrossRef]
- Kung, L.; Taylor, C.C.; Lynch, M.P.; Neylon, J.M. The Effect of treating alfalfa with Lactobacillus buchneri 40788 on silage fermentation, aerobic stability, and nutritive value for lactating dairy cows. J. Dairy Sci. 2003, 86, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Xu, D.; Xie, D.; Wang, M.; Li, Z.; Guo, X. Effects of antibacterial peptide-producing Bacillus subtilis and Lactobacillus buchneri on fermentation, aerobic stability, and microbial community of alfalfa silage. Bioresour. Technol. 2020, 315, 123881. [Google Scholar] [CrossRef]
- Bolsen, K.; Ashbell, G.; Weinberg, Z. Silage fermentation and silage additives-review. Asian Australas. J. Anim. 1996, 9, 483–494. [Google Scholar] [CrossRef]
- Wang, T.; Teng, K.; Cao, Y.; Shi, W.; Xuan, Z.; Zhou, J.; Zhang, J.; Zhong, J. Effects of Lactobacillus hilgardii 60TS-2, with or without homofermentative Lactobacillus plantarum B90, on the aerobic stability, fermentation quality and microbial community dynamics in sugarcane top silage. Bioresour. Technol. 2020, 312, 123600. [Google Scholar] [CrossRef]
- Kung, L., Jr.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef] [PubMed]
- Avila, C.L.; Carvalho, B.F.; Pinto, J.C.; Duarte, W.F.; Schwan, R.F. The use of Lactobacillus species as starter cultures for enhancing the quality of sugar cane silage. J. Dairy Sci. 2014, 97, 940–951. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Schmidt, R.J.; McDonell, E.E.; Klingerman, C.M.; Kung, L., Jr. The effect of Lactobacillus buchneri 40788 or Lactobacillus plantarum MTD-1 on the fermentation and aerobic stability of corn silages ensiled at two dry matter contents. J. Dairy Sci. 2009, 92, 3907–3914. [Google Scholar] [CrossRef] [PubMed]
- Ning, T.; Wang, H.; Zheng, M.; Niu, D.; Zuo, S.; Xu, C. Effects of microbial enzymes on starch and hemicellulose degradation in total mixed ration silages. Asian-Austral. J. Anim. Sci. 2017, 30, 171–180. [Google Scholar] [CrossRef]
- Liu, B.; Huang, H.; Gu, H.; Xu, N.; Shen, Q.; Ding, C. Dynamics of a microbial community during ensiling and upon aerobic exposure in lactic acid bacteria inoculation-treated and untreated barley silages. Bioresour. Technol. 2019, 273, 212–219. [Google Scholar] [CrossRef]
- Sonker, N.; Pandey, A.K.; Singh, P. Efficiency of Artemisia nilagirica (Clarke) Pamp. essential oil as a mycotoxicant against postharvest mycobiota of table grapes. J. Sci. Food Agric. 2015, 95, 1932–1939. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, X.; Gu, Q.; Liang, M.; Mu, S.; Zhou, B.; Huang, F.; Lin, B.; Zou, C. Analysis of the correlation between bacteria and fungi in sugarcane tops silage prior to and after aerobic exposure. Bioresour. Technol. 2019, 291, 121835. [Google Scholar] [CrossRef]
- Ni, K.; Wang, F.; Zhu, B.; Yang, J.; Zhou, G.; Pan, Y.; Tao, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresour. Technolo. 2017, 238, 706–715. [Google Scholar] [CrossRef]
- Ma, H.; Wang, W.; Wang, Z.; Tan, Z.; Qin, G.; Wang, Y.; Pang, H. Microbial population succession and community diversity and its correlation with fermentation quality in soybean meal treated with Enterococcus faecalis during fermentation and aerobic exposure. Microorganisms 2022, 10, 530. [Google Scholar] [CrossRef]
- Namihira, T.; Shinzato, N.; Akamine, H.; Maekawa, H.; Matsui, T. Influence of nitrogen fertilization on tropical-grass silage assessed by ensiling process monitoring using chemical and microbial community analyses. J. Appl. Microbiol. 2010, 108, 1954–1965. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela Saldinger, S. Microbiome dynamics during ensiling of corn with and without Lactobacillus plantarum inoculant. Appl. Microbiol. Biot. 2018, 102, 4025–4037. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ma, Q.; Ma, S.; Zhang, J.; Jia, R.; Ji, C.; Zhao, L. Ameliorating effects of Bacillus subtilis ANSB060 on growth performance, antioxidant functions, and aflatoxin residues in ducks fed diets contaminated with aflatoxins. Toxins 2016, 9, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanna-Filho, R.; Souza, R.M.; Alves, E. Induced resistance in tomato plants promoted by two endophytic bacilli against bacterial speck. Trop. Plant Pathol. 2017, 42, 96–108. [Google Scholar] [CrossRef]
- Santos, M.C.; Golt, C.; Joerger, R.D.; Mechor, G.D.; Mourao, G.B.; Kung, L., Jr. Identification of the major yeasts isolated from high moisture corn and corn silages in the United States using genetic and biochemical methods. J. Dairy Sci. 2017, 100, 1151–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Zhou, W.; Xing, Y.; Pian, R.; Chen, X.; Zhang, Q. Improving the quality of rice straw silage with Moringa oleifera leaves and propionic acid: Fermentation, nutrition, aerobic stability and microbial communities. Bioresour. Technol. 2020, 299, 122579. [Google Scholar] [CrossRef] [PubMed]
- Moon, N.J.; Ely, L.O.; Sudweeks, E.M. Aerobic deterioration of wheat, lucerne and maize silages prepared with Lactobacillus acidophilus and a Candida spp. J. Appl. Microbiol. 2008, 49, 75–87. [Google Scholar] [CrossRef]
- Duan, J.; Yin, J.; Wu, M.; Liao, P.; Deng, D.; Liu, G.; Wen, Q.; Wang, Y.; Qiu, W.; Liu, Y. Dietary glutamate supplementation ameliorates mycotoxin-induced abnormalities in the intestinal structure and expression of amino acid transporters in young pigs. PLoS ONE 2014, 9, e112357. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yang, X.; Li, J.; Dong, Z.; Zhao, J.; Shao, T.; Yuan, X. Effects of hexanoic acid on microbial communities, fermentation, and hygienic quality of corn silages infested with toxigenic fungi. J. Sci. Food Agric. 2021. [Google Scholar] [CrossRef]
- Li, J.; Wang, W.; Chen, S.; Shao, T.; Tao, X.; Yuan, X. Effect of lactic acid bacteria on the fermentation quality and mycotoxins concentrations of corn silage infested with mycotoxigenic Fungi. Toxins 2021, 13, 699. [Google Scholar] [CrossRef]
- Weiss, K.; Kroschewski, B.; Auerbach, H. Effects of air exposure, temperature and additives on fermentation characteristics, yeast count, aerobic stability and volatile organic compounds in corn silage. J. Dairy Sci. 2016, 99, 8053–8069. [Google Scholar] [CrossRef] [Green Version]
- Mugabe, W.; Yuan, X.; Li, J.; Dong, Z.; Shao, T. Effects of hexanoic acid, Lactobacillus plantarum and their combination on the fermentation characteristics of Napier grass. Anim. Feed Sci. Technol. 2019, 253, 135–140. [Google Scholar] [CrossRef]
- Alencar, S.M.; Oldoni, T.L.; Castro, M.L.; Cabral, I.S.; Costa-Neto, C.M.; Cury, J.A.; Rosalen, P.L.; Ikegaki, M. Chemical composition and biological activity of a new type of Brazilian propolis: Red propolis. J. Ethnopharmacol. 2007, 113, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Koru, O.; Toksoy, F.; Acikel, C.H.; Tunca, Y.M.; Baysallar, M.; Uskudar Guclu, A.; Akca, E.; Ozkok Tuylu, A.; Sorkun, K.; Tanyuksel, M. In vitro antimicrobial activity of propolis samples from different geographical origins against certain oral pathogens. Anaerobe 2007, 13, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Pok, P.S.; Garcia Londono, V.A.; Vicente, S.; Pacin, A.; Alzamora, S.M.; Resnik, S.L. Citrus flavonoids against Fusarium verticillioides in post-harvest maize: Minimization of fumonisins and alteration of fungal ultrastructure. J. Appl. Microbiol. 2021, 132, 2234–2248. [Google Scholar] [CrossRef] [PubMed]
- Simard, F.; Gauthier, C.; Legault, J.; Lavoie, S.; Mshvildadze, V.; Pichette, A. Structure elucidation of anti-methicillin resistant Staphylococcus aureus (MRSA) flavonoids from balsam poplar buds. Bioorg. Med. Chem. 2016, 24, 4188–4198. [Google Scholar] [CrossRef] [PubMed]
- Ullah, R.; Ahmad, S.; Atiq, A.; Hussain, H.; Ur Rehman, N.; Abd Elsalam, N.M.; Adnan, M. Quantification and antibacterial activity of flavonoids in coffee samples. Afr. J. Tradit. Complem. 2015, 12, 84. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Cao, S.; Wang, C.; Wang, K.; Wei, Y.; Shao, X.; Wang, H. Antimicrobial activity of flavonoids from Sedum aizoon L. against Aeromonas in culture medium and in frozen pork. Food Sci. Nutr. 2019, 7, 3224–3232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Li, H.; Qin, X.; Wang, T.; Hao, J.; Zhao, J.; Wang, J.; Wang, R.; Wang, D.; Wei, S. Antibacterial evaluation of plants extracts against ampicillin-resistant Escherichia coli (E. coli) by microcalorimetry and principal component analysis. AMB Express 2019, 9, 101. [Google Scholar] [CrossRef]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- Arthur Thomas, T. An automated procedure for the determination of soluble carbohydrates in herbage. J. Sci. Food Agric. 1977, 28, 639–642. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Agricultural Chemists: Rockville, MD, USA, 1990. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary Fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Pang, H.; Qin, G.; Tan, Z.; Li, Z.; Wang, Y.; Cai, Y. Natural populations of lactic acid bacteria associated with silage fermentation as determined by phenotype, 16S ribosomal RNA and recA gene analysis. Syst. Appl. Microbiol. 2011, 34, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ma, H.; Yu, H.; Qin, G.; Tan, Z.; Wang, Y.; Pang, H. Screening of Lactobacillus plantarum subsp. plantarum with potential probiotic activities for inhibiting ETEC K88 in weaned piglets. Molecules 2020, 25, 4481. [Google Scholar] [CrossRef] [PubMed]
- Hudz, N.; Yezerska, O.; Grygorieva, O.; Brindza, J.; Felsöciová, S.; Kačániová, M.; Wieczorek, P. Analytical procedure elaboration of total flavonoid content determination and antimicrobial activity of bee bread extracts. Acta Pol. Pharm. 2019, 76, 439–452. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | WCC | AA |
|---|---|---|
| pH | 6.61 ± 0.05 | 5.58 ± 0.01 |
| DM (g/kg) | 277.03 ± 3.29 | 371.55 ± 3.77 |
| WSC (% DM) | 12.12 ± 0.22 | 5.41 ± 0.25 |
| CP (% DM) | 9.43 ± 0.31 | 10.92 ± 0.02 |
| NDF (% DM) | 60.07 ± 0.56 | 53.76 ± 0.84 |
| ADF (% DM) | 35.66 ± 0.78 | 30.42 ± 0.51 |
| EE (% DM) | 2.51 ± 0.06 | 3.06 ± 0.08 |
| Microbial population (lg cfu/g FM) | ||
| LAB | 6.33 ± 0.17 | 5.59 ± 0.08 |
| Aerobic bacteria | 8.32 ± 0.24 | 5.84 ± 0.05 |
| Coliform bacteria | 7.81 ± 0.15 | 5.63 ± 0.46 |
| Yeast | 5.76 ± 0.09 | ND |
| Bacilli | 4.68 ± 0.13 | 6.61 ± 0.01 |
| Item | Treatment | pH | NH3-N (% TN) | Organic Acid (g/Kg DM) | WSC (%DM) | |
|---|---|---|---|---|---|---|
| Lactic Acid | Acetic Acid | |||||
| 60 d ensiling | CK | 3.73 ± 0.02 bC | 16.11 ± 0.23 aB | 25.96 ± 0.97 bA | 3.16 ± 0.3 b | 1.51 ± 0.01 aA |
| ZA3 | 3.73 ± 0.01 cC | 15.42 ± 0.11 aB | 32.90 ± 1.57 aA | 3.39 ± 0.33 ab | 1.43 ± 0.01 aA | |
| 6% AA | 3.75 ± 0.01 aC | 14.63 ± 0.41 abA | 26.40 ± 1.67 bA | 3.57 ± 0.38 ab | 1.45 ± 0.02 aB | |
| 12% AA | 3.70 ± 0.01 bB | 19.21 ± 0.22 bA | 32.44 ± 1.24 aA | 3.87 ± 0.25 aB | 1.72 ± 0.03 aA | |
| 6% AA+ZA3 | 3.52 ± 0.02 dC | 12.42 ± 0.64 bcA | 33.18 ± 0.40 aA | ND | 1.44 ± 0.01 aA | |
| 12% AA+ZA3 | 3.52 ± 0.01 dC | 12.07 ± 0.51 cAB | 27.05 ± 1.60 bA | ND | 1.09 ± 0.01 bC | |
| aerobic exposure for 3 d | CK | 5.14 ± 0.09 aB | 21.12 ± 0.33 aC | 9.71 ± 0.69 cB | ND | 1.16 ± 0.01 aB |
| ZA3 | 4.34 ± 0.1 bB | 19.06 ± 0.52 bB | 8.58 ± 1.37 cB | ND | 1.61 ± 0.02 aA | |
| 6% AA | 3.90 ± 0.02 dB | 17.56 ± 0.22 bcA | 19.29 ± 4.45 bB | ND | 1.16 ± 0.03 aB | |
| 12% AA | 3.70 ± 0.03 eB | 13.17 ± 0.21 cB | 41.45 ± 8.66 aA | 6.52 ± 1.53 aA | 1.27 ± 0.02 aA | |
| 6% AA+ZA3 | 4.19 ± 0.1 cB | 13.63 ± 0.63 cB | 17.14 ± 1.53 bB | ND | 1.29 ± 0.01 aA | |
| 12% AA+ZA3 | 4.44 ± 0.03 bB | 14.22 ± 0.92 cB | 4.67 ± 0.16 dB | ND | 1.35 ± 0.01 aB | |
| aerobic exposure for 7 d | CK | 7.74 ± 0.1 aA | 41.09 ± 0.41 aA | ND | ND | 1.63 ± 0.04 aA |
| ZA3 | 6.98 ± 0.06 bA | 32.51 ± 0.63 bA | ND | ND | 1.48 ± 0.03 aA | |
| 6% AA | 6.48 ± 0.1 cA | 24.32 ± 0.79 bcA | 4.56 ± 0.44 bC | ND | 1.11 ± 0.07 bA | |
| 12% AA | 4.64 ± 0.16 dA | 19.93 ± 0.18 cB | 7.48 ± 1.71 aB | ND | 0.50 ± 0.01 cB | |
| 6% AA+ZA3 | 6.66 ± 0.01 cA | 24.47 ± 0.74 bcA | ND | ND | 1.63 ± 0.05 aA | |
| 12% AA+ZA3 | 6.54 ± 0.04 cA | 22.29 ± 0.52 bcA | ND | ND | 1.23 ± 0.04 bA | |
| SEM | 0.01 | 0.006 | 0.342 | 0.053 | 0.019 | |
| Period | *** | *** | *** | *** | ** | |
| Treatment | *** | *** | *** | *** | ** | |
| Interaction | *** | *** | *** | *** | *** | |
| Item | Treatment | Chemical Composition (%DM) | ||||
|---|---|---|---|---|---|---|
| CP | NDF | ADF | EE | DM | ||
| 60 d ensiling | CK | 9.85 ± 0.34 aA | 60.75 ± 0.62 aA | 38.15 ± 0.67 aC | 2.59 ± 0.02 cA | 212.08 ± 6.47 cA |
| ZA3 | 9.60 ± 0.14 aA | 59.05 ± 0.79 abA | 36.95 ± 0.58 abB | 2.77 ± 0.05 dA | 211.42 ± 7.44 cB | |
| 6% AA | 9.67 ± 0.17 aA | 56.08 ± 0.98 cA | 35.86 ± 0.10 bcB | 2.93 ± 0.06 cAB | 267.26 ± 6.86 bB | |
| 12% AA | 10.11 ± 0.10 aA | 56.33 ± 1.01 cA | 36.11 ± 0.62 bcB | 3.04 ± 0.12 abB | 294.29 ± 7.97 aA | |
| 6% AA+ZA3 | 9.72 ± 0.28 aA | 57.79 ± 0.91 bcA | 34.86 ± 0.71 cB | 2.92 ± 0.08 cB | 256.91 ± 6.22 bA | |
| 12% AA+ZA3 | 9.76 ± 0.21 aA | 58.00 ± 0.52 bcA | 35.48 ± 0.27 bcB | 3.16 ± 0.06 aA | 301.87 ± 9.71 aA | |
| aerobic exposure for 3 d | CK | 9.04 ± 0.07 bB | 59.26 ± 1.13 aA | 37.76 ± 0.61 abB | 2.53 ± 0.01 eA | 220.99 ± 4.83 cA |
| ZA3 | 9.64 ± 0.02 abA | 59.08 ± 0.11 aA | 37.85 ± 1.24 aB | 2.76 ± 0.14 dA | 228.26 ± 6.91 cA | |
| 6% AA | 9.76 ± 0.12 aA | 56.65 ± 0.74 bA | 35.42 ± 1.21 cB | 2.84 ± 0.13 cdB | 274.87 ± 4.02 bB | |
| 12% AA | 10.06 ± 0.43 aA | 56.60 ± 0.49 bA | 35.31 ± 0.29 cB | 2.99 ± 0.07 bcB | 305.32 ± 6.99 aA | |
| 6% AA+ZA3 | 9.47 ± 0.45 abA | 58.37 ± 0.66 abA | 35.47 ± 0.43 bcB | 3.13 ± 0.07 abA | 268.64 ± 4.06 bA | |
| 12% AA+ZA3 | 10.03 ± 0.27 aA | 57.86 ± 0.85 abA | 36.15 ± 0.76 abcB | 3.28 ± 0.11 aA | 303.05 ± 3.97 aA | |
| aerobic exposure for 7 d | CK | 8.92 ± 0.08 cB | 58.53 ± 0.74 aB | 39.46 ± 0.68 aA | 2.49 ± 0.10 bA | 209.39 ± 1.07 dA |
| ZA3 | 8.98 ± 0.23 bcB | 58.41 ± 0.93 aB | 39.65 ± 0.49 aA | 2.53 ± 0.11 bA | 210.55 ± 5.88 dB | |
| 6% AA | 9.31 ± 0.07 bcB | 56.06 ± 0.48 bA | 37.94 ± 0.87 abA | 3.14 ± 0.11 aA | 293.97 ± 6.94 bA | |
| 12% AA | 10.05 ± 0.07 aA | 56.01 ± 0.88 bA | 37.33 ± 0.68 bA | 3.24 ± 0.02 aA | 318.23 ± 11.12 aA | |
| 6% AA+ZA3 | 9.09 ± 0.19 bcA | 56.47 ± 1.17 abB | 38.51 ± 0.40 aA | 3.15 ± 0.11 aA | 277.24 ± 3.84 cA | |
| 12% AA+ZA3 | 9.40 ± 0.31 bA | 56.73 ± 0.39 abB | 38.35 ± 0.70 aA | 3.29 ± 0.11 aA | 309.02 ± 7.94 abA | |
| SEM | 0.03 | 0.106 | 0.091 | 0.012 | 0.172 | |
| Period | ** | * | ** | * | * | |
| Treatment | *** | ** | ** | *** | *** | |
| Interaction | ** | ** | ** | *** | *** | |
| Item | Treatment | Microbial Population (lg cfu/g FM) | ||||
|---|---|---|---|---|---|---|
| Lactic Acid Bacteria | Aerobic Bacteria | Coliform Bacteria | Yeast | Bacilli | ||
| 60 d ensiling | CK | 6.97 ± 0.05 cA | 5.64 ± 0.21 aC | 3.96 ± 0.24 bC | 5.11 ± 0.14 aB | 3.70 ± 0.01 aC |
| ZA3 | 8.02 ± 0.03 aA | 5.36 ± 0.30 aC | 4.16 ± 0.41 bB | 4.25 ± 0.52 bC | ND | |
| 6% AA | 6.83 ± 0.13 cA | 5.31 ± 0.11 aC | 4.28 ± 0.27 bC | 4.88 ± 0.07 aC | 4.44 ± 0.02 bC | |
| 12% AA | 7.48 ± 0.06 bA | 5.68 ± 0.05 aB | 4.20 ± 0.17 bC | 4.76 ± 0.19 aC | 4.00 ± 0.13 cB | |
| 6% AA+ZA3 | 6.51 ± 0.06 cA | 5.75 ± 0.14 aC | 4.99 ± 0.16 aC | 5.09 ± 0.14 aC | 4.57 ± 0.02 bC | |
| 12% AA+ZA3 | 6.35 ± 0.07 cA | 5.65 ± 0.12 aC | 5.10 ± 0.12 aC | 4.90 ± 0.19 aC | 5.39 ± 0.11 aB | |
| aerobic exposure for 3 d | CK | 4.27 ± 0.11 dB | 8.14 ± 0.14 aB | 7.81 ± 0.12 aB | 10.17 ± 0.12 aA | 5.10 ± 0.06 aB |
| ZA3 | 6.02 ± 0.15 bB | 7.36 ± 0.12 bcB | 4.56 ± 0.23 bB | 9.28 ± 0.14 bB | 3.90 ± 0.12 cB | |
| 6% AA | 6.03 ± 0.10 bA | 7.24 ± 0.27 cB | 5.10 ± 0.40 bB | 9.20 ± 0.26 bB | 4.93 ± 0.17 aB | |
| 12% AA | 7.12 ± 0.09 aA | 4.93 ± 0.11 dB | 5.00 ± 0.27 bB | 7.61 ± 0.33 cB | 3.70 ± 0.13 cC | |
| 6% AA+ZA3 | 5.03 ± 0.07 cB | 7.40 ± 0.05 bcB | 7.34 ± 0.31 aB | 9.49 ± 0.06 bB | 5.16 ± 0.04 aB | |
| 12% AA+ZA3 | 4.93 ± 0.04 cB | 7.68 ± 0.10 bB | 7.39 ± 0.26 aB | 9.39 ± 0.14 bB | 4.71 ± 0.14 bC | |
| aerobic exposure for 7 d | CK | ND | 10.09 ± 0.04 aA | 10.13 ± 0.08 aA | 10.20 ± 0.15 aA | 7.45 ± 0.21 aA |
| ZA3 | ND | 10.18 ± 0.01 aA | 9.79 ± 0.16 bA | 9.94 ± 0.04 bA | 7.36 ± 0.32 abA | |
| 6% AA | 3.30 ± 0.15 bB | 9.82 ± 0.18 bA | 9.57 ± 0.05 bcA | 9.85 ± 0.05 bA | 7.31 ± 0.07 abA | |
| 12% AA | 4.73 ± 0.09 aB | 9.46 ± 0.11 cA | 9.09 ± 0.14 dA | 9.20 ± 0.20 cA | 6.32 ± 0.16 cA | |
| 6% AA+ZA3 | ND | 9.62 ± 0.26 bcA | 9.59 ± 0.21 bcA | 9.86 ± 0.20 bA | 7.26 ± 0.11 abA | |
| 12% AA+ZA3 | ND | 9.75 ± 0.20 bA | 9.47 ± 0.08 cA | 9.78 ± 0.08 bA | 7.15 ± 0.27 bA | |
| SEM | 0.008 | 0.02 | 0.031 | 0.027 | 0.014 | |
| Period | *** | *** | *** | *** | *** | |
| Treatment | *** | *** | *** | *** | *** | |
| Interaction | *** | *** | *** | *** | *** | |
| Item | Treatment | Bacteria | Fungi | ||
|---|---|---|---|---|---|
| Shannon | Chao1 | Shannon | Chao1 | ||
| 60 d ensiling | CK | 4.37 ± 0.18 aB | 2717.73 ± 168.19 bA | 2.50 ± 0.12 bA | 126.42 ± 10.99 bA |
| ZA3 | 2.97 ± 0.66 cB | 1543.12 ± 209.71 cB | 1.96 ± 0.15 cA | 127.65 ± 8.99 bA | |
| 6% AA | 3.19 ± 0.17 bB | 2289.73 ± 211.50 bA | 1.76 ± 0.18 cA | 132.53 ± 12.62 bA | |
| 12% AA | 3.56 ± 0.31 bB | 2436.86 ± 234.93 bA | 1.94 ± 0.04 cA | 130.49 ± 7.81 bB | |
| 6% AA+ZA3 | 4.23 ± 0.22 aC | 2745.72 ± 244.11 bB | 3.22 ± 0.07 aA | 160.47 ± 5.92 aB | |
| 12% AA+ZA3 | 4.65 ± 0.30 aB | 3746.15 ± 81.39 aA | 2.46 ± 0.17 bA | 164.33 ± 6.81 aB | |
| aerobic exposure for 3 d | CK | 2.25 ± 0.23 cC | 2100.83 ± 200.52 bA | 1.69 ± 0.07 aB | 146.67 ± 5.82 bA |
| ZA3 | 2.60 ± 0.36 cB | 1550.55 ± 129.86 cB | 1.31 ± 0.15 bB | 142.42 ± 2.03 bA | |
| 6% AA | 3.32 ± 0.56 bB | 1816.18 ± 159.40 cB | 1.35 ± 0.10 bB | 114.67 ± 2.52 cA | |
| 12% AA | 3.73 ± 0.22 bB | 1886.84 ± 119.26 cB | 1.82 ± 0.19 aA | 111.33 ± 1.53 cA | |
| 6% AA+ZA3 | 4.31 ± 0.16 abB | 2791.93 ± 182.80 bB | 2.29 ± 0.14 aB | 223.94 ± 3.28 aA | |
| 12% AA+ZA3 | 5.16 ± 0.19 aA | 4732.19 ± 178.50 aA | 1.85 ± 0.07 aB | 264.45 ± 11.09 aA | |
| aerobic exposure for 7 d | CK | 5.21 ± 0.28 aA | 2407.26 ± 154.68 bA | 2.36 ± 0.12 aA | 74.50 ± 2.50 bB |
| ZA3 | 5.34 ± 0.34 aA | 2249.07 ± 184.11 bA | 2.08 ± 0.06 aA | 99.50 ± 4.77 bB | |
| 6% AA | 4.31 ± 0.24 bA | 2096.86 ± 68.13 cA | 1.67 ± 0.21 bA | 65.33 ± 2.52 cB | |
| 12% AA | 4.53 ± 0.13 bA | 2161.91 ± 46.62 cA | 1.82 ± 0.03 bA | 54.61 ± 2.85 cB | |
| 6% AA+ZA3 | 5.17 ± 0.25 aA | 3131.24 ± 154.94 aA | 2.46 ± 0.12 aB | 127.48 ± 7.88 aB | |
| 12% AA+ZA3 | 5.72 ± 0.30 aA | 3264.59 ± 105.54 aA | 2.47 ± 0.18 aA | 141.33 ± 10.50 aB | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Tan, Z.; Gu, L.; Ma, H.; Wang, Z.; Wang, L.; Wu, G.; Qin, G.; Wang, Y.; Pang, H. Variation of Microbial Community and Fermentation Quality in Corn Silage Treated with Lactic Acid Bacteria and Artemisia argyi during Aerobic Exposure. Toxins 2022, 14, 349. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14050349
Wang W, Tan Z, Gu L, Ma H, Wang Z, Wang L, Wu G, Qin G, Wang Y, Pang H. Variation of Microbial Community and Fermentation Quality in Corn Silage Treated with Lactic Acid Bacteria and Artemisia argyi during Aerobic Exposure. Toxins. 2022; 14(5):349. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14050349
Chicago/Turabian StyleWang, Weiwei, Zhongfang Tan, Lingbiao Gu, Hao Ma, Zhenyu Wang, Lei Wang, Guofang Wu, Guangyong Qin, Yanping Wang, and Huili Pang. 2022. "Variation of Microbial Community and Fermentation Quality in Corn Silage Treated with Lactic Acid Bacteria and Artemisia argyi during Aerobic Exposure" Toxins 14, no. 5: 349. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins14050349