
Effects of Wheat Naturally Contaminated with Fusarium Mycotoxins on Growth Performance and Selected Health Indices of Red Tilapia (Oreochromis niloticus × O. mossambicus)

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Composition of Experimental Diets
2.2. Determination of Mycotoxins and Fungal Metabolites in Diets
2.3. Growth Performance

{kind=link}
| Diet | DON | WG a | TGC b | FI c | FE d | Mortality |
|---|---|---|---|---|---|---|
| (mg·kg−1) | (g/Fish) | (g/Fish) | (Gain/Feed) | (%) | ||
| 1 (0.0% e) | 0.07 | 37.4 | 0.108 | 52.5 | 0.71 | 12 |
| 2 (1.2%) | 0.31 | 34.9 | 0.104 | 49.5 | 0.70 | 14 |
| 3 (2.5%) | 0.50 | 34.0 | 0.102 | 49.5 | 0.69 | 17 |
| 4 (5.0%) | 0.92 | 31.6 | 0.099 | 47.2 | 0.67 | 10 |
| 5 (10.0%) | 1.15 | 29.6 | 0.095 | 45.8 | 0.65 | 6 |
| Significance f | ||||||
| Linear | p < 0.01 | p < 0.01 | p < 0.01 | p < 0.01 | p < 0.05 | |
| Quadratic | N.S. g | N.S. | N.S. | N.S. | p < 0.05 | |
| S.E.M h | 1.3 | 0.002 | 1.3 | 0.01 | 2.2 | |
2.4. Hematological and Biochemical Parameters
| Diet | DON a | AST b | ALT c | Hct d | HSI e |
|---|---|---|---|---|---|
| (mg·kg−1) | IU/L | IU/L | (%) | (%) | |
| 1 (0.0% f) | 0.07 | 592 | 96 | 34 | 2.40 |
| 2 (1.2%) | 0.31 | 477 | 69 | 32 | 2.18 |
| 3 (2.5%) | 0.50 | 574 | 103 | 33 | 1.94 |
| 4 (5.0%) | 0.92 | 350 | 90 | 32 | 1.87 |
| 5 (10.0%) | 1.15 | 527 | 146 | 31 | 1.97 |
| Significance g | |||||
| Linear | N.S. h | N.S. | N.S. | N.S. | |
| Quadratic | N.S. | N.S. | N.S. | N.S. | |
| S.E.M. i | 142 | 38 | 1.5 | 0.18 | |
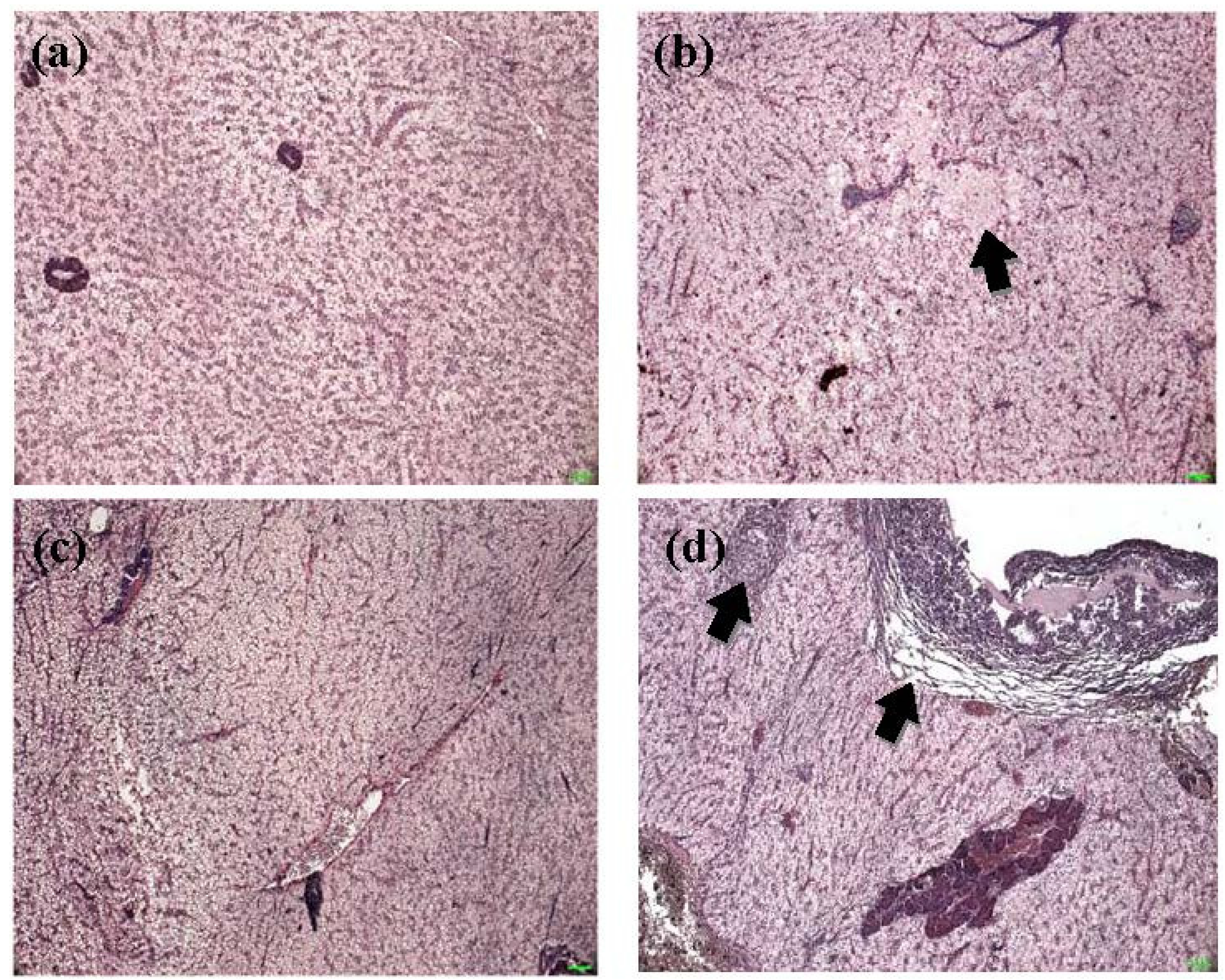
2.5. Histopathological alterations in livers

3. Experimental Section
3.1. Experimental Condition
3.2. Experimental Animal
3.3. Experimental Diets
| Ingredients | Experimental Diets | ||||
|---|---|---|---|---|---|
| (100%) | 1 | 2 | 3 | 4 | 5 |
| Cassava | 15.0 | 15.0 | 15.0 | 15.0 | 15.0 |
| Soybean meal | 25.0 | 25.0 | 25.0 | 25.0 | 25.0 |
| Clean wheat a | 19.6 | 18.4 | 17.1 | 14.6 | 9.6 |
| Contaminated wheat b | - | 1.2 | 2.5 | 5.0 | 10.0 |
| Rice bran | 20.0 | 20.0 | 20.0 | 20.0 | 20.0 |
| Fish meal c | 15.0 | 15.0 | 15.0 | 15.0 | 15.0 |
| Vitamin premix d | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 |
| Fish oil | 2.0 | 2.0 | 2.0 | 2.0 | 2.0 |
| Soybean oil | 3.0 | 3.0 | 3.0 | 3.0 | 3.0 |
| Total | 100 | 100 | 100 | 100 | 100 |
| Proximate analysis (% dry matter basis) | |||||
| Dry matter (%) | 94.4 | 95.6 | 94.9 | 95.5 | 94.6 |
| Crude protein (%) | 25.1 | 25.2 | 24.3 | 24.5 | 24.1 |
| Lipid (%) | 6.7 | 6.7 | 6.9 | 6.9 | 7.0 |
| Ash (%) | 8.5 | 8.5 | 8.5 | 8.5 | 8.6 |
| Gross energy (kJ/g) | 17.8 | 17.9 | 17.9 | 17.8 | 17.8 |
| Fusarium mycotoxins (mg·kg−1 diet) | |||||
| Deoxynivalenol (DON) | 0.07 | 0.31 | 0.50 | 0.92 | 1.15 |
| Zearalenone (ZEN) | 0.01 | 0.09 | 0.21 | 0.37 | 0.98 |
| Other Fusarium metabolites (mg·kg−1 diet) | |||||
| Aurofusarin | 0.01 | 0.30 | 0.66 | 1.28 | 2.46 |
| Rubrofusarin | 0.02 | 0.05 | 0.12 | 0.23 | 0.49 |
| Culmorin | 0.02 | 0.18 | 0.26 | 0.50 | 1.39 |
| 15-Hydroxyculmorin | 0.06 | 0.24 | 0.37 | 0.67 | 1.83 |
| Alternaria metabolites (mg·kg−1 diet) | |||||
| Alternariol | 0.001 | 0.01 | 0.02 | 0.04 | 0.12 |
| Alternariolmonomethyle-ther | 0.01 | 0.02 | 0.04 | 0.06 | 0.14 |
3.4. Mycotoxin Analysis of Dietary Samples
3.5. Sample Collection
3.6. Laboratory Analysis
3.7. Histopathological Examination
3.8. Calculations and Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2014; Food and Agriculture Organization of the United Nations: Rome, Italy, 2014; p. 223. [Google Scholar]
- Rana, K.J.; Siriwardena, S.; Hasan, M.R. Impact of Rising Feed Ingredient Prices on Aquafeeds and Aquaculture Production Food and Agriculture Organization of the United Nations (FAO); FAO Fisheries and Aquaculture Technical Paper; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2009. [Google Scholar]
- Tacon, A.G.J.; Metian, M.; Hasan, M.R. Feed Ingredients and Fertilizers for Farmed Aquatic Animals: Sources and Composition; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2009. [Google Scholar]
- Binder, E.M.; Tan, L.M.; Chin, L.J.; Handl, J.; Richard, J. Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Technol. 2007, 137, 265–282. [Google Scholar] [CrossRef]
- Placinta, C.M.; D’Mello, J.P.F.; Macdonald, A.M.C. A review of worldwide contamination of cereal grains and animal feed with Fusarium mycotoxins. Anim. Feed Sci. Technol. 1999, 78, 21–37. [Google Scholar] [CrossRef]
- Rodrigues, I.; Handl, J.; Binder, E.M. Mycotoxin occurrence in commodities, feeds and feed ingredients sourced in the Middle East and Africa. Food Addit. Contam. Part B 2011, 4, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, I.; Nährer, K. A three-year survey on the worldwide occurrence of mycotoxins in feedstuffs and feed. Toxins 2012, 4, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, C.; Kersten, S.; Burkhardt-Holm, P.; Valenta, H.; Dänicke, S. Occurrence of deoxynivalenol and zearalenone in commercial fish feed: An initial study. Toxins 2013, 5, 184–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Council for Agricultural Science and Technology (CAST). Mycotoxins: Risks in Plant, Animal and Human Systems; CAST: Ames, IA, USA, 2003. [Google Scholar]
- Leung, M.C.K.; Díaz-Llano, G.; Smith, T.K. Mycotoxins in pet food: A review on worldwide prevalence and preventative strategies. J. Agric. Food Chem. 2006, 54, 9623–9635. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.D. Fungi and mycotoxins in grain: Implications for stored product research. J. Stored Prod. Res. 1995, 31, 1–16. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Toxicity, mechanisms and animal health risks. Anim. Feed Sci. Technol. 2007, 137, 283–298. [Google Scholar] [CrossRef]
- Nahrer, K.; Kovalsky, K. Biomin Mycotoxin Survey: A Summary of the Major Threats. Available online: http://www.biomin.net/uploads/tx_news/ART_No09_MYC_EN_0214.pdf (accessed on 20 November 2013).
- Rodrigues, I.; Nährer, K. Prevalence of mycotoxins in feedstuffs and feed surveyed worldwide in 2009 and 2010. Phytopathol. Mediterr. 2012, 51, 175–192. [Google Scholar]
- Ueno, Y. Toxicological features of T-2 toxin and related trichothecenes. Fundam. Appl. Toxicol. 1984, 4, S124–S132. [Google Scholar] [CrossRef]
- Dänicke, S.; Goyarts, T.; Döll, S.; Grove, N.; Spolders, M.; Flachowsky, G. Effects of the Fusarium toxin deoxynivalenol on tissue protein synthesis in pigs. Toxicol. Lett. 2006, 165, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Friend, D.W.; Trenholm, H.L.; Elliot, J.I.; Hartin, K.E.; Thompson, B.K. Effect of feeding vomitoxin-contaminated wheat to pigs. Can. J. Anim. Sci. 1982, 62, 1211–1222. [Google Scholar] [CrossRef]
- Tiemann, U.; Brüssow, K.P.; Küchenmeister, U.; Jonas, L.; Kohlschein, P.; Pöhland, R.; Dänicke, S. Influence of diets with cereal grains contaminated by graded levels of two Fusarium toxins on selected enzymatic and histological parameters of liver in gilts. Food Chem. Toxicol. 2006, 44, 1228–1235. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.K.; MacDonald, E.J. Effect of fusaric acid on brain regional neurochemistry and vomiting behavior in swine. J. Anim. Sci. 1991, 69, 2044–2049. [Google Scholar] [PubMed]
- Fitzpatrick, D.W.; Boyd, K.E.; Watts, B.M. Comparison of the trichothecenes deoxynivalenol and T-2 toxin for their effects on brain biogenic monoamines in the rat. Toxicol. Lett. 1988, 40, 241–245. [Google Scholar] [CrossRef]
- Manning, B.B.; Abbas, H.K.; Wise, D.J.; Greenway, T. The effect of feeding diets containing deoxynivalenol contaminated corn on channel catfish (Ictalurus punctatus) challenged with Edwardsiella ictaluri. Aquac. Res. 2013, 1–5. [Google Scholar]
- Pestka, J.J.; Zhou, H.-R.; Moon, Y.; Chung, Y.J. Cellular and molecular mechanisms for immune modulation by deoxynivalenol and other trichothecenes: Unraveling a paradox. Toxicol. Lett. 2004, 153, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Rotter, B.A. Invited review: Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health Part A 1996, 48, 1–34. [Google Scholar] [CrossRef]
- Hooft, J.M.; Elmor, A.E.H.I.; Encarnação, P.; Bureau, D.P. Rainbow trout (Oncorhynchus mykiss) is extremely sensitive to the feed-borne Fusarium mycotoxin deoxynivalenol (DON). Aquaculture 2011, 311, 224–232. [Google Scholar] [CrossRef]
- Forsyth, D.M.; Yoshizawa, T.; Morooka, N.; Tuite, J. Emetic and refusal activity of deoxynivalenol to swine. Appl. Environ. Microbiol. 1977, 34, 547–552. [Google Scholar] [PubMed]
- Young, L.G.; McGirr, L.; Valli, V.E.; Lumsden, J.H.; Lun, A. Vomitoxin in corn fed to young pigs. J. Anim. Sci. 1983, 57, 655–664. [Google Scholar] [PubMed]
- Swamy, H.V.; Smith, T.K.; Karrow, N.A.; Boermans, H.J. Effects of feeding blends of grains naturally contaminated with Fusarium mycotoxins on growth and immunological parameters of broiler chickens. Poult. Sci. 2004, 83, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Ingalls, J.R. Influence of deoxynivalenol on feed consumption by dairy cows. Anim. Feed Sci. Technol. 1996, 60, 297–300. [Google Scholar] [CrossRef]
- Woodward, B.; Young, L.G.; Lun, A.K. Vomitoxin in diets for rainbow trout (Salmo gairdneri). Aquaculture 1983, 35, 93–101. [Google Scholar] [CrossRef]
- Manning, B. Mycotoxin in aquaculture. In The Mycotoxin Blue Book; Diaz, D., Ed.; Nottingham University Press: Nottingham, UK, 2005; pp. 139–156. [Google Scholar]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-Mycotoxin screening reveals the occurrence of 139 different secondary metabolites in feed and feed ingredients. Toxin 2013, 5, 504–523. [Google Scholar] [CrossRef] [PubMed]
- Abia, W.A.; Warth, B.; Sulyok, M.; Krska, R.; Tchana, A.N.; Njobeh, P.B.; Dutton, M.F.; Moundipa, P.F. Determination of multi-mycotoxin occurrence in cereals, nuts and their products in Cameroon by liquid chromatography tandem mass spectrometry (LC-MS/MS). Food Control 2013, 31, 438–453. [Google Scholar] [CrossRef]
- Ghebremeskel, M.; Langseth, W. The occurrence of culmorin and hydroxy-culmorins in cereals. Mycopathologia 2001, 152, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Langseth, W. Mycotoxin production and cytotoxicity of Fusarium strains isolated from Norwegian cereals. Mycopathologia 1998, 144, 103–113. [Google Scholar] [CrossRef] [PubMed]
- NRC. Nutrient Requirements of Fish and Shrimp; National Research Council, The National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Harvey, R.B.; Edrington, T.S.; Kubena, L.F.; Elissalde, M.H.; Casper, H.H.; Rottinghaus, G.E.; Turk, J.R. Effects of dietary fumonisin B1-containing culture material, deoxynivalenol-contaminated wheat, or their combination on growing barrows. Am. J. Vet. Res. 1996, 57, 1790–1794. [Google Scholar] [PubMed]
- Harvey, R.B.; Edrington, T.S.; Kubena, L.F.; Elissalde, M.H.; Corrier, D.E.; Rottinghaus, G.E. Effect of aflatoxin and diacetoxyscirpenol in ewe lambs. Bull. Environ. Contam. Toxicol. 1995, 54, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Kubena, L.F.; Edrington, T.S.; Harvey, R.B.; Buckley, S.A.; Phillips, T.D.; Rottinghaus, G.E.; Casper, H.H. Individual and combined effects of fumonisin B1 present in Fusarium moniliforme culture material and T-2 toxin or deoxynivalenol in broiler chicks. Poult. Sci. 1997, 76, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Sauer, D.B.; Seitz, L.M.; Burroughs, R.; Mohr, H.E.; West, J.L.; Milleret, R.J.; Anthony, H.D. Toxicity of Alternaria metabolites found in weathered sorghum grain at harvest. J. Agric. Food Chem. 1978, 26, 1380–1383. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Benford, D.; Boobis, A.; Ceccatelli, S.; Cottrill, B.; Cravedi, J.; di Domenico, A.; Doerge, D.; Dogliotti, E.; Edler, L.; et al. Scientific opinion on the risks for animal and public health related to the presence of Alternaria toxins in feed and food. EFSA J. 2011, 9, 2407–2504. [Google Scholar]
- Rotter, R.G.; Trenholm, H.L.; Prelusky, D.B.; Hartin, K.E.; Thompson, B.K.; Miller, J.D. A preliminary examination of potential interactions between deoxynivalenol (DON) and other selected Fusarium metabolites in growing pigs. Can. J. Anim. Sci. 1992, 72, 107–116. [Google Scholar] [CrossRef]
- Kleinova, M.; Zöllner, P.; Kahlbacher, H.; Hochsteiner, W.; Lindner, W. Metabolic profiles of the mycotoxin zearalenone and of the growth promoter zeranol in urine, liver, and muscle of heifers. J. Agric. Food Chem. 2002, 50, 4769–4776. [Google Scholar] [CrossRef] [PubMed]
- Le Guevel, R.; Pakdel, F. Assessment of oestrogenic potency of chemicals used as growth promoter by in vitro methods. Hum. Reprod. 2001, 16, 1030–1036. [Google Scholar] [CrossRef] [PubMed]
- Swamy, H.V.L.N.; Smith, T.K.; MacDonald, E.J.; Boermans, H.J.; Squires, E.J. Effects of feeding a blend of grains naturally contaminated with Fusarium mycotoxins on swine performance, brain regional neurochemistry, and serum chemistry and the efficacy of a polymeric glucomannan mycotoxin adsorbent. J. Anim. Sci. 2002, 80, 3257–3267. [Google Scholar] [PubMed]
- Rotter, B.A.; Thompson, B.K.; Lessard, M.; Trenholm, H.L.; Tryphonas, H. Influence of low-level exposure to Fusarium mycotoxins on selected immunological and hematological parameters in young swine. Toxicol. Sci. 1994, 23, 117–124. [Google Scholar] [CrossRef]
- Pietsch, C.; Bucheli, T.D.; Wettstein, F.E.; Burkhardt-Holm, P. Frequent biphasic cellular responses of permanent fish cell cultures to deoxynivalenol (DON). Toxicol. Appl. Pharmacol. 2011, 256, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Maul, R.; Warth, B.; Kant, J.-S.; Schebb, N.H.; Krska, R.; Koch, M.; Sulyok, M. Investigation of the hepatic glucuronidation pattern of the Fusarium mycotoxin deoxynivalenol in various species. Chem. Res. Toxicol. 2012, 25, 2715–2717. [Google Scholar] [CrossRef] [PubMed]
- Guan, S.; He, J.; Young, J.C.; Zhu, H.; Li, X.-Z.; Ji, C.; Zhou, T. Transformation of trichothecene mycotoxins by microorganisms from fish digesta. Aquaculture 2009, 290, 290–295. [Google Scholar] [CrossRef]
- Fioramonti, J.; Dupuy, C.; Dupuy, J.; Bueno, L. The mycotoxin, deoxynivalenol, delays gastric emptying through serotonin-3 receptors in rodents. J. Pharmacol. Exp. Ther. 1993, 266, 1255–1260. [Google Scholar] [PubMed]
- Lun, A.K.; Young, L.G.; Lumsden, J.H. The effects of vomitoxin and feed intake on the performance and blood characteristics of young pigs. J. Anim. Sci. 1985, 61, 1178–1185. [Google Scholar] [PubMed]
- Ryerse, I.; Hooft, J.; Bureau, D.; Hayes, M.; Lumsden, J. Purified deoxynivalenol or feed restriction reduces mortality in rainbow trout, Oncorhynchus mykiss (Walbaum), with experimental bacterial coldwater disease but biologically relevant concentrations of deoxynivalenol do not impair the growth of Flavobacterium psychrophilum. J. Fish Dis. 2014. [Google Scholar] [CrossRef]
- Ryerse, I.A.; Hooft, J.M.; Bureau, D.P.; Anthony Hayes, M.; Lumsden, J.S. Diets containing corn naturally contaminated with deoxynivalenol reduces the susceptibility of rainbow trout (Oncorhynchus mykiss) to experimental Flavobacterium psychrophilum infection. Aquac. Res. 2014. [Google Scholar] [CrossRef]
- Matejova, I.; Modra, H.; Blahova, J.; Franc, A.; Fictum, P.; Sevcikova, M.; Svobodova, Z. The effect of mycotoxin deoxynivalenol on haematological and biochemical indicators and histopathological changes in rainbow trout (Oncorhynchus mykiss). BioMed Res. Int. 2014. [Google Scholar] [CrossRef] [PubMed]
- Hinton, D.E.; Lauren, D.J.; Holliday, T.; Giam, C. Liver Structural Alterations Accompanying Chronic Toxicity in Fishes: Potential Biomarkers of Exposure; Preprints of Papers Presented at National Meeting; Division of Water, Air and Waste Chemistry, American Chemical Society: Washington, DC, USA, 1988. [Google Scholar]
- Bernet, D.; Schmidt, H.; Meier, W.; Burkhardt-Holm, P.; Wahli, T. Histopathology in fish: Proposal for a protocol to assess aquatic pollution. J. Fish Dis. 1999, 22, 25–34. [Google Scholar] [CrossRef]
- Rašković, B.S.; Stanković, M.B.; Marković, Z.Z.; Poleksić, V.D. Histological methods in the assessment of different feed effects on liver and intestine of fish. J. Agric. Sci. 2011, 56, 87–100. [Google Scholar]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 6th ed.; AOAC: Arlington, VA, USA, 1995; Volume 1, pp. 16–17. [Google Scholar]
- Malachová, A.; Sulyok, M.; Beltrán, E.; Berthiller, F.; Krska, R. Optimization and validation of a quantitative liquid chromatography-tandem mass spectrometric method covering 295 bacterial and fungal metabolites including all regulated mycotoxins in four model food matrices. J. Chromatogr. A 2014, 1362, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Clark, G. Staining Procedures, 3rd ed.; Williams & Wilkins: Baltimore, MD, USA, 1973; pp. xvii, 418. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tola, S.; Bureau, D.P.; Hooft, J.M.; Beamish, F.W.H.; Sulyok, M.; Krska, R.; Encarnação, P.; Petkam, R. Effects of Wheat Naturally Contaminated with Fusarium Mycotoxins on Growth Performance and Selected Health Indices of Red Tilapia (Oreochromis niloticus × O. mossambicus). Toxins 2015, 7, 1929-1944. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7061929
Tola S, Bureau DP, Hooft JM, Beamish FWH, Sulyok M, Krska R, Encarnação P, Petkam R. Effects of Wheat Naturally Contaminated with Fusarium Mycotoxins on Growth Performance and Selected Health Indices of Red Tilapia (Oreochromis niloticus × O. mossambicus). Toxins. 2015; 7(6):1929-1944. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7061929
Chicago/Turabian StyleTola, Siriporn, Dominique P. Bureau, Jamie M. Hooft, Frederick W. H. Beamish, Michael Sulyok, Rudolf Krska, Pedro Encarnação, and Rakpong Petkam. 2015. "Effects of Wheat Naturally Contaminated with Fusarium Mycotoxins on Growth Performance and Selected Health Indices of Red Tilapia (Oreochromis niloticus × O. mossambicus)" Toxins 7, no. 6: 1929-1944. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7061929