
Reduced Toxicity of Shiga Toxin (Stx) Type 2c in Mice Compared to Stx2d Is Associated with Instability of Stx2c Holotoxin

Abstract
:1. Introduction

2. Results and Discussion
2.1. Results
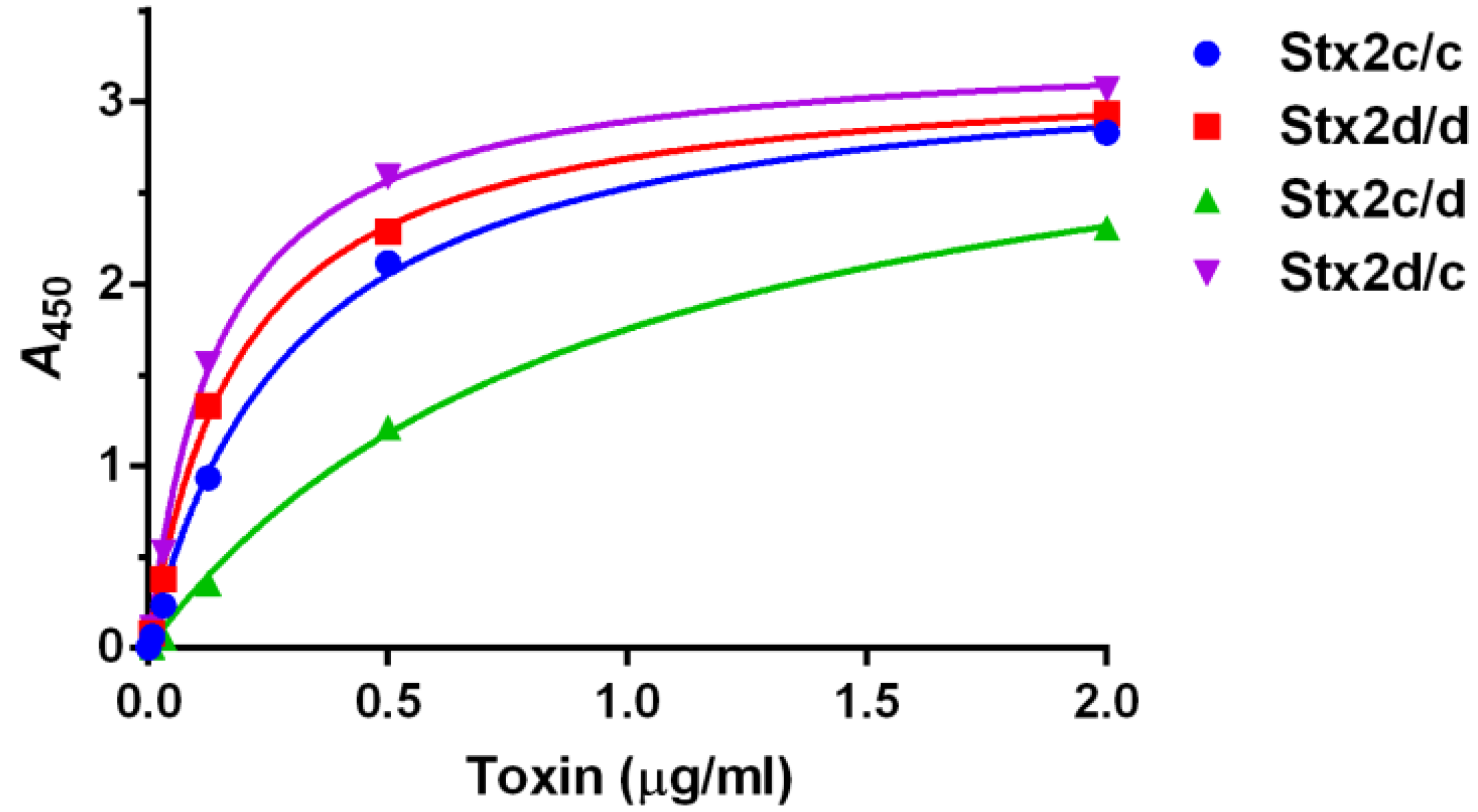
2.1.1. Assessment of Purified Toxins in Vitro and in Vivo

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin | Amino Acids at Position 291 and 297, Respectively | Log Specific Activity, CD50/mg | CD50 95% Confidence Interval | LD50, ng | LD50 95% Confidence Interval, ng |
|---|---|---|---|---|---|
| Stx2c/c | F, K | 7.0 | 6.5–7.5 | 14 | 5.1–38 |
| Stx2d/d | S, E | 7.0 | 5.5–7.0 | 1.8 | 0.94–3.0 |
| Stx2c/d | F, E | 6.3 * | 7.0–7.9 | 24 | 12–49 |
| Stx2d/c | S, K | 7.4 | 6.8–7.1 | 2.5 | 1.5–3.9 |
2.1.2. Binding of Toxins to Purified Gb3

2.1.3. Thermostability of Toxins

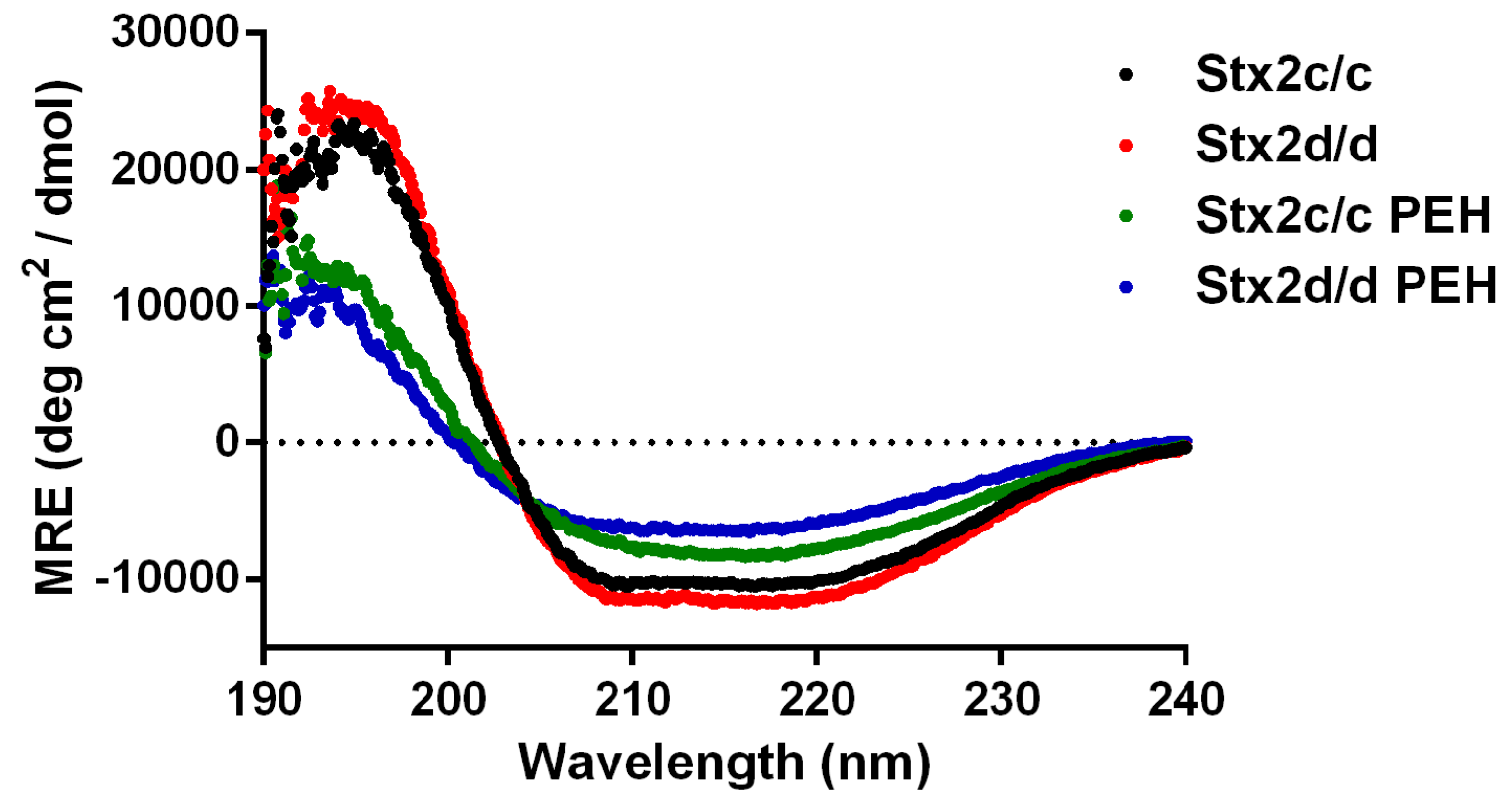
2.1.4. Secondary Structure of Stx2c and Stx2d

| Toxin | Helix, % (SD *) | Strand, % (SD) | Turns, % (SD) | Unordered, % (SD) |
|---|---|---|---|---|
| Stx2c/c | 31.2 (0.9) | 26.3 (1.6) | 10.6 (0.3) | 31.9 (1.0) |
| Stx2d/d | 31.0 (1.0) | 25.3 (1.5) | 20.0 (0.7) | 23.6 (0.8) |
| Stx2c/c PEH | 18.4 (0.5) | 33.4 (1.3) | 11.6 (0.3) | 36.6 (1.0) |
| Stx2d/d PEH | 12.5 (0.4) | 37.9 (2.0) | 11.8 (0.4) | 37.8 (1.2) |
2.2. Discussion

3. Experimental Section
3.1. Toxins and Cell Culture
3.2. Cytotoxicity Assay
3.3. Animal Research Ethics Statement
3.4. Mouse Toxicity Assay
3.5. Thermostability Assay
3.6. Gb3 ELISA
3.7. Far-UV CD Spectroscopy of Stx2c/c and Stx2d/d
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Strockbine, N.A.; Marques, L.R.; Newland, J.W.; Smith, H.W.; Holmes, R.K.; O’Brien, A.D. Two toxin-converting phages from Escherichia coli O157:H7 strain 933 encode antigenically distinct toxins with similar biologic activities. Infect. Immun. 1986, 53, 135–140. [Google Scholar] [PubMed]
- Mead, P.S.; Griffin, P.M. Escherichia coli O157:H7. Lancet 1998, 352, 1207–1212. [Google Scholar] [CrossRef]
- Richardson, S.E.; Karmali, M.A.; Becker, L.E.; Smith, C.R. The histopathology of the hemolytic uremic syndrome associated with verocytotoxin-producing Escherichia coli infections. Hum. Pathol. 1988, 19, 1102–1108. [Google Scholar] [CrossRef]
- Karmali, M.A.; Petric, M.; Lim, C.; Fleming, P.C.; Arbus, G.S.; Lior, H. The association between idiopathic hemolytic uremic syndrome and infection by verotoxin-producing Escherichia coli. J. Infect. Dis. 1985, 151, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Rangel, J.M.; Sparling, P.H.; Crowe, C.; Griffin, P.M.; Swerdlow, D.L. Epidemiology of Escherichia coli O157:H7 outbreaks, United States, 1982–2002. Emerg Infect. Dis. 2005, 11, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.L.; Leibowitz, C.S.; Kurosawa, S.; Stearns-Kurosawa, D.J. Shiga toxins and the pathophysiology of hemolytic uremic syndrome in humans and animals. Toxins 2012, 4, 1261–1287. [Google Scholar] [CrossRef] [PubMed]
- Tarr, P.I.; Gordon, C.A.; Chandler, W.L. Shiga-toxin-producing Escherichia coli and haemolytic uraemic syndrome. Lancet 2005, 365, 1073–1086. [Google Scholar] [CrossRef]
- Scheiring, J.; Andreoli, S.P.; Zimmerhackl, L.B. Treatment and outcome of Shiga-toxin-associated hemolytic uremic syndrome (HUS). Pediatr. Nephrol. 2008, 23, 1749–1760. [Google Scholar] [CrossRef] [PubMed]
- McGannon, C.M.; Fuller, C.A.; Weiss, A.A. Different classes of antibiotics differentially influence Shiga toxin production. Antimicrob. Agents Chemother. 2010, 54, 3790–3798. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; McDaniel, A.D.; Wolf, L.E.; Keusch, G.T.; Waldor, M.K.; Acheson, D.W. Quinolone antibiotics induce Shiga toxin-encoding bacteriophages, toxin production, and death in mice. J. Infect. Dis. 2000, 181, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Idelevich, E.A.; Zhang, W.; Bauwens, A.; Schaumburg, F.; Mellmann, A.; Peters, G.; Karch, H. Effects of antibiotics on Shiga toxin 2 production and bacteriophage induction by epidemic Escherichia coli O104:H4 strain. Antimicrob. Agents Chemother. 2012, 56, 3277–3282. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.S.; Mooney, J.C.; Brandt, J.R.; Staples, A.O.; Jelacic, S.; Boster, D.R.; Watkins, S.L.; Tarr, P.I. Risk factors for the hemolytic uremic syndrome in children infected with Escherichia coli O157:H7: A multivariable analysis. Clin. Infect. Dis.: Off. Publ. Infect. Dis. Soc. Am. 2012, 55, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Fraser, M.E.; Fujinaga, M.; Cherney, M.M.; Melton-Celsa, A.R.; Twiddy, E.M.; O’Brien, A.D.; James, M.N. Structure of Shiga toxin type 2 (Stx2) from Escherichia coli O157:H7. J. Biol. Chem. 2004, 279, 27511–27517. [Google Scholar] [CrossRef] [PubMed]
- Fraser, M.E.; Chernaia, M.M.; Kozlov, Y.V.; James, M.N. Crystal structure of the holotoxin from Shigella dysenteriae at 2.5 a resolution. Nat. Struct. Biol. 1994, 1, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Jacewicz, M.; Clausen, H.; Nudelman, E.; Donohue-Rolfe, A.; Keusch, G.T. Pathogenesis of Shigella diarrhea. Xi. Isolation of a Shigella toxin-binding glycolipid from rabbit jejunum and hela cells and its identification as globotriaosylceramide. J. Exp. Med. 1986, 163, 1391–1404. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, A.A.; Brown, J.E.; Stromberg, N.; Westling-Ryd, M.; Schultz, J.E.; Karlsson, K.A. Identification of the carbohydrate receptor for Shiga toxin produced by Shigella dysenteriae type 1. J. Biol. Chem. 1987, 262, 1779–1785. [Google Scholar] [PubMed]
- Waddell, T.; Head, S.; Petric, M.; Cohen, A.; Lingwood, C. Globotriosyl ceramide is specifically recognized by the Escherichia coli verocytotoxin 2. Biochem. Biophys. Res. Commun. 1988, 152, 674–679. [Google Scholar] [CrossRef]
- Muthing, J.; Meisen, I.; Zhang, W.; Bielaszewska, M.; Mormann, M.; Bauerfeind, R.; Schmidt, M.A.; Friedrich, A.W.; Karch, H. Promiscuous Shiga toxin 2e and its intimate relationship to forssman. Glycobiology 2012, 22, 849–862. [Google Scholar] [CrossRef] [PubMed]
- Melton-Celsa, A.R.; Kokai-Kun, J.F.; O’Brien, A.D. Activation of Shiga toxin type 2d (Stx2d) by elastase involves cleavage of the C-terminal two amino acids of the A2 peptide in the context of the appropriate B pentamer. Mol. Microbiol. 2002, 43, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Bunger, J.C.; Melton-Celsa, A.R.; O’Brien, A.D. Shiga toxin type 2dact displays increased binding to globotriaosylceramide in vitro and increased lethality in mice after activation by elastase. Toxins 2013, 5, 2074–2092. [Google Scholar] [CrossRef] [PubMed]
- Yosief, H.O.; Iyer, S.S.; Weiss, A.A. Binding of pk-trisaccharide analogs of globotriaosylceramide to Shiga toxin variants. Infect. Immun. 2013, 81, 2753–2760. [Google Scholar] [CrossRef] [PubMed]
- Bergan, J.; Dyve Lingelem, A.B.; Simm, R.; Skotland, T.; Sandvig, K. Shiga toxins. Toxicon: Off. J. Int. Soc. Toxinol. 2012, 60, 1085–1107. [Google Scholar] [CrossRef] [PubMed]
- Melton-Celsa, A.R. Shiga toxin (Stx) classification, structure, and function. Microbiol. Spectr. 2014. [Google Scholar] [CrossRef]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Pierard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, A.D.; Tesh, V.L.; Donohue-Rolfe, A.; Jackson, M.P.; Olsnes, S.; Sandvig, K.; Lindberg, A.A.; Keusch, G.T. Shiga toxin: Biochemistry, genetics, mode of action, and role in pathogenesis. Curr. Top. Microbiol. Immunol. 1992, 180, 65–94. [Google Scholar] [PubMed]
- Ethelberg, S.; Olsen, K.E.; Scheutz, F.; Jensen, C.; Schiellerup, P.; Enberg, J.; Petersen, A.M.; Olesen, B.; Gerner-Smidt, P.; Molbak, K. Virulence factors for hemolytic uremic syndrome, denmark. Emerg. Infect. Dis. 2004, 10, 842–847. [Google Scholar] [CrossRef] [PubMed]
- Siegler, R.L.; Obrig, T.G.; Pysher, T.J.; Tesh, V.L.; Denkers, N.D.; Taylor, F.B. Response to Shiga toxin 1 and 2 in a baboon model of hemolytic uremic syndrome. Pediatr. Nephrol. 2003, 18, 92–96. [Google Scholar] [PubMed]
- Persson, S.; Olsen, K.E.; Ethelberg, S.; Scheutz, F. Subtyping method for Escherichia coli Shiga toxin (verocytotoxin) 2 variants and correlations to clinical manifestations. J. Clin. Microbiol. 2007, 45, 2020–2024. [Google Scholar] [CrossRef] [PubMed]
- Lindgren, S.W.; Samuel, J.E.; Schmitt, C.K.; O’Brien, A.D. The specific activities of Shiga-like toxin type II (Slt-II) and Slt-II-related toxins of enterohemorrhagic Escherichia coli differ when measured by vero cell cytotoxicity but not by mouse lethality. Infect. Immun. 1994, 62, 623–631. [Google Scholar] [PubMed]
- Fuller, C.A.; Pellino, C.A.; Flagler, M.J.; Strasser, J.E.; Weiss, A.A. Shiga toxin subtypes display dramatic differences in potency. Infect. Immun. 2011, 79, 1329–1337. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, D.L.; Jackson, M.P.; Perera, L.P.; Holmes, R.K.; O’Brien, A.D. In vivo formation of hybrid toxins comprising Shiga toxin and the Shiga-like toxins and role of the B subunit in localization and cytotoxic activity. Infect. Immun. 1989, 57, 3743–3750. [Google Scholar] [PubMed]
- Russo, L.M.; Melton-Celsa, A.R.; Smith, M.J.; O’Brien, A.D. Comparisons of native Shiga toxins (stxs) type 1 and 2 with chimeric toxins indicate that the source of the binding subunit dictates degree of toxicity. PLoS ONE 2014, 9, e93463. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Friedrich, A.W.; Aldick, T.; Schurk-Bulgrin, R.; Karch, H. Shiga toxin activatable by intestinal mucus in Escherichia coli isolated from humans: Predictor for a severe clinical outcome. Clin. Infect. Dis.: Off. Publ. Infect. Dis. Soc. Am. 2006, 43, 1160–1167. [Google Scholar] [CrossRef] [PubMed]
- Iyoda, S.; Manning, S.D.; Seto, K.; Kimata, K.; Isobe, J.; Etoh, Y.; Ichihara, S.; Migita, Y.; Ogata, K.; Honda, M.; et al. Phylogenetic clades 6 and 8 of enterohemorrhagic Escherichia coli O157:H7 with particular Stx subtypes are more frequently found in isolates from hemolytic uremic syndrome patients than from asymptomatic carriers. Open Forum Infect. Dis. 2014, 1, ofu061. [Google Scholar] [CrossRef] [PubMed]
- National Research Council Committee for the update of the Guide for care and use of laboratory animals. Guide for The Care and Use Of Laboratory Animals, 8th ed.; The National Academies Press: Washington, D.C., USA, 2011. [Google Scholar]
- Wadolkowski, E.A.; Sung, L.M.; Burris, J.A.; Samuel, J.E.; O'Brien, A.D. Acute renal tubular necrosis and death of mice orally infected with Escherichia coli strains that produce Shiga-like toxin type II. Infect. Immun. 1990, 58, 3959–3965. [Google Scholar] [PubMed]
- Perera, L.P.; Marques, L.R.; O'Brien, A.D. Isolation and characterization of monoclonal antibodies to Shiga-like toxin II of enterohemorrhagic Escherichia coli and use of the monoclonal antibodies in a colony enzyme-linked immunosorbent assay. J. Clin. Microbiol. 1988, 26, 2127–2131. [Google Scholar] [PubMed]
- DichroWeb. Available online: http://dichroweb.cryst.bbk.ac.uk/html/home.shtml (accessed on 20 June 2015).
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bunger, J.C.; Melton-Celsa, A.R.; Maynard, E.L.; O'Brien, A.D. Reduced Toxicity of Shiga Toxin (Stx) Type 2c in Mice Compared to Stx2d Is Associated with Instability of Stx2c Holotoxin. Toxins 2015, 7, 2306-2320. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7062306
Bunger JC, Melton-Celsa AR, Maynard EL, O'Brien AD. Reduced Toxicity of Shiga Toxin (Stx) Type 2c in Mice Compared to Stx2d Is Associated with Instability of Stx2c Holotoxin. Toxins. 2015; 7(6):2306-2320. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7062306
Chicago/Turabian StyleBunger, Joshua C., Angela R. Melton-Celsa, Ernest L. Maynard, and Alison D. O'Brien. 2015. "Reduced Toxicity of Shiga Toxin (Stx) Type 2c in Mice Compared to Stx2d Is Associated with Instability of Stx2c Holotoxin" Toxins 7, no. 6: 2306-2320. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7062306