
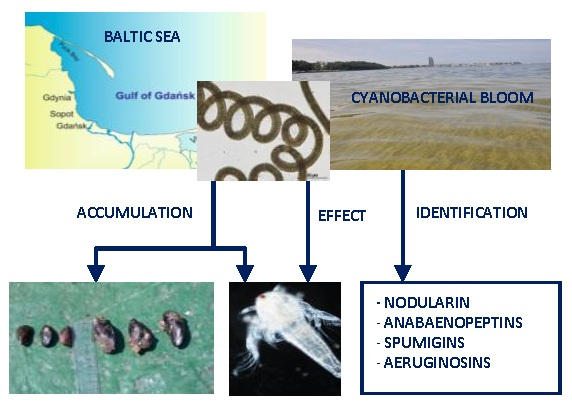
Nodularia spumigena Peptides—Accumulation and Effect on Aquatic Invertebrates

 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Accumulation of Cyanopeptides in Crustaceans (Laboratory Experiment) and in Blue Mussels (Environmental Samples)


{kind=link}
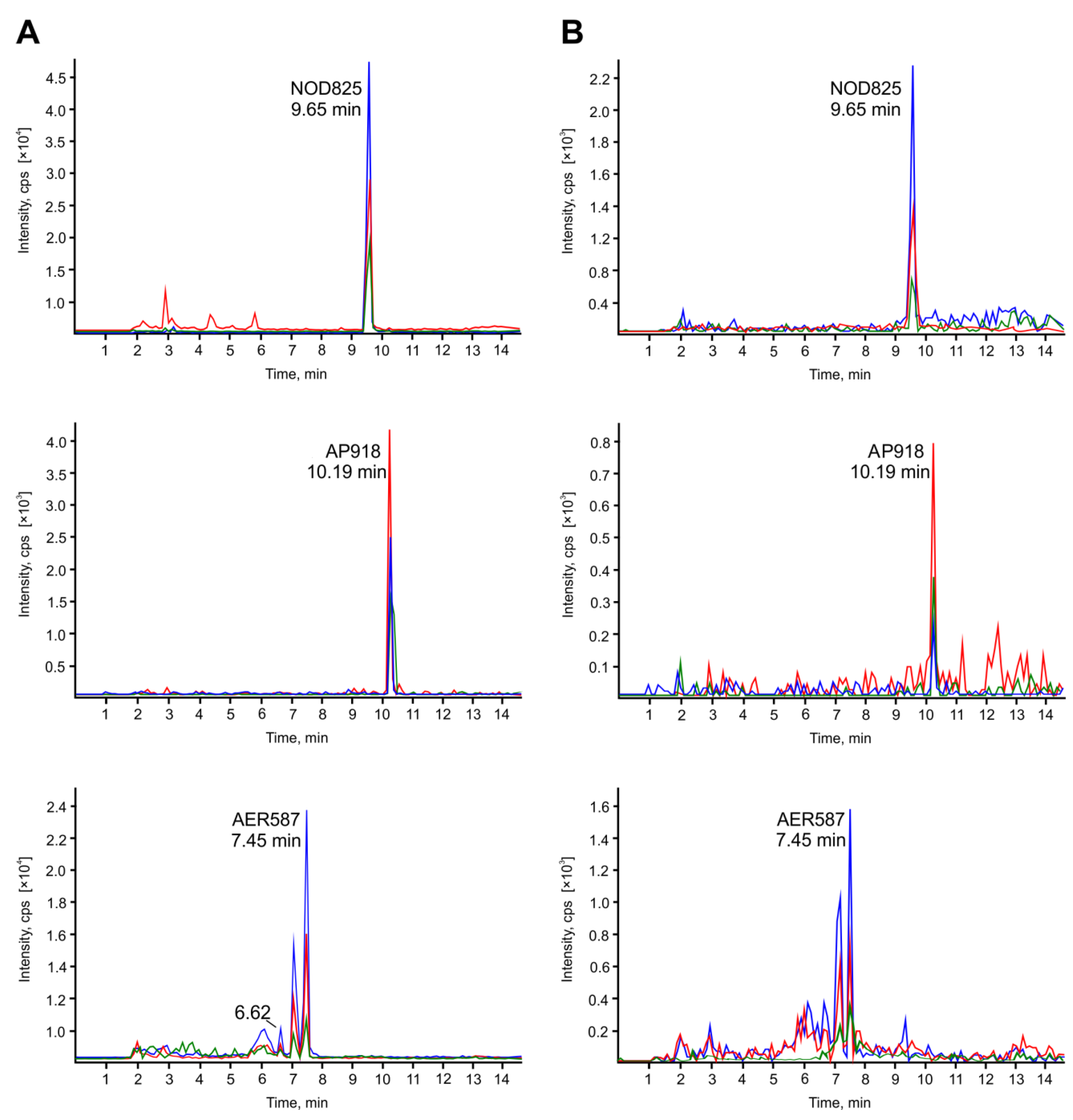{kind=link}
{kind=link}
{kind=link}
| m/z [M+H]+ | Peptide structure | Peak area (counts) | ||||
|---|---|---|---|---|---|---|
| Laboratory experiment | Environmental samples | |||||
| A. franciscana 100 mg d.w. | T. platyurus 100 mg d.w. | Blue mussels 100 mg d.w. 20 August 2012 | Cyanobacteria 100 mg d.w. 27 July 2012 | |||
| L > 3 cm | L < 2 cm | |||||
| Cyclic peptides: anabaenopeptins, nodularin | ||||||
| AP 918 | Phe-CO-[Lys-Val-Hph-MeHty-MetO] | 1.1 × 105 | 0.6 × 105 | 0.5 × 107 | 0.7 × 107 | 0.9 × 107 |
| AP 916 | Phe-CO-[Lys-Val-Hty-MeHty-AcSer] | - | Traces | - | - | 0.3 × 107 |
| AP 902 | Phe-CO-[Lys-Val-Hph-MeHty-Met] | 0.9 × 105 | 0.2 × 105 | - | 0.3 × 107 | 0.4 × 107 |
| AP 900 | Phe-CO-[Lys-Val-Hph-MeHty-AcSer] | 1.3 × 105 | 0.26 × 105 | - | - | 0.5 × 107 |
| AP 884 | Ile-CO-[Lys-Val-Hph-MeHty-MetO] | 3.2 × 105 | 1.8 × 105 | 0.8 × 107 | 0.9 × 107 | 1.2 × 107 |
| AP 868 | Ile-CO-[Lys-MetO-Hty-MeHty-Met] | - | Traces | 1.7 × 107 | 1.9 × 107 | 1.5 × 107 |
| AP 851 (E) | Arg-CO-[Lys-Val-Hty-MeAla-Phe] | 13.5 × 105 | 3.8 × 105 | 1.4 × 107 | 1.5 × 107 | 2.1 × 107 |
| AP 837 (B) | Arg-CO-[Lys-Val-Hty-MeAla-Phe] | - | - | 0.9 × 107 | 1.0 × 107 | 0.5 × 107 |
| AP 828 | Phe-CO-[Lys-Val-Hty-MeAla-Phe] | 22.9 × 105 | 5.9 × 105 | - | 0.9 × 107 | 1.7 × 107 |
| AP 824 | Ile-CO-[Lys-Val-Hph-MeHty-Ser] | - | - | 1.2 × 107 | 1.4 × 107 | 5.6 × 107 |
| AP 808 | Ile-CO-[Lys-Val-Hty-MeAla-Hph ] | - | - | 1.3 × 107 | 1.5 × 107 | 1.5 × 107 |
| NOD 825 | Cyclo[MeAsp-Arg-Adda-Glu-Mdhb] | 16.0 × 106 | 1.1 × 106 | 2.6 × 107 | 2.8 × 107 | 5.7 × 107 |
| [Asp]3NOD811 | Cyclo[Asp-Arg-Adda-Glu-Mdhb] | - | - | 1.3 × 107 | 1.5 × 107 | 3.2 × 107 |
| Linear peptides: spumigins, aeruginosin | ||||||
| SPU 627 | Hpla-Hty-MePro-Arg | - | - | - | - | 0.2 × 107 |
| SPU 613 | Hpla-Hty-MePro-Argol | - | - | - | - | 0.3 × 104 |
| SPU 611 | Hpla-Hty-MePro-Argal | 7.1 × 106 | 0.6 × 105 | - | - | 2.5 × 107 |
| SPU 597 | Hpla-Hty-Pro-Argal | - | - | - | - | 0.2 × 104 |
| AER 587 | HA-Tyr-Choi-Argal | 0.2 × 106 | 0.1 × 106 | - | - | 1.1 × 107 |
2.2. Effect of N. spumigena Spent Medium and Cell Extracts on Crustaceans

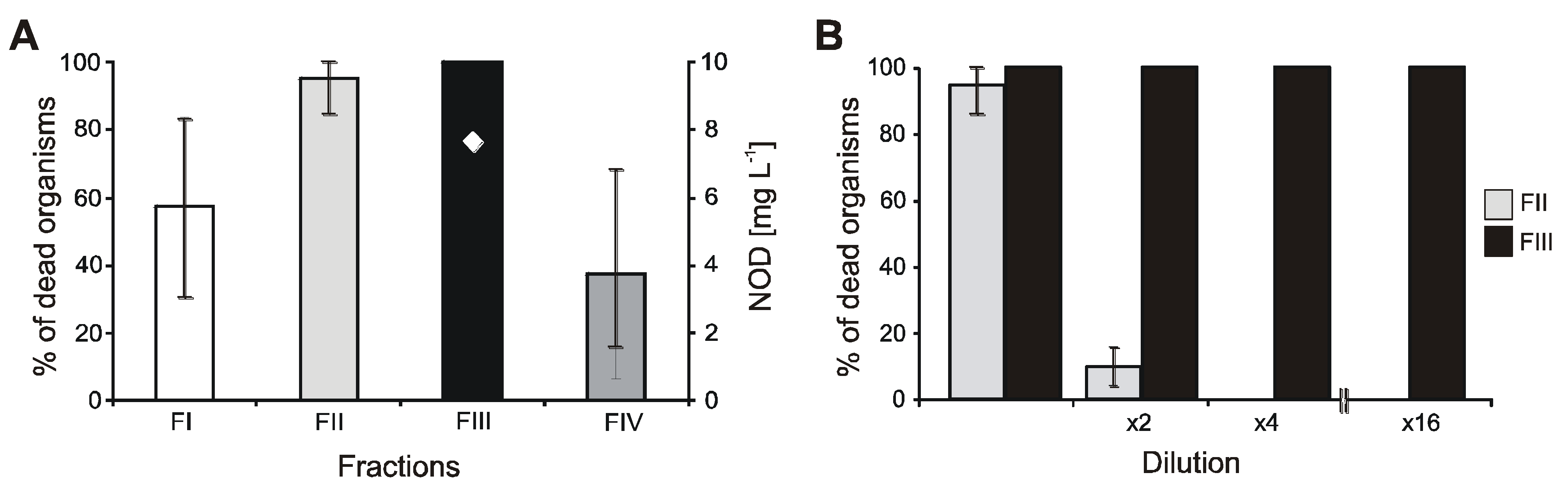
2.3. Effect of Individual Fractions Obtained from N. spumigena Cell Extracts on A. franciscana

| HPLC fraction | I | II | III | IV | ||||
|---|---|---|---|---|---|---|---|---|
| Retention time [min] | 0.0–3.0 | 3.1–7.7 | 7.8–10.0 | 10.1–15.0 | ||||
| Peptides identified in fractions | m/z [M+H]+ | Ion peak area | m/z [M+H]+ | Ion peak area | m/z [M+H]+ | Ion peak area | m/z [M+H]+ | Ion peak area |
| SPU 641 | 2.0 × 107 | SPU 653 | 6.0 × 106 | AP 932 | 5.0 × 106 | AP 842 | 3.0 × 107 | |
| SPU 639 | 2.0 × 107 | SPU 639 | 3.0 × 107 | AP 872 | 6.0 × 106 | AP 828 | 3.0 × 107 | |
| SPU 597 | 3.5 × 107 | SPU 627 | 8.0 × 106 | NOD 825 | 3.2 × 107 | NOD 825 | 1.5 × 106 | |
| SPU 599 | 3.3 × 107 | SPU 613 | 1.1 × 107 | [Asp3]NOD 811 | 2.8 × 107 | |||
| SPU 611 | 1.5 × 107 | |||||||
| SPU 597 | 3.4 × 107 | |||||||
| AER 595 | 1.2 × 107 | |||||||
| AER 589 | 4.0 × 106 | |||||||
| AER 587 | 1.4 × 107 | |||||||
3. Discussion
3.1. Accumulation of Cyanobacteria Metabolites in Aquatic Invertebrates
3.2. The Effect of Cyanobacteria Metabolites on Crustaceans
4. Experimental Section
4.1. Sample Collection and Growth of Cyanobacteria
4.2. Extraction and Fractionation of Cyanobacteria Metabolites
4.3. Biological Tests
4.4. Extraction of Cyanobacterial Peptides from Crustaceans and Mussels
4.5. Chemical Analysis of Cyanobacterial Peptides
4.6. Statistical Analysis
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Paerl, H.W.; Fulton, R.S. Ecology of harmful cyanobacteria. In Ecology of Harmful Algae; Granéli, E., Turner, J., Eds.; Springer: Verlag, Berlin, Germany, 2006; Volume 189, pp. 95–107. [Google Scholar]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Karjalainen, M.; Engstrӧm-Ӧst, J.; Korpinen, S.; Peltonen, H.; Pӓӓkkӧnen, J.P.; Rӧnkkӧnen, S.; Suikkanen, S.; Viitasalo, M. Ecosystem consequences of cyanobacteria in the northern Baltic Sea. Ambio 2007, 36, 195–202. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.S.; Kozlowsky-Suzuki, B. Cyanotoxins: Bioaccumulation and effects on aquatic animals. Mar. Drugs 2011, 9, 2729–2772. [Google Scholar] [CrossRef] [PubMed]
- Marsálek, B.; Blaha, L. Comparison of 17 biotests for detection of cyanobacterial toxicity. Environ. Toxicol. 2004, 19, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Qiu, T.; Xie, P.; Ke, Z.; Li, L.; Guo, L. In situ studies on physiological and biochemical responses of four fishes with different trophic levels to toxic cyanobacterial blooms in a large Chinese lake. Toxicon 2007, 50, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Stal, L.J.; Albertano, P.; Bergman, B.; von Bröockel, K.; Gallon, J.R.; Hayes, P.K.; Sivonen, K.; Walsby, A.E. BASIC: Baltic Sea cyanobacteria. An investigation of the structure and dynamics of water blooms of cyanobacteria in the Baltic Sea—Responses to a changing environment. Cont. Shelf. Res. 2003, 23, 1695–1714. [Google Scholar] [CrossRef]
- Mazur-Marzec, H.; Sutryk, K.; Kobos, J.; Hebel, A.; Hohlfeld, N.; Błaszczyk, A.; Toruńska, A.; Kaczkowska, M.J.; Łysiak-Pastuszak, E.; Kraśniewski, W.; et al. Occurrence of cyanobacteria and cyanotoxin in the Southern Baltic Proper. Filamentous cyanobacteria versus single-cell picocyanobacteria. Hydrobiologia 2013, 701, 235–252. [Google Scholar] [CrossRef]
- Lampert, W. Laboratory studies on zooplankton-cyanobacteria interactions. New Zeal. J. Mar. Fresh. 1987, 21, 483–490. [Google Scholar] [CrossRef]
- Gorokhova, E. Toxic cyanobacteria Nodularia spumigena in the diet of Baltic mysids: Evidence from molecular diet analysis. Harmful Algae 2009, 8, 264–272. [Google Scholar] [CrossRef]
- Hogfors, H.; Motwani, N.H.; Hajdu, S.; El-Shehawy, R.; Holmborn, T.; Vehmaa, A.; Engström-Öst, J.; Brutemark, A.; Gorokhova, E. Bloom-forming cyanobacteria support copepod reproduction and development in the Baltic Sea. PLoS ONE 2014, 9, e112692. [Google Scholar] [CrossRef] [PubMed]
- Karlson, A.M.L.; Duberg, J.; Motwani, N.H.; Hogfors, H.; Klawonn, I.; Ploug, H.; Svedén, J.B.; Garbaras, A.; Sundelin, B.; Hajdu, S.; et al. Nitrogen fixation by cyanobacteria stimulates production in Baltic food webs. AMBIO 2015, 44 (Suppl. 3), S413–S426. [Google Scholar] [CrossRef] [PubMed]
- Engström, J.; Viherluoto, M.; Viitasalo, M. Effects of toxic and non-toxic cyanobacteria on grazing, zooplanktivory and survival of the mysid shrimp Mysis mixta. J. Exp. Mar. Biol. Ecol. 2001, 257, 269–280. [Google Scholar] [CrossRef]
- Engström-Öst, J.; Brutemark, A.; Vehmaa, A.; Motwani, N.H.; Katajisto, T. Consequences of a cyanobacteria bloom for copepod reproduction, mortality and sex ratio. J. Plankton Res. 2015, 37, 388–398. [Google Scholar] [CrossRef]
- Korpinen, S.; Karjalainen, M.; Viitasalo, M. Effects of cyanobacteria on survival and reproduction of the littoral crustacean Gammarus zaddachi (Amphipoda). Hydrobiologia 2006, 559, 285–295. [Google Scholar] [CrossRef]
- Pietsch, C.; Wiegand, C.; Ame, M.V.; Nicklisch, A.; Wunderlin, D.; Pflugmacher, S. The effects of a cyanobacterial crude extract on different aquatic organisms: Evidence for cyanobacterial toxin modulating factors. Environ. Toxicol. 2001, 16, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Nagle, D.G.; Paul, V.J. Production of secondary metabolites by filamentous tropical marine cyanobacteria: ecological functions of the compounds. J. Phycol. 1999, 35, 1412–1421. [Google Scholar] [CrossRef]
- Rohrlack, T.; Christoffersen, K.; Kaebernick, M.; Neilan, B.A. Cyanobacterial protease inhibitor microviridin J causes a lethal molting disruption in Daphnia pulicaria. Appl. Environ. Microbiol. 2004, 70, 5047–5050. [Google Scholar] [CrossRef] [PubMed]
- Rohrlack, T.; Christiansen, G.; Kurmayer, R. Putative antiparasite defensive system involving ribosomal and nonribosomal oligopeptides in cyanobacteria of the genus Planktothrix. Appl. Environ. Microbiol. 2013, 79, 2642–2647. [Google Scholar] [CrossRef] [PubMed]
- Czarnecki, O.; Henning, M.; Lippert, I.; Welker, M. Identification of peptide metabolites of Microcystis (Cyanobacteria) that inhibit trypsin-like activity in planktonic herbivorous Daphnia (Cladocera). Environ. Microbiol. 2006, 8, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Rivera, E.; Paul, V.J. Chemical deterrence of a cyanobacterial metabolite against generalized and specialized grazers. J. Chem. Ecol. 2007, 33, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Baumann, H.I.; Jüttner, F. Inter-annual stability of oligopeptide patterns of Planktothrix rubescens blooms and mass mortality of Daphnia in Lake Hallwilersee. Limnologica 2008, 38, 350–359. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.S.; Soares, M.C.S.; Magalhães, V.F.; Azevedo, S.M.F.O. Biomonitoring of cyanotoxins in two tropical reservoirs by cladoceran toxicity bioassays. Ecotox. Environ. Safe. 2009, 72, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Kankaanpää, H.; Leinio, S.; Olin, M.; Sjovall, O.; Meriluoto, J.; Lehtonen, K.K. Accumulation and depuration of cyanobacterial toxin nodularin and biomarker responses in the mussel Mytilus edulis. Chemosphere 2007, 68, 1210–1217. [Google Scholar] [CrossRef] [PubMed]
- Vuorinen, P.J.; Sipiä, V.O.; Karlsson, K.; Keinänen, M.; Furey, A.; Allis, O.; James, K.; Perttilä, U.; Rimaila-Pärnänen, E.; Meriluoto, J.A.O. Accumulation and effects of nodularin from a single and repeated oral doses of cyanobacterium Nodularia spumigena on flounder (Platichthys flesus L.). Arch. Environ. Contam. Toxicol. 2009, 57, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Dyble, J.; Gossiaux, D.; Landrum, P.; Kashian, D.; Pothoven, S. A kinetic study of accumulation and elimination of microcystin-LR in yellow perch (Perca flavescens) tissue and implications for human fish consumption. Mar. Drugs 2011, 9, 2553–2571. [Google Scholar] [CrossRef] [PubMed]
- Zanchett, G.; Oliveira-Filho, E.C. Cyanobacteria and cyanotoxins: from impacts on aquatic ecosystems and human health to anticarcinogenic effects. Toxins 2013, 5, 1896–1917. [Google Scholar] [CrossRef] [PubMed]
- Shams, S.; Cerasino, L.; Salmaso, N.; Dietrich, D.R. Experimental models of microcystin accumulation in Daphnia magna grazing on Planktothrix rubescens: Implications for water management. Aquat. Toxicol. 2014, 148, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Namikoshi, M.; Rinehart, K.L. Bioactive compounds produced by cyanobacteria. J. Ind. Microbiol. 1996, 17, 373–384. [Google Scholar] [CrossRef]
- Ishida, K.; Okita, Y.; Matsuda, H.; Okino, T.; Murakami, M. Aeruginosins, protease inhibitors from the cyanobacterium Microcystis aeruginosa. Tetrahedron 1999, 55, 10971–10988. [Google Scholar] [CrossRef]
- Murakami, M.; Suzuki, S.; Itou, Y.; Kodani, S.; Ishida, K. New anabaenopeptins, potent carboxypeptidase-A inhibitors from the cyanobacterium Aphanizomenon flos-aquae. J. Nat. Prod. 2000, 83, 1280–1282. [Google Scholar] [CrossRef]
- Welker, M.; von Döhren, H. Cyanobacterial peptides—Nature’s own combinatorial synthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef] [PubMed]
- Chlipala, G.E.; Mo, S.Y.; Orjala, J. Chemodiversity in freshwater and terrestrial cyanobacteria—A source for drug discovery. Curr. Drug Targets 2011, 12, 1654–1673. [Google Scholar] [CrossRef] [PubMed]
- Kehr, J.C.; Picchi, D.G.; Dittmann, E. Natural product biosyntheses in cyanobacteria: a treasure trove of unique enzymes. Beilstein J. Org. Chem. 2011, 7, 1622–1635. [Google Scholar] [CrossRef] [PubMed]
- Mazur-Marzec, H.; Błaszczyk, A.; Felczykowska, A.; Hohlfeld, N.; Kobos, J.; Toruńska-Sitarz, A.; Devi, P.; Montalvão, S.; D’souza, L.; Tammela, P.; et al. Baltic cyanobacteria—A source of biologically active compounds. Eur. J. Phycol. 2015, 50, 343–360. [Google Scholar] [CrossRef]
- Mayorga, P.; Pérez, K.R.; Cruz, S.M.; Cáceres, S. Comparison of bioassays using the anostracean crustaceans Artemia salina and Thamnocephalus platyurus for plant extract toxicity screening. Rev. Bras. Farmacogn. 2010, 20, 897–903. [Google Scholar] [CrossRef]
- Drobniewska, A.; Tarczyńska, M.; Mankiewicz, J.; Jurczak, T.; Zalewski, M. Increase of crustacean sensitivity to purified hepatotoxic cyanobacterial extracts by manipulation of experimental conditions. Environ. Toxicol. 2004, 19, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Töröknë, A.; Vasdinnyei, R.; Asztalos, B.M. A rapid microbiotest for the detection of cyanobacterial toxins. Environ. Toxicol. 2007, 22, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Karjalainen, M.; Reinikainen, M.; Lindvall, F.; Spoof, L.; Meriluoto, J.A.O. Uptake and accumulation of dissolved, radiolabeled nodularin in Baltic Sea zooplankton. Environ. Toxicol. 2003, 18, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.F.; Tsuji, K.; Watanabe, Y.; Harada, K.I.; Suzuki, M. Release of heptapeptide toxin (microcystin) during decomposition process of Microcystis aeruginosa. Nat. Toxins 1992, 1, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Sipiä, V.; Kankaanpӓӓ, H.; Lahti, K.; Carmichael, W.W.; Meriluoto, J. Detection of nodularin in flounders and cod from the Baltic Sea. Environ. Toxicol. 2001, 16, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, K.; Sipiä, V.; Kankaanpää, H.; Meriluoto, J. Mass spectrometric detection of nodularin and desmethylnodularin in mussels and flounders. J. Chromatogr. B. 2003, 784, 243–253. [Google Scholar] [CrossRef]
- Svensen, C.; Strogyloudi, E.; Wexels Riser, C.; Dahlmann, J.; Legrand, C.; Wassmann, P.; Granéli, E.; Pagou, K. Reduction of cyanobacterial toxins through coprophagy in Mytilus edulis. Harmful Algae 2005, 4, 329–336. [Google Scholar] [CrossRef]
- Gkelis, S.; Lanaras, T.; Sivonen, K. The presence of microcystins and other cyanobacterial bioactive peptides in aquatic fauna collected from Greek freshwaters. Aquat. Toxicol. 2006, 78, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Rouhiainen, L.; Jokela, J.; Fewer, D.P.; Urmann, M.; Sivonen, K. Two alternative starter modules for the nonribosomal biosynthesis of specific anabaenopeptin variants in Anabaena (cyanobacteria). Chem. Biol. 2010, 17, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Canuel, E.A.; Lerberg, E.J.; Dickhut, R.M.; Kuehl, S.A.; Bianchi, T.S.; Wakeham, S.G. Changes in sediment and organic carbon accumulation in a highly disturbed ecosystem: the Sacramento-San Joaquin River Delta (California, USA). Mar. Pollut. Bull. 2009, 59, 154–163. [Google Scholar] [CrossRef] [PubMed]
- DeMott, W.R.; Zhang, Q.X.; Carmichael, W.W. Effects of toxic cyanobacteria and purified toxins on the survival and feeding of a copepod and three species of Daphnia. Limnol. Oceanogr. 1991, 36, 1346–1357. [Google Scholar] [CrossRef]
- Holliland, P.B.; Ahlbeck, I.; Westlund, E.; Hansson, S. Ontogenetic and seasonal changes in diel vertical migration amplitude of the calanoid copepods Eurytemora affinis and Acartia spp. in a coastal area of the northern Baltic proper. J. Plankton Res. 2012, 34, 298–307. [Google Scholar] [CrossRef]
- Gustafsson, S.; Hansson, L.A. Development of tolerance against toxic cyanobacteria in Daphnia. Aquat. Ecol. 2004, 38, 37–44. [Google Scholar] [CrossRef]
- Davies, W.R.; Siu, W.H.L.; Jack, R.W.; Wu, R.S.S.; Lam, P.K.S.; Nugegoda, D. Comparative effects of the blue green algae Nodularia spumigena and a lysed extract on detoxification and antioxidant enzymes in the green lipped mussel (Perna viridis). Mar. Pollut. Bull. 2005, 51, 1026–1033. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, P.R.; Reis-Santos, P.; Fonseca, V.; Maia, A.; Ruano, M.; França, S.; Vinagre, C.; Costa, M.J.; Cabral, H. Assessing anthropogenic pressures on estuarine fish nurseries along the Portuguese coast: a multi-index and conceptual approach. Sci. Total Environ. 2007, 374, 199–215. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, M.; Reis, B.; Azevedo, J.; Campos, A.; Osório, H.; Vasconcelos, V.; Martins, J.C. Glutathione transferases responses induced by microcystin-LR in the gills and hepatopancreas of the clam Venerupis philippinarum. Toxins 2015, 7, 2096–2120. [Google Scholar] [CrossRef] [PubMed]
- Keil, C.; Forchert, A.; Fastner, J.; Szewzyk, U.; Rotard, W.; Chorus, I.; Krätke, R. Toxicity and microcystin content of extracts from a Planktothrix bloom and two laboratory strains. Water Res. 2002, 36, 2133–2139. [Google Scholar] [CrossRef]
- Vasconcelos, V.; Oliveira, S.; Teles, F.O. Impact of a toxic and a non-toxic strain of Microcystis aeruginosa on the crayfish Procambarus clarkii. Toxicon 2001, 39, 1461–1470. [Google Scholar] [CrossRef]
- Ersmark, K.; Del Valle, J.R.; Hanessian, S. Chemistry and biology of the aeruginosin family of serine protease inhibitors. Angew. Chem. Int. Ed. 2008, 47, 1202–1223. [Google Scholar] [CrossRef] [PubMed]
- Fewer, D.P.; Jokela, J.; Rouhiainen, L.; Wahlsten, M.; Koskenniemi, K.; Stal, L.J.; Sivonen, K. The non-ribosomal assembly and frequent occurrence of the protease inhibitors spumigins in the bloom-forming cyanobacterium Nodularia spumigena. Mol. Microbiol. 2009, 73, 924–937. [Google Scholar] [CrossRef] [PubMed]
- Sano, T.; Usui, T.; Ueda, K.; Osada, H.; Kaya, K. Isolation of new protein phosphatase inhibitors from two cyanobacteria species, Planktothrix sp. J. Nat. Prod. 2001, 64, 1052–1055. [Google Scholar] [CrossRef] [PubMed]
- Sano, T.; Kaya, K. Oscillamide Y, a chymotrypsin inhibitor from toxic Oscillatoria agardhii. Tetrahedron Lett. 1995, 36, 5933–5936. [Google Scholar] [CrossRef]
- Itou, Y.; Suzuki, S.; Ishida, K.; Murakami, M. Anabaenopeptins G and H, potent carboxypeptidase A inhibitors from the cyanobacterium Oscillatoria agardhii (NIES-595). Bioorg. Med. Chem. Lett. 1999, 9, 1243–1246. [Google Scholar] [CrossRef]
- Agrawal, M.K.; Bagchi, D.; Bagchi, S.N. Acute inhibition of protease and suppression of growth in zooplankter, Moina macrocopa, by Microcystis blooms collected in central India. Hydrobiologia 2001, 464, 37–44. [Google Scholar] [CrossRef]
- Rohrlack, T.; Christoffersen, K.; Friberg-Jensen, U. Frequency of inhibitors of daphnid trypsin in the widely distributed cyanobacterial genus Planktothrix. Environ. Microbiol. 2005, 7, 1667–1669. [Google Scholar] [CrossRef] [PubMed]
- DeMott, W.R.; Dhawale, S. Inhibition of in vitro protein phosphatase activity in three zooplankton species by microcystin-LR, a toxin from cyanobacteria. Arch. Hydrobiol. 1995, 134, 417–424. [Google Scholar]
- Agrawal, M.K.; Bagchi, D.; Bagchi, S.N. Cysteine and serine protease-mediated proteolysis in body homogenate of a zooplankter, Moina macrocopa, is inhibited by the toxic cyanobacterium, Microcystis aeruginosa PCC7806. Comp. Biochem. Physiol. B 2005, 141, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Baumann, H.I.; Jüttner, F. Strong differences in the efficiency of digestive protease inhibitors of the cyanobacterium Planktothrix rubescens. Verh. Internat. Verein. Limnol. 2006, 29, 1849–1853. [Google Scholar]
- Blom, J.F.; Baumann, H.I.; Codd, G.A.; Jüttner, F. Sensitivity and adaptation of aquatic organisms to oscillapeptin J and [d-Asp3, (E)-Dhb7] microcystin-RR. Arch. Hydrobiol. 2006, 167, 547–559. [Google Scholar] [CrossRef]
- Von Elert, E.; Zitt, A.; Schwarzenberger, A. Inducible tolerance to dietary protease inhibitors in Daphnia magna. J. Exp. Biol. 2012, 215, 2051–2059. [Google Scholar] [CrossRef] [PubMed]
- Von Elert, E.; Agrawal, M.K.; Gebauer, C.; Jaensch, H.; Bauer, U.; Zitt, A. Protease activity in guts of Daphnia magna: evidence for trypsin and chymotrypsin enzymes. Comp. Biochem. Physiol. B 2004, 137, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Blom, J.F.; Bister, B.; Bischoff, D.; Nicholson, G.; Jung, G.; Süssmuth, R.D.; Jüttner, F. Oscillapeptin J, a new grazer toxin of the freshwater cyanobacterium Planktothrix rubescens. J. Nat. Prod. 2003, 66, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, C.; Peuthert, A.; Pflugmacher, S.; Carmeli, S. Effects of microcin SF608 and microcystin-LR, two cyanobacterial compounds produced by Microcystis sp., on aquatic organisms. Environ. Toxicol. 2002, 17, 400–406. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazur-Marzec, H.; Sutryk, K.; Hebel, A.; Hohlfeld, N.; Pietrasik, A.; Błaszczyk, A. Nodularia spumigena Peptides—Accumulation and Effect on Aquatic Invertebrates. Toxins 2015, 7, 4404-4420. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7114404
Mazur-Marzec H, Sutryk K, Hebel A, Hohlfeld N, Pietrasik A, Błaszczyk A. Nodularia spumigena Peptides—Accumulation and Effect on Aquatic Invertebrates. Toxins. 2015; 7(11):4404-4420. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7114404
Chicago/Turabian StyleMazur-Marzec, Hanna, Katarzyna Sutryk, Agnieszka Hebel, Natalia Hohlfeld, Anna Pietrasik, and Agata Błaszczyk. 2015. "Nodularia spumigena Peptides—Accumulation and Effect on Aquatic Invertebrates" Toxins 7, no. 11: 4404-4420. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins7114404