White Adipose Tissue as a Site for Islet Transplantation


1
Department of Regenerative Medicine and Transplantation, Faculty of Medicine, Fukuoka University, 7-45-1 Nanakuma, Jonan, Fukuoka 814-0180, Japan
2
Center for Regenerative Medicine, Fukuoka University Hospital, 7-45-1 Nanakuma, Jonan, Fukuoka 814-0180, Japan
*
Author to whom correspondence should be addressed.
Transplantology 2020, 1(2), 55-70; https://0-doi-org.brum.beds.ac.uk/10.3390/transplantology1020006
Submission received: 21 August 2020
/
Revised: 27 August 2020
/
Accepted: 31 August 2020
/
Published: 1 September 2020
(This article belongs to the Special Issue 2021 Profile Papers by Transplantology’s Editorial Board Members)
Abstract
:Although islet transplantation is recognized as a useful cellular replacement therapy for severe diabetes, surgeons face difficulties in islet engraftment. The transplant site is a pivotal factor that influences the engraftment. Although the liver is the current representative site for clinical islet transplantation, it is not the best site because of limitations in immunity, inflammation, and hypoxia. White adipose tissue, including omentum, is recognized as a useful candidate site for islet transplantation. Its effectiveness has been evaluated in not only various basic and translational studies using small and large animals but also in some recent clinical trials. In this review, we attempt to shed light on the characteristics and usefulness of white adipose tissue as a transplant site for islets.
1. Introduction
Islet transplantation (ITx) is considered as a promising and reliable cellular replacement therapy for severe diabetes mellitus (DM) patients with unstable condition of blood glucose (BG) level despite intensive insulin therapy, especially for insulin-dependent type 1 DM patients. The therapeutic outcomes of ITx have gradually, but dramatically, improved through innovations in technology regarding islet isolation, transplantation procedures, and immunosuppressants [1]. The most recent report from the Collaborative Islet Transplant Registry indicated that the insulin-independence (no necessity to use daily insulin injections) rate at 3 years after ITx was 44% [2], and that approximately 80% of the recipients who received 600,000 or more total islet equivalents (IEQs) achieved insulin independence [3]. Furthermore, a phase III study for elucidating the therapeutic effects of clinical ITx in type 1 DM (CIT-07) performed at eight centers in the United States until 2017 revealed that 87.5% and 71% of the diabetic participants achieved an HbA1c level of less than 7.0% and prevention of severe hypoglycemic events at 1 and 2 years after the first ITx, respectively [4]. Although ITx is recognized as a useful therapy that enables an appropriate physiological supply of insulin responding to the changes of blood glucose levels and prevents severe hypoglycemia and life-threatening complications related to micro- and macroangiopathy, including cardiomyopathy, nephropathy, retinopathy, and neuropathy, it still involves some problems that compromise the therapeutic effects.
One of the problems surgeons face is the unsatisfactory transplant efficacy, which depends on the difficulty in engraftment of transplanted islets. Many transplanted islets fail to engraft in a couple days after transplantation [5] because they suffer from harsh environmental factors of immunity [6], inflammation triggered by the innate immune system [7], and ischemia [5], which are affected by the transplant site. For instance, the liver is a major clinical transplant site for islets. However, the liver is not the best site in terms of immunity (owing to liver-resident macrophage (Kupffer cells) and natural killer cells [8,9,10]), inflammation (instant blood-mediated inflammatory reaction [IBMIR], an acute thrombotic and inflammatory reaction that causes damage to transplanted islets [11]), and hypoxia (owing to embolization of the peripheral portal vein by the transplanted islets themselves [5,12]). To date, various organs, including the renal subcapsular space [13], gastrointestinal tract [14], bone marrow [15], spleen [6,16], and muscle and subcutaneous tissue [17], have been examined to assess their characteristics as alternative sites for islets in an effort to establish the ideal transplant site (Table 1). Though they offer various attractive advantages, all of these sites have also limitations, which become obstacles for use in the clinical setting [6,17,18].
In this review, we attempted to shed light on the characteristics and usefulness of white adipose tissue as a transplant site for islets. White adipose tissue, such as omentum (i.e., omental pouch) and mesentery, has the advantages of a high capacity for accommodating a high volume of islets and of belonging to the portal venous system, which provides physiological insulin delivery (Table 1), and is considered a useful candidate as a transplant site of islets. It is believed that these characteristics might be suitable for the clinical setting, especially when the liver cannot be used owing to portal thrombosis and hypertension.
2. Previous Trials Using White Adipose Tissue as a Transplant Site for Islet Transplantation
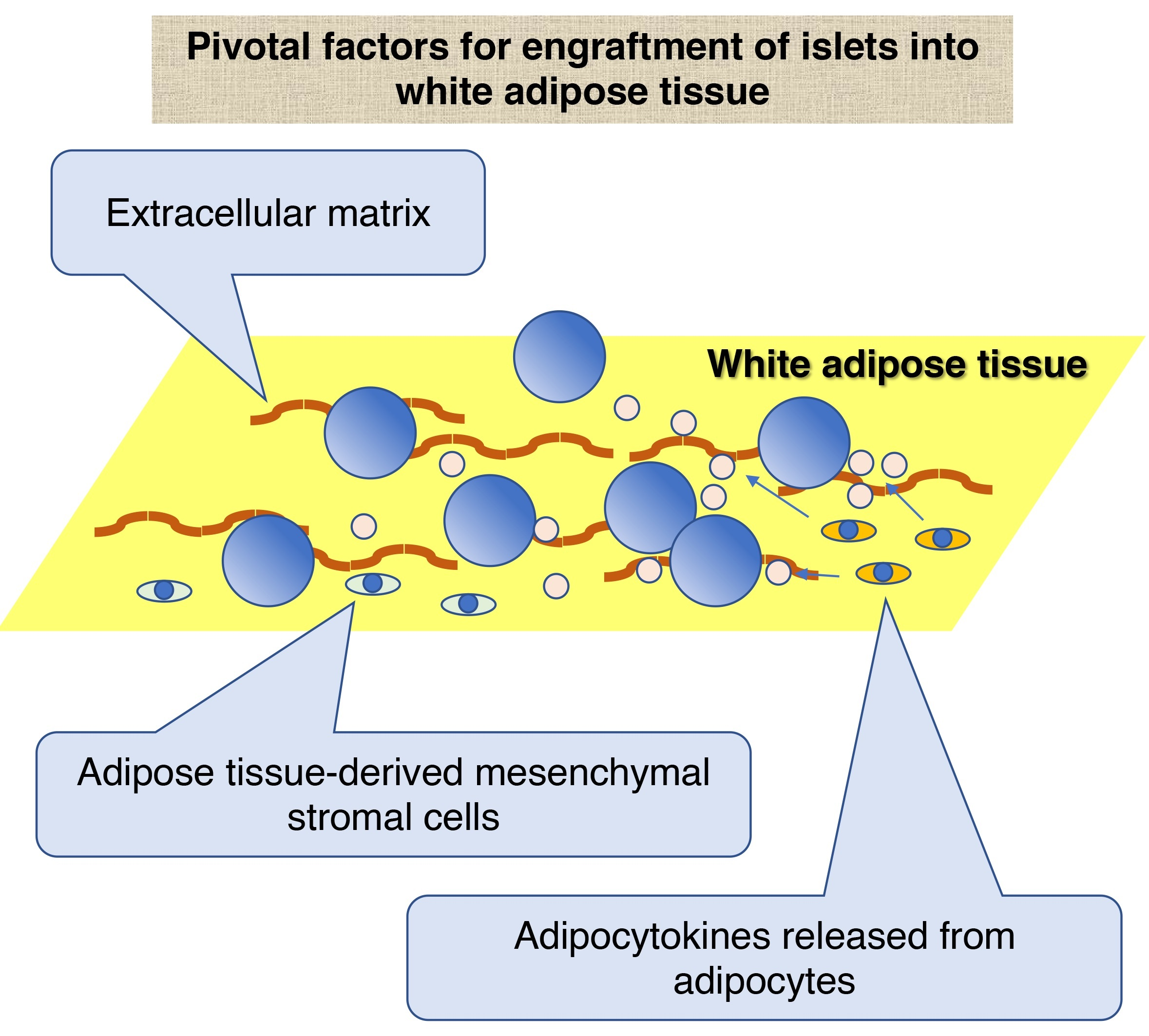
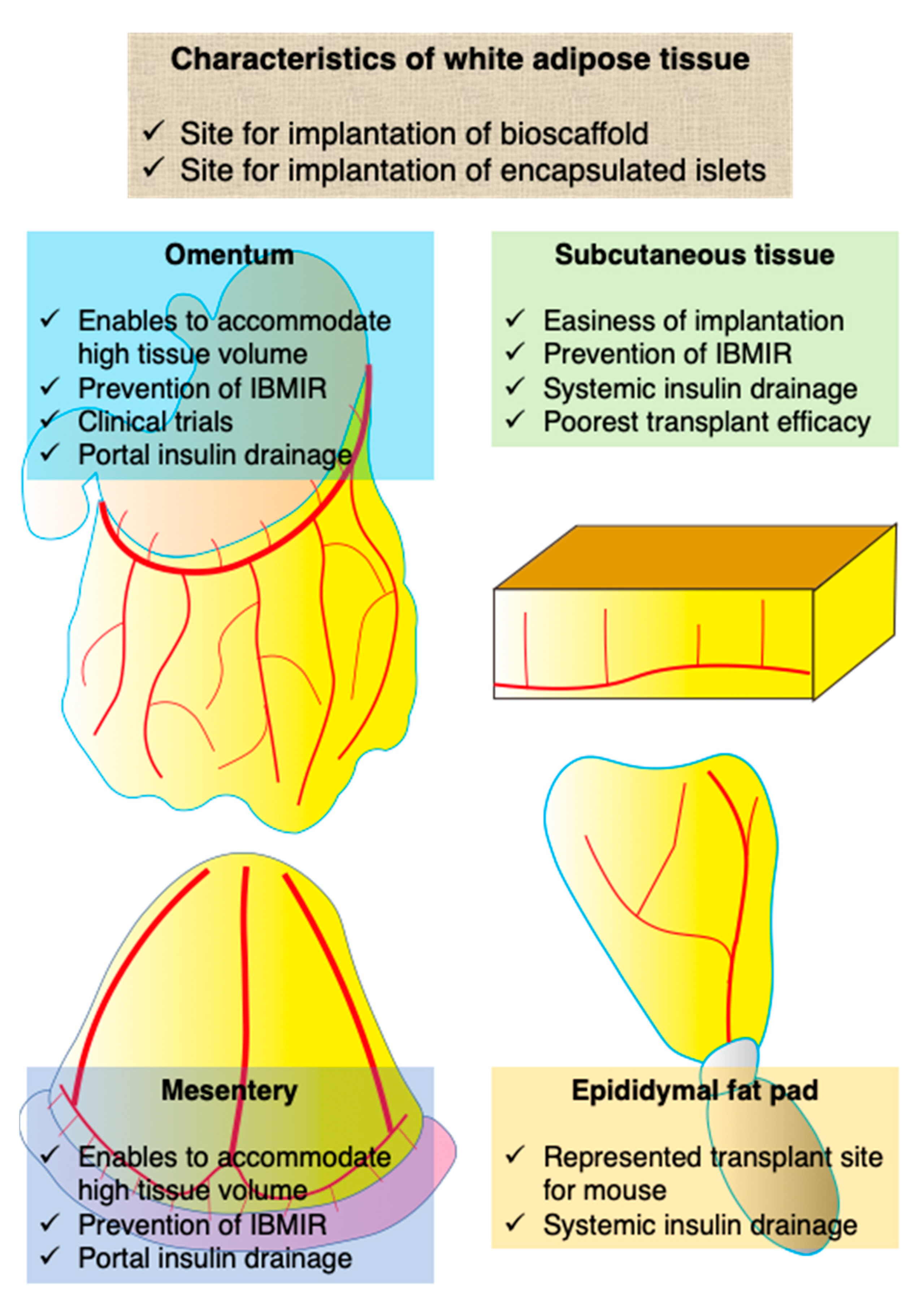
White adipose tissue is a connective tissue that contains mainly lipids and many small vessels. It is found in subcutaneous tissue, abdominal cavity (e.g., omentum and mesentery), bone marrow, muscle, and the breast. Of note, it is believed that white adipose tissue has mainly two advantages as a transplant site for ITx as follows: its spatial capacity and prevention of direct contact between islets and blood. The former is a preferable characteristic for ITx, which enables the accommodation of high tissue volume. The latter contributes to the prevention of IBMIR, which impairs the engraftment of transplanted islets and leads to complications, including bleeding and thrombosis. Because of these characteristics, white adipose tissue has been used as the site for implantation of bioscaffolds, which support the engraftment of islets or encapsulated islets that cannot be infused via vessels. The encapsulation technique is used to protect transplanted islets from the recipient’s immune system (Figure 1).
The first ITx study using white adipose tissue was published in the early 1980s. Outzen and Leiter performed syngeneic and allogeneic rodent ITx into the mammary fat pad and assessed its therapeutic effect and engraftment [19]. To date, various studies have been performed using mainly the three following kinds of white adipose tissues: omentum, mesentery, and epididymal fat pad.
Omentum has been well examined as a candidate islet transplant site. The greater omentum is a large visceral peritoneum that extends from the greater curvature of the stomach, passing in front of the small intestines, and doubling back to ascend to the transverse colon before reaching the posterior abdominal wall. The greater omentum plays a key role in not only peritoneal immunity but also storage of adipose tissue. The omental pouch is a major target of islet transplant site in the omentum. The omental pouch is defined as a space consisting of the greater omentum, which enables the transplantation of a high volume of cells or tissues. Another merit of the omentum is in the physiological drainage route of insulin, because the vascular system of the greater omentum belongs to the portal system, as mentioned in the Introduction (Table 1, Figure 1) [20]. The usefulness of the omentum as an islet transplant site was first revealed in 1983 [21]. Since then, various groups have examined its effectiveness in clinical studies (Table 2) [22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39]. For example, Tuch et al. transplanted allogeneic fetal pancreatic tissues into the omental pouch and muscle of a 29-year-old insulin-dependent diabetic woman in the mid-1980s. Although this was the first clinical case of allogeneic islet transplantation into the omental pouch, there was no engraftment of β cells and no detection of plasma C-peptide at 13 months after transplantation, despite the use of immunosuppressants [40]. In early studies of the omentum, its usefulness was assessed using large animals such as dogs and monkeys [22,34,35,38]. Regarding rodents, rats are mainly used for basic studies. Although the mouse is a major animal model in general, the greater omentum of mice is too small to use for ITx studies [41]. Instead, the epididymal fat pad is popular for ITx studies using mouse models. The focus of recent studies using omentum is to assess the usefulness of bioscaffold, which supports islet engraftment and encapsulated islets that cannot be infused intravenously. In particular, bioscaffolds are used to induce prevascularization in the omentum, which protects transplanted islets from hypoxia resulting from ischemia. Kriz et al. developed a vascularized pocket using the omental pouch by preimplantation of a polymer spacer. They succeeded in achieving normoglycemia of diabetic rats by transplantation of syngeneic islets into the prevascularized pocket [30,39]. Pedraza et al. evaluated the characteristics of a microporous bioscaffold by implantation with islets into the omental pouch of diabetic rats [29]. As in other trials, growth factors such as vascular endothelial growth factor [26,27,39] and angiogenesis-promoting cells such as mesenchymal stromal cells (MSCs) [27] and endothelial cells [31] were used for vascularization of transplanted islets into the omentum. Regarding encapsulated islets, agarose gel (Kobayashi et al.), alginate gel (Pareta et al.), and alginate gel (Ibarra et al.) were assessed to determine the therapeutic effect of implantation into the omental pouch [25,28,36]. The encapsulation technique is aimed at rendering protection, including islets in biomaterials, from the immune response of the recipient.
The advantage of the mesentery as a transplant site for islets is its large space and physiological insulin delivery, much like the omentum. The mesentery belongs to the portal venous system, which contributes to achieving physiological insulin delivery, and it is able to accommodate high volumes of islets (Figure 1). On the other hand, it presents challenges in terms of graft removal without sacrificing the intestinal tract [42]. To the best of our knowledge, there have been few studies that have assessed the characteristics and usefulness of the mesentery as an islet transplant site. The groups led by Rajotte and Michalska demonstrated the usefulness of the mesentery in the 1990s and early 2000s [43,44]. Later, Weaver et al. evaluated the islet transplant efficacy of various white adipose tissues, including the subcutaneous tissue, mesentery, and epididymal fat pad. They showed that the normoglycemia rate of islet-transplanted mice (600 IEQs) into the mesentery was superior to that into subcutaneous tissue but inferior to that into the epididymal fat pad. Additionally, some studies regarding implantation with bioscaffolds for immune isolation [45] and promotion of vascularization [46,47,48] have been reported.
The epididymal fat pad is also used as a transplant site ITx. It is defined as white adipose tissue in the perigonadal region, is considered the major visceral white adipose tissue in rodents [41], and is recognized as one of the largest visceral white adipose tissues, whereas the greater omentum of rodents, especially that of mice, is too small, as previously described. Although the epididymal fat pad has the disadvantage of systemic insulin release, it has been used for the experimental ITx studies instead of the omentum because of its ease in operability (Figure 1). Many studies have shown its usefulness in assessing various types of bioscaffold, including elastin [49], fibrin [50], poly(lactide-co-glycolide) [51,52,53,54], polyethylene glycol [13,42,55,56,57], alginate (ALG) [58,59], and poly-l-lysine [60], much like the omentum (Table 3).
Unlike in rodents, dogs, and monkeys, subcutaneous tissue in humans is composed of connective tissue and white adipose tissue. In other words, subcutaneous tissue can be defined as one of the white adipose tissues in humans. Although subcutaneous tissue has the merit of easiness and safety with minimum invasion of the transplant procedure and monitoring the condition of islet graft, it harbors the poorest transplant efficacy owing to hypoxia, hypovascularity, and immunity (Figure 1) [17]. Conversely, if hypoxic and hypovascular condition can be removed (e.g., prevascularization and hyperbaric oxygenation [68]), subcutaneous tissue can be a promising site for islets. We previously reviewed subcutaneous islet transplantation [17] and thus, show the recent topics about this area in this review. Recently, subcutaneous white adipose tissue in the inguinal area was assessed in a new trial of ITx using white adipose tissue. In general, subcutaneous tissue requires prevascularization for success in ITx because of its hypovascular environment. For example, Forster et al. performed prevascularization of subcutaneous white adipose tissue in the inguinal area using a silicone spacer with Matrigel® and a growth factor and then evaluated the effect of ITx into the spacer [69]. Yasunami et al. performed ITx into the subcutaneous white adipose tissue in the inguinal area without prevascularization and revealed that the transplant efficacy was superior to that of intrahepatic ITx. Furthermore, they also successfully performed allogeneic ITx with 4 months of normoglycemia under immunosuppressants using this site [70]. They showed that it is possible to use subcutaneous tissue as a transplant site for islets with promising therapeutic effects by selecting a preferable site.
The transplant efficacy of ITx using white adipose tissue is unclear. However, some groups have successfully achieved normoglycemia using the epididymal fat pad by one donor [49,53,56] and others by more than two donors [42,50,52,55,62,63,64,65,67]. In our experience, 200 islets can be obtained from a single mouse. It is estimated that the transplant efficacy of the epididymal fat pad might be inferior to that of the kidney, but superior to that of the liver. Furthermore, the transplant efficacy might depend on the white adipose tissue. Weaver et al. compared the transplant efficacy of the epididymal fat pad, mesentery, and subcutaneous white adipose tissue and revealed that the epididymal fat pad was the best transplant site among the white adipose tissues [42]. On the other hand, other groups have shown that the transplant efficacy using the omentum was similar to that of the epididymal fat pad [32].
3. Characteristics of White Adipose Tissue as a Site for Islet Transplantation
Although previous studies have revealed the usefulness of white adipose tissue as a transplant site for ITx, the mechanism that enables the engraftment of the transplanted islets in the white adipose tissue has not been fully discussed. We discuss the mechanism from the views of extracellular matrix (ECM) and cellular components of white adipose tissue.
3.1. ECM of White Adipose Tissue
White adipose tissue is a connective tissue composed of various ECMs, including collagen types I, IV, V, and VI, laminin, and fibronectin [71,72,73]. It is believed that these ECMs support the adhesion of transplanted islets into the white adipose tissue via integrin.
Integrin is one of the adhesion factors expressed on the surface of cellular membranes [74]. It is a heterodimer composed of a combination of 18 types of α subunits (α1–11, E, V, L, M, X, D, and IIb) and 8 types of β subunits (β1–8) that works as a receptor for ECMs. For instance, integrins α1β1, α2β1, α10β1, and α11β1 work as primary receptors for collagens [75]. Integrin α3β1 also mediates attachment to collagen [76]. Regarding other ECMs, integrins α1β1, α2β1, α3β1, α6β1, α7β1, and α6β4 are the receptors for laminin [77], and integrins α3β1, α4β1, α5β1, α8β1, α9β1, αvβ1, αIIbβ3, αvβ3, αvβ6, and α4β7 are receptors for fibronectin [78,79]. On the other hand, it has been shown that islets harbor at least five integrin heterodimers, including α1β1, α3β1, α5β1, αvβ1, and α6β1 [80]. In other words, islets can attach to white adipose tissue via a combination of collagen, laminin, fibronectin, and integrin, which may contribute to the engraftment of islets to white adipose tissue. Furthermore, these correlations between ECMs and adhesion molecules may also support the improvement of the function of engrafted islets. Kaido et al. revealed that the insulin-releasing function of cultured human islet β cells was improved by attachment to collagen IV via integrin α1β1 expressed on the β cells [81]. Bosco et al. also showed that a combination of laminin and integrin α6β1 enhanced the insulin-releasing function of rat islets in glucose-stimulated insulin secretion [82]. Furthermore, Wang’s group clarified the importance of integrin β1 in islets in the development of fetal pancreas and prevention of islet-cell apoptosis via activation of the FAK, MAPK, and ERK signaling pathways [83,84,85]. They later showed integrin α3-regulated islet-cell survival and function via the PI3K and Akt signaling pathway [86].
Recently, we showed that the transplant efficacy of “fat-covered” ITx using epididymal white adipose tissue was superior to that of intraperitoneal ITx and nearly equal to that of renal subcapsular ITx in a rodent model. This “fat-covered” transplantation method is very simple. The epididymal white adipose tissue was mobilized outside of the peritoneal cavity and distended under general anesthesia. Islets were dropped onto the white adipose tissue using a micropipette and covered. Suturing of the adipose tissue was not performed and bioscaffolds and biobinding agents (i.e., ECM) for binding islets to the white adipose tissue were not used. In this study, we showed that fibronectin was expressed in the epididymal adipose tissue and that islets expressed integrin β1, a receptor of fibronectin. It was estimated that transplanted islets were attached to epididymal white adipose tissue via a combination of fibronectin and integrin β1 and, therefore, engraftment of the islets was promoted (Sakata et al., Transplantation 2020, doi:10.1097/TP.0000000000003400; in press). However, the role of adhesion factors in the engraftment of islets into white adipose tissue has not been fully discussed. However, further studies are necessary.
3.2. Cellular Components of White Adipose Tissue
White adipose tissue consists of various cellular components, including adipocytes, adipose tissue-derived mesenchymal stromal (or stem) cells (ADMSCs), vascular endothelial cells, pericytes, macrophages, and different types of white blood cells. Among them, ADMSCs and adipocytes are the main cellular components of white adipose tissue that might contribute to the engraftment of islets.
3.2.1. ADMSCs
Although ADMSCs represent only 5% of the cellular components of white adipose tissue, they play important roles in tissue repair and immune modulation, as do other MSCs in other organs [87,88]. ADMSCs have received attention as cellular candidates for co-transplantation with islets, and many groups have examined their therapeutic effects. The functions of cotransplanted ADMSCs, which might promote the engraftment of islets, are classified into the following three types: neovascularization, reduction of inflammation, and regulation of immunity. At first, ADMSCs promote the formation of a neovascular network between the transplanted islets and recipient by secretion of various growth factors, including vascular endothelial growth factor [89,90], hepatocyte growth factor [89], and transforming growth factor-β [91]. For instance, Ren et al. showed that co-transplantation of ADMSCs increased islet revascularization by induction of hepatocyte growth factor and angiopoietin-1 expressions [92]. Furthermore, ADMSCs reduce proinflammatory cytokines such as tumor necrosis factor (TNF)-α [93,94], interferon-γ [91], interleukin (IL)-6β [93], and IL-17 [91]. Regarding immunomodulation, ADMSCs support immunotolerance in the transplant site by hindering the infiltration of CD4+ and CD8+ T cells [95] and macrophages [94], as well as by promoting the production and infiltration of Tregs [96]. A recent study revealed that xenogeneic co-transplantation of human ADMSCs with neonatal porcine islets led to earlier achievement normoglycemia in diabetic mice compared with transplantation of porcine islets only [97]. These functions of ADMSCs are characterized as supportive effects for promoting the engraftment of transplanted islets. On the other hand, transplanted ADMSCs can ameliorate the diabetic condition by enhancing the proliferation of transplanted islet cells [98], by differentiation into insulin-producing cells themselves [99,100], and by promoting insulin-releasing function of cotransplanted islets [91,93].
Although many studies on the therapeutic effect of ADMSC transplantation have been published, it is still unclear whether resident ADMSCs support the engraftment of transplanted islets into white adipose tissue because of the difficulty of proving their therapeutic effect with the current experimental models. Furthermore, the population of ADMSCs in white adipose tissue is too small, whereas adipocytes represent approximately 90% of white adipose tissue [101]. That resident ADMSCs contributing to the engraftment of transplanted islets offer no doubt. However, their therapeutic effect might be limited because of the smaller population of ADMSCs in white adipose tissue.
3.2.2. Adipocytes
As described previously, adipocytes are the major components of white adipose tissue. They are histologically seen as the cells that contain a large lipid droplet in their cytoplasm. Adipocytes play an important role in lipid regulation and cytokine (“adipocytokines”) release [102]. Regarding lipid regulation, adipocytes store free fatty acids (FFAs) and esterify into triglycerides in energy excess and turn triglycerides into FFAs by using contained enzymes, including adipose triglyceride lipase in a decrease of energy. FFAs work for insulin resistance. Recently, adipocytes became known as the cells producing various adipocytokines. For example, the expression of TNF-α, which induces inflammation and apoptosis, was seen in the white adipose tissue of obese mice [103]. Production of TNF-α in adipocytes is increased in type 2 diabetes [103,104]. Resistin and IL-6 are also produced in white adipose tissue and cause systemic inflammation and induce insulin resistance, similarly to TNF-α and FFA [105]. Monocyte chemoattractant protein-1 (MCP-1) is one of the adipocytokines that induces inflammatory reaction and insulin resistance via infiltration of macrophages in obesity [106,107,108]. It is not clear whether these adipocytokines promote islet engraftment. However, it is well known that TNF-α induced in the peritransplant period damages transplanted islets [7]. Furthermore, previous studies have shown that MCP-1 plays a pivotal role as an inflammatory cytokine in the early graft loss of islets [109,110]. On the other hand, it is difficult to evaluate the effect of IL-6 on transplanted islets. Min et al. revealed both beneficial and harmful effects of IL-6 blockade in ITx. Although blockade of IL-6 reduced innate inflammation after ITx, it inhibited revascularization of transplanted islets [111].
The usefulness for ITx has been shown in two adipocytokines as follows: leptin and adiponectin. Leptin is one of the representative adipocytokines that controls body weight by regulating food intake [112]. Regarding the therapeutic effect in ITx, Denroche et al. demonstrated that administration of low-dose leptin improved the transplant efficacy of ITx using a rodent diabetic model [113]. Lee et al. showed that leptin might enhance the therapeutic effect of ITx by reducing lipotoxicity, which impaired engraftment of intraportal transplanted islets in obese rats’ abnormal leptin signaling [114]. On the other hand, adiponectin has anti-diabetic effects; it contributes to increased insulin sensitivity in target organs like muscle, liver, and endothelial cells [115] and plays anti-inflammatory and anti-atherogenic roles [116,117]. Therefore, adiponectin is a protective factor against obesity, type 2 diabetes, and cardiovascular diseases [118]. It is believed that the beneficial effects of adiponectin depend on its anti-inflammatory and angiogenic activities. Regarding its anti-inflammatory activity, Du et al. showed the therapeutic effect of adiponectin for transplanted islets by prevention of ischemic and reperfusion injury induced by activation of TNF-α-induced nuclear transcription factor-κB pathways [119]. Regarding its angiogenic activity, we showed that (1) vascularization of engrafted islets into white adipose tissue was prominent compared with engrafted islets in the renal subcapsular space and that (2) expressions of adhesion (Fn1, Itgb1, Itgb2) and angiogenic (Vegfa-c) factors in islets were enhanced by incubation of adiponectin (Sakata et al., Transplantation 2020, DOI: 10.1097/TP.0000000000003400; In press). These roles of adiponectin might contribute to the engraftment of islets into white adipose tissue.
3.2.3. Adipose Tissue Macrophage
Macrophage is characterized as an immune cell, which plays roles of phagocytosis and destruction of harmful organisms including bacteria, presents antigens to T cells and induces inflammation via releasing cytokines. Regarding islet transplantation, Kupffer cells, liver tissue macrophages, contribute to prevention of islet engraftment. Kupfer cells are activated during IBMIR, which is occurred through complemental pathway in intraportal transplanted islets and attack to them by phagocytosis and secretion of inflammatory cytokines and free radicals [120]. On the other hand, the roles of adipose tissue macrophage against transplanted islets in white adipose tissue have not been fully discussed. Recently, Russo reviewed about the function of adipose tissue macrophages using lean and obese animal models. Adipose tissue macrophages are classified into to subtypes: tissue resident and monocyte-derived macrophages. While the population of adipose tissue macrophage is 10% of all cells in lean mice, they are increased to over 50% in obese mice [121,122]. It is considered that the 10% macrophages are tissue resident macrophages and increased macrophages are monocyte-derived macrophages. The tissue resident macrophages play as “M2” anti-inflammatory macrophages, which contribute to attenuation of inflammation by anti-inflammatory cytokines and regulation of immune cells in lean adipose tissue. On the other hand, in obese adipose tissue, monocyte-derived macrophages, which are characterized as “M1” macrophages, are increased and induce inflammation via activation of T cells and production of pro-inflammatory cytokines [123]. It is considered that the balance between tissue resident and monocyte-derived macrophages might influence of islet engraftment in white adipose tissue. However, further studies are necessary.
4. Clinical Trial of Islet Transplantation into White Adipose Tissue and Conclusion
Recent progress in clinical trials of islet transplantation using the omentum is seen in total pancreatectomy with islet autotransplantation. Stice et al. performed total pancreatectomy with islet autotransplantation into the omental pouch in chronic pancreatitis patients who could not receive complete intraportal infusion of islets due to portal hypertension and showed a similar glycemic control to “normal” intraportal islet autotransplantation [124]. Regarding allogeneic ITx, a phase I and II clinical trial for allogeneic islet cells transplanted onto the omentum, conducted by the University of Miami beginning in 2014 (ClinicalTrials.gov: NCT02213003), is ongoing. This trial aims to elucidate the therapeutic outcome of clinical ITx onto the omentum. The primary endpoint is achieving at least 6.5% HbA1c and no severe hypoglycemia by transplantation of at least 5000 IEQs/kg of body weight [125]. The primary outcomes will be cleared on May 2022. Another clinical trial from the University of Alberta was immaturely completed on November 2019 (NCT02821026) with no available results in published articles. The outcome of clinical intraomental islet transplantation is not fully discussed at this present moment. Further studies are needed.
Under this clinical trial, a case of laparoscopic ITx (602,395 IEQs/53.4 kg) onto the omentum was performed in a 43-year-old woman with type 1 DM who required more than 30 units of daily insulin and had episodes of unawareness and severe hypoglycemia [125]. Ameliorations in plasma C-peptide level, β cell function, and insulin sensitivity and resistance using the homeostasis model assessment index could be detectable 1 year after transplantation. This case report showed that severe metabolic condition can be recovered by intraomental ITx if sufficient numbers of islets can be implanted.
Finally, we touch on the issue of brown adipose tissue, which is the other type of adipose tissue located in the interscapular, supraclavicular, or para-aortic regions. Brown adipose tissue is a hypervascularized tissue [126] that contributes to maintaining body temperature. As a transplant site for ITx, brown adipose tissue has a unique rich vascularization that might support the engraftment of islets. On the other hand, the distribution of brown adipose tissue is limited, whereas white adipose tissue is located in the entire body. Recently, Xu et al. examined the transplant efficacy of ITx into white and brown adipose tissues. Their data did not show the superiority of brown adipose tissue compared with white adipose tissue in transplant efficacy [127].
5. Conclusions
The usefulness of white adipose tissue as a candidate transplant site for ITx has been proven in various animal studies. Clinical trials using white adipose tissue are ongoing.
Author Contributions
Conceptualization, N.S.; writing—original draft preparation, N.S.; writing—review and editing, G.Y.; supervision, S.K. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by a Grant-in-Aid for Scientific Research (C) (grant number 19K09839, NS) from the Ministry of Education, Culture, Sports, Science, and Technology of Japan and an intramural grant from Fukuoka University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bellin, M.D.; Kandaswamy, R.; Parkey, J.; Zhang, H.J.; Liu, B.; Ihm, S.H.; Ansite, J.D.; Witson, J.; Bansal-Pakala, P.; Balamurugan, A.N.; et al. Prolonged insulin independence after islet allotransplants in recipients with type 1 diabetes. Am. J. Transplant. 2008, 8, 2463–2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, F.B.; Rickels, M.R.; Alejandro, R.; Hering, B.J.; Wease, S.; Naziruddin, B.; Oberholzer, J.; Odorico, J.S.; Garfinkel, M.R.; Levy, M.; et al. Improvement in outcomes of clinical islet transplantation: 1999–2010. Diabetes Care 2012, 35, 1436–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balamurugan, A.N.; Naziruddin, B.; Lockridge, A.; Tiwari, M.; Loganathan, G.; Takita, M.; Matsumoto, S.; Papas, K.; Trieger, M.; Rainis, H.; et al. Islet product characteristics and factors related to successful human islet transplantation from the Collaborative Islet Transplant Registry (CITR) 1999–2010. Am. J. Transplant. 2014, 14, 2595–2606. [Google Scholar] [CrossRef] [PubMed]
- Hering, B.J.; Clarke, W.R.; Bridges, N.D.; Eggerman, T.L.; Alejandro, R.; Bellin, M.D.; Chaloner, K.; Czarniecki, C.W.; Goldstein, J.S.; Hunsicker, L.G.; et al. Phase 3 Trial of Transplantation of Human Islets in Type 1 Diabetes Complicated by Severe Hypoglycemia. Diabetes Care 2016, 39, 1230–1240. [Google Scholar] [CrossRef] [Green Version]
- Sakata, N.; Hayes, P.; Tan, A.; Chan, N.K.; Mace, J.; Peverini, R.; Sowers, L.; Pearce, W.J.; Chinnock, R.; Obenaus, A.; et al. MRI assessment of ischemic liver after intraportal islet transplantation. Transplantation 2009, 87, 825–830. [Google Scholar] [CrossRef] [Green Version]
- Sakata, N.; Yoshimatsu, G.; Kodama, S. The Spleen as an Optimal Site for Islet Transplantation and a Source of Mesenchymal Stem Cells. Int. J. Mol. Sci. 2018, 19, 1391. [Google Scholar] [CrossRef] [Green Version]
- Naziruddin, B.; Kanak, M.A.; Chang, C.A.; Takita, M.; Lawrence, M.C.; Dennison, A.R.; Onaca, N.; Levy, M.F. Improved outcomes of islet autotransplant after total pancreatectomy by combined blockade of IL-1beta and TNFalpha. Am. J. Transplant. 2018, 18, 2322–2329. [Google Scholar] [CrossRef] [Green Version]
- Ishiyama, K.; Rawson, J.; Omori, K.; Mullen, Y. Liver natural killer cells play a role in the destruction of islets after intraportal transplantation. Transplantation 2011, 91, 952–960. [Google Scholar] [CrossRef]
- Saeki, Y.; Ishiyama, K.; Ishida, N.; Tanaka, Y.; Ohdan, H. Memory-like Liver Natural Killer Cells are Responsible for Islet Destruction in Secondary Islet Transplantation. Sci. Rep. 2019, 9, 1022. [Google Scholar] [CrossRef] [Green Version]
- Bottino, R.; Fernandez, L.A.; Ricordi, C.; Lehmann, R.; Tsan, M.F.; Oliver, R.; Inverardi, L. Transplantation of allogeneic islets of Langerhans in the rat liver: Effects of macrophage depletion on graft survival and microenvironment activation. Diabetes 1998, 47, 316–323. [Google Scholar] [CrossRef]
- Bennet, W.; Groth, C.G.; Larsson, R.; Nilsson, B.; Korsgren, O. Isolated human islets trigger an instant blood mediated inflammatory reaction: Implications for intraportal islet transplantation as a treatment for patients with type 1 diabetes. Ups. J. Med. Sci. 2000, 105, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Sakata, N.; Obenaus, A.; Chan, N.; Mace, J.; Chinnock, R.; Hathout, E. Factors affecting islet graft embolization in the liver of diabetic mice. Islets 2009, 1, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Mellgren, A.; Schnell Landstrom, A.H.; Petersson, B.; Andersson, A. The renal subcapsular site offers better growth conditions for transplanted mouse pancreatic islet cells than the liver or spleen. Diabetologia 1986, 29, 670–672. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; McGrath, K.M.; Bottino, R.; Dons, E.M.; Long, C.; Kumar, G.; Ekser, B.; Echeverri, G.J.; Hata, J.; Haruma, K.; et al. Technique of endoscopic biopsy of islet allografts transplanted into the gastric submucosal space in pigs. Cell Transplant. 2013, 22, 2335–2344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantarelli, E.; Citro, A.; Pellegrini, S.; Mercalli, A.; Melzi, R.; Dugnani, E.; Jofra, T.; Fousteri, G.; Mondino, A.; Piemonti, L. Transplant Site Influences the Immune Response After Islet Transplantation: Bone Marrow Versus Liver. Transplantation 2017, 101, 1046–1055. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Nishinakamura, H.; Kumano, K.; Takahashi, H.; Kodama, S. The Spleen Is an Ideal Site for Inducing Transplanted Islet Graft Expansion in Mice. PLoS ONE 2017, 12, e0170899. [Google Scholar] [CrossRef]
- Sakata, N.; Aoki, T.; Yoshimatsu, G.; Tsuchiya, H.; Hata, T.; Katayose, Y.; Egawa, S.; Unno, M. Strategy for clinical setting in intramuscular and subcutaneous islet transplantation. Diabetes Metab. Res. Rev. 2014, 30, 1–10. [Google Scholar] [CrossRef]
- Merani, S.; Toso, C.; Emamaullee, J.; Shapiro, A.M. Optimal implantation site for pancreatic islet transplantation. Br. J. Surg. 2008, 95, 1449–1461. [Google Scholar] [CrossRef]
- Outzen, H.C.; Leiter, E.H. Transplantation of pancreatic islets into cleared mammary fat pads. Transplantation 1981, 32, 101–105. [Google Scholar] [CrossRef]
- Cuthbertson, R.A.; Mandel, T.E. A comparison of portal versus systemic venous drainage in murine foetal pancreatic islet transplantation. Aust. J. Exp. Biol. Med. Sci. 1986, 64 Pt 2, 175–184. [Google Scholar] [CrossRef]
- Yasunami, Y.; Lacy, P.E.; Finke, E.H. A new site for islet transplantation—A peritoneal-omental pouch. Transplantation 1983, 36, 181–182. [Google Scholar] [CrossRef] [PubMed]
- Ao, Z.; Matayoshi, K.; Lakey, J.R.; Rajotte, R.V.; Warnock, G.L. Survival and function of purified islets in the omental pouch site of outbred dogs. Transplantation 1993, 56, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Kin, T.; Korbutt, G.S.; Rajotte, R.V. Survival and metabolic function of syngeneic rat islet grafts transplanted in the omental pouch. Am. J. Transplant. 2003, 3, 281–285. [Google Scholar] [CrossRef]
- Lu, Y.; Zou, S.; Bertera, S.; Bottino, R.; Cooper, D.K.C.; Liu, Z.; Huang, Y.; Wang, C.; Hong, C.; He, T.; et al. A Method for Islet Transplantation to the Omentum in Mouse. J. Vis. Exp. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarra, V.; Appel, A.A.; Anastasio, M.A.; Opara, E.C.; Brey, E.M. This paper is a winner in the Undergraduate category for the SFB awards: Evaluation of the tissue response to alginate encapsulated islets in an omentum pouch model. J. Biomed. Mater. Res. A 2016, 104, 1581–1590. [Google Scholar] [CrossRef] [Green Version]
- Montazeri, L.; Hojjati-Emami, S.; Bonakdar, S.; Tahamtani, Y.; Hajizadeh-Saffar, E.; Noori-Keshtkar, M.; Najar-Asl, M.; Ashtiani, M.K.; Baharvand, H. Improvement of islet engrafts by enhanced angiogenesis and microparticle-mediated oxygenation. Biomaterials 2016, 89, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Hajizadeh-Saffar, E.; Tahamtani, Y.; Aghdami, N.; Azadmanesh, K.; Habibi-Anbouhi, M.; Heremans, Y.; De Leu, N.; Heimberg, H.; Ravassard, P.; Shokrgozar, M.A.; et al. Inducible VEGF expression by human embryonic stem cell-derived mesenchymal stromal cells reduces the minimal islet mass required to reverse diabetes. Sci. Rep. 2015, 5, 9322. [Google Scholar] [CrossRef]
- Pareta, R.; McQuilling, J.P.; Sittadjody, S.; Jenkins, R.; Bowden, S.; Orlando, G.; Farney, A.C.; Brey, E.M.; Opara, E.C. Long-term function of islets encapsulated in a redesigned alginate microcapsule construct in omentum pouches of immune-competent diabetic rats. Pancreas 2014, 43, 605–613. [Google Scholar] [CrossRef] [Green Version]
- Pedraza, E.; Brady, A.C.; Fraker, C.A.; Molano, R.D.; Sukert, S.; Berman, D.M.; Kenyon, N.S.; Pileggi, A.; Ricordi, C.; Stabler, C.L. Macroporous three-dimensional PDMS scaffolds for extrahepatic islet transplantation. Cell Transplant. 2013, 22, 1123–1135. [Google Scholar] [CrossRef] [Green Version]
- Kriz, J.; Vilk, G.; Mazzuca, D.M.; Toleikis, P.M.; Foster, P.J.; White, D.J. A novel technique for the transplantation of pancreatic islets within a vascularized device into the greater omentum to achieve insulin independence. Am. J. Surg. 2012, 203, 793–797. [Google Scholar] [CrossRef]
- Gupta, R.; Sefton, M.V. Application of an endothelialized modular construct for islet transplantation in syngeneic and allogeneic immunosuppressed rat models. Tissue Eng. Part A 2011, 17, 2005–2015. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.I.; Yu, J.E.; Park, C.G.; Kim, S.J. Comparison of four pancreatic islet implantation sites. J. Korean Med. Sci. 2010, 25, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Solari, M.G.; Srinivasan, S.; Boumaza, I.; Unadkat, J.; Harb, G.; Garcia-Ocana, A.; Feili-Hariri, M. Marginal mass islet transplantation with autologous mesenchymal stem cells promotes long-term islet allograft survival and sustained normoglycemia. J. Autoimmun. 2009, 32, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Berman, D.M.; O’Neil, J.J.; Coffey, L.C.; Chaffanjon, P.C.; Kenyon, N.M.; Ruiz, P., Jr.; Pileggi, A.; Ricordi, C.; Kenyon, N.S. Long-term survival of nonhuman primate islets implanted in an omental pouch on a biodegradable scaffold. Am. J. Transplant. 2009, 9, 91–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kin, T.; O’Neil, J.J.; Pawlick, R.; Korbutt, G.S.; Shapiro, A.M.; Lakey, J.R. The use of an approved biodegradable polymer scaffold as a solid support system for improvement of islet engraftment. Artif. Organs 2008, 32, 990–993. [Google Scholar] [CrossRef]
- Kobayashi, T.; Aomatsu, Y.; Iwata, H.; Kin, T.; Kanehiro, H.; Hisanga, M.; Ko, S.; Nagao, M.; Harb, G.; Nakajima, Y. Survival of microencapsulated islets at 400 days posttransplantation in the omental pouch of NOD mice. Cell Transplant. 2006, 15, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Guan, J.; Behme, M.T.; Zucker, P.; Atkison, P.; Hramiak, I.; Zhong, R.; Dupre, J. Glucose turnover and insulin sensitivity in rats with pancreatic islet transplants. Diabetes 1998, 47, 1020–1026. [Google Scholar] [CrossRef]
- al-Abdullah, I.H.; Anil Kumar, M.S.; Kelly-Sullivan, D.; Abouna, G.M. Site for unpurified islet transplantation is an important parameter for determination of the outcome of graft survival and function. Cell Transplant. 1995, 4, 297–305. [Google Scholar] [CrossRef]
- Kasoju, N.; Patikova, A.; Wawrzynska, E.; Vojtiskova, A.; Sedlacik, T.; Kumorek, M.; Pop-Georgievski, O.; Sticova, E.; Kri, Z.J.; Kubies, D. Bioengineering a pre-vascularized pouch for subsequent islet transplantation using VEGF-loaded polylactide capsules. Biomater. Sci. 2020, 8, 631–647. [Google Scholar] [CrossRef] [Green Version]
- Tuch, B.E.; Sheil, A.G.; Ng, A.B.; Turtle, J.R. Long-term survival of human fetal pancreatic tissue transplanted into an insulin-dependent diabetic patient. Diabet Med. 1986, 3, 24–28. [Google Scholar] [CrossRef]
- Chusyd, D.E.; Wang, D.; Huffman, D.M.; Nagy, T.R. Relationships between Rodent White Adipose Fat Pads and Human White Adipose Fat Depots. Front. Nutr. 2016, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Weaver, J.D.; Headen, D.M.; Aquart, J.; Johnson, C.T.; Shea, L.D.; Shirwan, H.; Garcia, A.J. Vasculogenic hydrogel enhances islet survival, engraftment, and function in leading extrahepatic sites. Sci. Adv. 2017, 3, e1700184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajotte, R.V.; Tighe, V.M.; Warnock, G.L.; Kneteman, N.M.; Finegood, D.T. New site for islet transplantation in rats—Liver-mesentery or liver-omental pouch. Transplant. Proc. 1994, 26, 675. [Google Scholar] [PubMed]
- Michalska, W.; Garnuszek, P.; Licinska, I.; Wilgus, J.; Szymanska, K.; Rowinski, W.; Mazurek, A.P.; Fiedor, P. Monitoring of pancreatic islets transplanted to colon mesentery. Transplant. Proc. 2002, 34, 653–654. [Google Scholar] [CrossRef]
- Hendrawan, S.; Yusuf, I.; Hatta, M.; Aman, M.; Patellongi, I.; Serra, A.L.; Lawrence, G.; Weber, U.; Sutedja, B.; Baer, H.U. Allogeneic islet cells implant on poly-l-lactide matrix to reduce hyperglycaemia in streptozotocin-induced diabetic rat. Pancreatology 2017, 17, 411–418. [Google Scholar] [CrossRef]
- Phelps, E.A.; Templeman, K.L.; Thule, P.M.; Garcia, A.J. Engineered VEGF-releasing PEG-MAL hydrogel for pancreatic islet vascularization. Drug Deliv. Transl. Res. 2015, 5, 125–136. [Google Scholar] [CrossRef]
- Phelps, E.A.; Headen, D.M.; Taylor, W.R.; Thule, P.M.; Garcia, A.J. Vasculogenic bio-synthetic hydrogel for enhancement of pancreatic islet engraftment and function in type 1 diabetes. Biomaterials 2013, 34, 4602–4611. [Google Scholar] [CrossRef] [Green Version]
- Vernon, R.B.; Preisinger, A.; Gooden, M.D.; D’Amico, L.A.; Yue, B.B.; Bollyky, P.L.; Kuhr, C.S.; Hefty, T.R.; Nepom, G.T.; Gebe, J.A. Reversal of diabetes in mice with a bioengineered islet implant incorporating a type I collagen hydrogel and sustained release of vascular endothelial growth factor. Cell Transplant. 2012, 21, 2099–2110. [Google Scholar] [CrossRef] [Green Version]
- Minardi, S.; Guo, M.; Zhang, X.; Luo, X. An elastin-based vasculogenic scaffold promotes marginal islet mass engraftment and function at an extrahepatic site. J. Immunol. Regen. Med. 2019, 3, 1–12. [Google Scholar] [CrossRef]
- Najjar, M.; Manzoli, V.; Abreu, M.; Villa, C.; Martino, M.M.; Molano, R.D.; Torrente, Y.; Pileggi, A.; Inverardi, L.; Ricordi, C.; et al. Fibrin gels engineered with pro-angiogenic growth factors promote engraftment of pancreatic islets in extrahepatic sites in mice. Biotechnol. Bioeng. 2015, 112, 1916–1926. [Google Scholar] [CrossRef]
- Liu, J.M.H.; Zhang, X.; Joe, S.; Luo, X.; Shea, L.D. Evaluation of biomaterial scaffold delivery of IL-33 as a localized immunomodulatory agent to support cell transplantation in adipose tissue. J. Immunol. Regen. Med. 2018, 1, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.M.H.; Zhang, J.; Zhang, X.; Hlavaty, K.A.; Ricci, C.F.; Leonard, J.N.; Shea, L.D.; Gower, R.M. Transforming growth factor-beta 1 delivery from microporous scaffolds decreases inflammation post-implant and enhances function of transplanted islets. Biomaterials 2016, 80, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibly, R.F.; Zhang, X.; Graham, M.L.; Hering, B.J.; Kaufman, D.B.; Lowe, W.L., Jr.; Shea, L.D. Extrahepatic islet transplantation with microporous polymer scaffolds in syngeneic mouse and allogeneic porcine models. Biomaterials 2011, 32, 9677–9684. [Google Scholar] [CrossRef] [Green Version]
- Kheradmand, T.; Wang, S.; Gibly, R.F.; Zhang, X.; Holland, S.; Tasch, J.; Graham, J.G.; Kaufman, D.B.; Miller, S.D.; Shea, L.D.; et al. Permanent protection of PLG scaffold transplanted allogeneic islet grafts in diabetic mice treated with ECDI-fixed donor splenocyte infusions. Biomaterials 2011, 32, 4517–4524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rios, P.D.; Zhang, X.; Luo, X.; Shea, L.D. Mold-casted non-degradable, islet macro-encapsulating hydrogel devices for restoration of normoglycemia in diabetic mice. Biotechnol. Bioeng. 2016, 113, 2485–2495. [Google Scholar] [CrossRef] [PubMed]
- Brubaker, C.E.; Kissler, H.; Wang, L.J.; Kaufman, D.B.; Messersmith, P.B. Biological performance of mussel-inspired adhesive in extrahepatic islet transplantation. Biomaterials 2010, 31, 420–427. [Google Scholar] [CrossRef] [Green Version]
- Salvay, D.M.; Rives, C.B.; Zhang, X.; Chen, F.; Kaufman, D.B.; Lowe, W.L., Jr.; Shea, L.D. Extracellular matrix protein-coated scaffolds promote the reversal of diabetes after extrahepatic islet transplantation. Transplantation 2008, 85, 1456–1464. [Google Scholar] [CrossRef]
- Manzoli, V.; Villa, C.; Bayer, A.L.; Morales, L.C.; Molano, R.D.; Torrente, Y.; Ricordi, C.; Hubbell, J.A.; Tomei, A.A. Immunoisolation of murine islet allografts in vascularized sites through conformal coating with polyethylene glycol. Am. J. Transplant. 2018, 18, 590–603. [Google Scholar] [CrossRef]
- Villa, C.; Manzoli, V.; Abreu, M.M.; Verheyen, C.A.; Seskin, M.; Najjar, M.; Molano, R.D.; Torrente, Y.; Ricordi, C.; Tomei, A.A. Effects of Composition of Alginate-Polyethylene Glycol Microcapsules and Transplant Site on Encapsulated Islet Graft Outcomes in Mice. Transplantation 2017, 101, 1025–1035. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.Y.; Nothias, J.M.; Scavone, A.; Garfinkel, M.; Millis, J.M. Biocompatibility investigation of polyethylene glycol and alginate-poly-L-lysine for islet encapsulation. ASAIO J. 2010, 56, 241–245. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Emami, F.; Yook, S.; Nguyen, H.T.; Pham, T.T.; Pathak, S.; Regmi, S.; Kim, J.O.; Yong, C.S.; Kim, J.R.; et al. Local release of NECA (5′-(N-ethylcarboxamido)adenosine) from implantable polymeric sheets for enhanced islet revascularization in extrahepatic transplantation site. J. Control Release 2020, 321, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Wang, X.; Han, C.S.; Chen, L.Y.; Luo, Y. Scaffold-supported Transplantation of Islets in the Epididymal Fat Pad of Diabetic Mice. J. Vis. Exp. 2017, 54995. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, K.; Zhang, W.; Qiang, M.; Luo, Y. A bilaminated decellularized scaffold for islet transplantation: Structure, properties and functions in diabetic mice. Biomaterials 2017, 138, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.; Zhu, M.; Zhang, X.; Ma, R.; Yang, X.; Ke, T.; Wang, L.; Li, Z.; Kong, D.; Li, C. A macroporous heparin-releasing silk fibroin scaffold improves islet transplantation outcome by promoting islet revascularisation and survival. Acta Biomater. 2017, 59, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Buitinga, M.; Assen, F.; Hanegraaf, M.; Wieringa, P.; Hilderink, J.; Moroni, L.; Truckenmuller, R.; van Blitterswijk, C.; Romer, G.W.; Carlotti, F.; et al. Micro-fabricated scaffolds lead to efficient remission of diabetes in mice. Biomaterials 2017, 135, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Gibly, R.F.; Zhang, X.; Lowe, W.L., Jr.; Shea, L.D. Porous scaffolds support extrahepatic human islet transplantation, engraftment, and function in mice. Cell Transplant. 2013, 22, 811–819. [Google Scholar] [CrossRef] [Green Version]
- Brady, A.C.; Martino, M.M.; Pedraza, E.; Sukert, S.; Pileggi, A.; Ricordi, C.; Hubbell, J.A.; Stabler, C.L. Proangiogenic hydrogels within macroporous scaffolds enhance islet engraftment in an extrahepatic site. Tissue Eng. Part A 2013, 19, 2544–2552. [Google Scholar] [CrossRef] [Green Version]
- Stiegler, P.; Matzi, V.; Pierer, E.; Hauser, O.; Schaffellner, S.; Renner, H.; Greilberger, J.; Aigner, R.; Maier, A.; Lackner, C.; et al. Creation of a prevascularized site for cell transplantation in rats. Xenotransplantation 2010, 17, 379–390. [Google Scholar] [CrossRef]
- Forster, N.A.; Penington, A.J.; Hardikar, A.A.; Palmer, J.A.; Hussey, A.; Tai, J.; Morrison, W.A.; Feeney, S.J. A prevascularized tissue engineering chamber supports growth and function of islets and progenitor cells in diabetic mice. Islets 2011, 3, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Yasunami, Y.; Nakafusa, Y.; Nitta, N.; Nakamura, M.; Goto, M.; Ono, J.; Taniguchi, M. A Novel Subcutaneous Site of Islet Transplantation Superior to the Liver. Transplantation 2018, 102, 945–952. [Google Scholar] [CrossRef]
- Gregoire, F.M.; Smas, C.M.; Sul, H.S. Understanding adipocyte differentiation. Physiol. Rev. 1998, 78, 783–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Divoux, A.; Clement, K. Architecture and the extracellular matrix: The still unappreciated components of the adipose tissue. Obes. Rev. 2011, 12, e494–e503. [Google Scholar] [CrossRef] [PubMed]
- Weiner, F.R.; Shah, A.; Smith, P.J.; Rubin, C.S.; Zern, M.A. Regulation of collagen gene expression in 3T3-L1 cells. Effects of adipocyte differentiation and tumor necrosis factor alpha. Biochemistry 1989, 28, 4094–4099. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O.; Zhao, Q. The evolution of cell adhesion. J. Cell Biol. 2000, 150, F89–F96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popova, S.N.; Lundgren-Akerlund, E.; Wiig, H.; Gullberg, D. Physiology and pathology of collagen receptors. Acta Physiol. 2007, 190, 179–187. [Google Scholar] [CrossRef]
- Kantengwa, S.; Baetens, D.; Sadoul, K.; Buck, C.A.; Halban, P.A.; Rouiller, D.G. Identification and characterization of alpha 3 beta 1 integrin on primary and transformed rat islet cells. Exp. Cell Res. 1997, 237, 394–402. [Google Scholar] [CrossRef]
- Belkin, A.M.; Stepp, M.A. Integrins as receptors for laminins. Microsc. Res. Tech. 2000, 51, 280–301. [Google Scholar] [CrossRef]
- Johansson, S.; Svineng, G.; Wennerberg, K.; Armulik, A.; Lohikangas, L. Fibronectin-integrin interactions. Front. Biosci. 1997, 2, d126–d146. [Google Scholar] [CrossRef] [Green Version]
- Hamidi, H.; Ivaska, J. Vascular Morphogenesis: An Integrin and Fibronectin Highway. Curr. Biol. 2017, 27, R158–R161. [Google Scholar] [CrossRef]
- Olaniru, O.E.; Persaud, S.J. Identifying novel therapeutic targets for diabetes through improved understanding of islet adhesion receptors. Curr. Opin. Pharmacol. 2018, 43, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Kaido, T.; Yebra, M.; Cirulli, V.; Montgomery, A.M. Regulation of human beta-cell adhesion, motility, and insulin secretion by collagen IV and its receptor alpha1beta1. J. Biol. Chem. 2004, 279, 53762–53769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosco, D.; Meda, P.; Halban, P.A.; Rouiller, D.G. Importance of cell-matrix interactions in rat islet beta-cell secretion in vitro: Role of alpha6beta1 integrin. Diabetes 2000, 49, 233–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Li, J.; Lyte, K.; Yashpal, N.K.; Fellows, F.; Goodyer, C.G. Role for beta1 integrin and its associated alpha3, alpha5, and alpha6 subunits in development of the human fetal pancreas. Diabetes 2005, 54, 2080–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yashpal, N.K.; Li, J.; Wheeler, M.B.; Wang, R. Expression of {beta}1 integrin receptors during rat pancreas development—Sites and dynamics. Endocrinology 2005, 146, 1798–1807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleem, S.; Li, J.; Yee, S.P.; Fellows, G.F.; Goodyer, C.G.; Wang, R. beta1 integrin/FAK/ERK signalling pathway is essential for human fetal islet cell differentiation and survival. J. Pathol. 2009, 219, 182–192. [Google Scholar] [CrossRef]
- Krishnamurthy, M.; Li, J.; Fellows, G.F.; Rosenberg, L.; Goodyer, C.G.; Wang, R. Integrin {alpha}3, but not {beta}1, regulates islet cell survival and function via PI3K/Akt signaling pathways. Endocrinology 2011, 152, 424–435. [Google Scholar] [CrossRef] [Green Version]
- Baer, P.C.; Geiger, H. Adipose-derived mesenchymal stromal/stem cells: Tissue localization, characterization, and heterogeneity. Stem Cells Int. 2012, 2012, 812693. [Google Scholar] [CrossRef] [Green Version]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [Green Version]
- Cavallari, G.; Olivi, E.; Bianchi, F.; Neri, F.; Foroni, L.; Valente, S.; La Manna, G.; Nardo, B.; Stefoni, S.; Ventura, C. Mesenchymal stem cells and islet cotransplantation in diabetic rats: Improved islet graft revascularization and function by human adipose tissue-derived stem cells preconditioned with natural molecules. Cell Transplant. 2012, 21, 2771–2781. [Google Scholar] [CrossRef] [Green Version]
- Bhang, S.H.; Jung, M.J.; Shin, J.Y.; La, W.G.; Hwang, Y.H.; Kim, M.J.; Kim, B.S.; Lee, D.Y. Mutual effect of subcutaneously transplanted human adipose-derived stem cells and pancreatic islets within fibrin gel. Biomaterials 2013, 34, 7247–7256. [Google Scholar] [CrossRef]
- Arzouni, A.A.; Vargas-Seymour, A.; Rackham, C.L.; Dhadda, P.; Huang, G.C.; Choudhary, P.; Nardi, N.; King, A.J.F.; Jones, P.M. Mesenchymal stromal cells improve human islet function through released products and extracellular matrix. Clin. Sci. 2017, 131, 2835–2845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, G.; Rezaee, M.; Razavi, M.; Taysir, A.; Wang, J.; Thakor, A.S. Adipose tissue-derived mesenchymal stem cells rescue the function of islets transplanted in sub-therapeutic numbers via their angiogenic properties. Cell Tissue Res. 2019, 376, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Navaei-Nigjeh, M.; Moloudizargari, M.; Baeeri, M.; Gholami, M.; Lotfibakhshaiesh, N.; Soleimani, M.; Vasheghani-Farahani, E.; Ai, J.; Abdollahi, M. Reduction of marginal mass required for successful islet transplantation in a diabetic rat model using adipose tissue-derived mesenchymal stromal cells. Cytotherapy 2018, 20, 1124–1142. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Sun, Z.; Kim, D.S.; Gou, W.; Strange, C.; Dong, H.; Cui, W.; Gilkeson, G.; Morgan, K.A.; Adams, D.B.; et al. Adipose stem cells from chronic pancreatitis patients improve mouse and human islet survival and function. Stem Cell Res. Ther. 2017, 8, 192. [Google Scholar] [CrossRef]
- Ohmura, Y.; Tanemura, M.; Kawaguchi, N.; Machida, T.; Tanida, T.; Deguchi, T.; Wada, H.; Kobayashi, S.; Marubashi, S.; Eguchi, H.; et al. Combined transplantation of pancreatic islets and adipose tissue-derived stem cells enhances the survival and insulin function of islet grafts in diabetic mice. Transplantation 2010, 90, 1366–1373. [Google Scholar] [CrossRef]
- Mohammadi Ayenehdeh, J.; Niknam, B.; Rasouli, S.; Hashemi, S.M.; Rahavi, H.; Rezaei, N.; Soleimani, M.; Liaeiha, A.; Niknam, M.H.; Tajik, N. Immunomodulatory and protective effects of adipose tissue-derived mesenchymal stem cells in an allograft islet composite transplantation for experimental autoimmune type 1 diabetes. Immunol. Lett. 2017, 188, 21–31. [Google Scholar] [CrossRef]
- Kuppan, P.; Seeberger, K.; Kelly, S.; Rosko, M.; Adesida, A.; Pepper, A.R.; Korbutt, G.S. Co-transplantation of human adipose-derived mesenchymal stem cells with neonatal porcine islets within a prevascularized subcutaneous space augments the xenograft function. Xenotransplantation 2020, e12581. [Google Scholar] [CrossRef]
- Tanaka, T.; Kojima, D.; Mera, T.; Matsumoto, M.; Yasunami, Y.; Yanase, T. Expansion of transplanted islets in mice by co-transplantation with adipose tissue-derived mesenchymal stem cells. Heliyon 2018, 4, e00632. [Google Scholar] [CrossRef]
- Karaoz, E.; Okcu, A.; Unal, Z.S.; Subasi, C.; Saglam, O.; Duruksu, G. Adipose tissue-derived mesenchymal stromal cells efficiently differentiate into insulin-producing cells in pancreatic islet microenvironment both in vitro and in vivo. Cytotherapy 2013, 15, 557–570. [Google Scholar] [CrossRef]
- Anitha, R.; Vaikkath, D.; Shenoy, S.J.; Nair, P.D. Tissue-engineered islet-like cell clusters generated from adipose tissue-derived stem cells on three-dimensional electrospun scaffolds can reverse diabetes in an experimental rat model and the role of porosity of scaffolds on cluster differentiation. J. Biomed. Mater. Res. A 2020, 108, 749–759. [Google Scholar] [CrossRef]
- Badimon, L.; Cubedo, J. Adipose tissue depots and inflammation: Effects on plasticity and resident mesenchymal stem cell function. Cardiovasc. Res. 2017, 113, 1064–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.K. Adipocytes. Curr. Biol. 2014, 24, R988–R993. [Google Scholar] [CrossRef] [Green Version]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose expression of tumor necrosis factor-alpha: Direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Uysal, K.T.; Becherer, J.D.; Arner, P.; Hotamisligil, G.S. Altered tumor necrosis factor-alpha (TNF-alpha) processing in adipocytes and increased expression of transmembrane TNF-alpha in obesity. Diabetes 2002, 51, 1876–1883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalupahana, N.S.; Claycombe, K.J.; Moustaid-Moussa, N. (n-3) Fatty acids alleviate adipose tissue inflammation and insulin resistance: Mechanistic insights. Adv. Nutr. 2011, 2, 304–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Mizuarai, S.; Araki, H.; Mashiko, S.; Ishihara, A.; Kanatani, A.; Itadani, H.; Kotani, H. Adiposity elevates plasma MCP-1 levels leading to the increased CD11b-positive monocytes in mice. J. Biol. Chem. 2003, 278, 46654–46660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamei, N.; Tobe, K.; Suzuki, R.; Ohsugi, M.; Watanabe, T.; Kubota, N.; Ohtsuka-Kowatari, N.; Kumagai, K.; Sakamoto, K.; Kobayashi, M.; et al. Overexpression of monocyte chemoattractant protein-1 in adipose tissues causes macrophage recruitment and insulin resistance. J. Biol. Chem. 2006, 281, 26602–26614. [Google Scholar] [CrossRef] [Green Version]
- Kanda, H.; Tateya, S.; Tamori, Y.; Kotani, K.; Hiasa, K.; Kitazawa, R.; Kitazawa, S.; Miyachi, H.; Maeda, S.; Egashira, K.; et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J. Clin. Investig. 2006, 116, 1494–1505. [Google Scholar] [CrossRef]
- Melzi, R.; Mercalli, A.; Sordi, V.; Cantarelli, E.; Nano, R.; Maffi, P.; Sitia, G.; Guidotti, L.G.; Secchi, A.; Bonifacio, E.; et al. Role of CCL2/MCP-1 in islet transplantation. Cell Transplant. 2010, 19, 1031–1046. [Google Scholar] [CrossRef]
- Citro, A.; Pellegrini, S.; Dugnani, E.; Eulberg, D.; Klussmann, S.; Piemonti, L. CCL2/MCP-1 and CXCL12/SDF-1 blockade by L-aptamers improve pancreatic islet engraftment and survival in mouse. Am. J. Transplant. 2019, 19, 3131–3138. [Google Scholar] [CrossRef]
- Min, B.H.; Shin, J.S.; Kim, J.M.; Kang, S.J.; Kim, H.J.; Yoon, I.H.; Park, S.K.; Choi, J.W.; Lee, M.S.; Park, C.G. Delayed revascularization of islets after transplantation by IL-6 blockade in pig to non-human primate islet xenotransplantation model. Xenotransplantation 2018, 25. [Google Scholar] [CrossRef] [PubMed]
- Landecho, M.F.; Tuero, C.; Valenti, V.; Bilbao, I.; de la Higuera, M.; Fruhbeck, G. Relevance of Leptin and Other Adipokines in Obesity-Associated Cardiovascular Risk. Nutrients 2019, 11, 2664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denroche, H.C.; Quong, W.L.; Bruin, J.E.; Tuduri, E.; Asadi, A.; Glavas, M.M.; Fox, J.K.; Kieffer, T.J. Leptin administration enhances islet transplant performance in diabetic mice. Diabetes 2013, 62, 2738–2746. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Ravazzola, M.; Park, B.H.; Bashmakov, Y.K.; Orci, L.; Unger, R.H. Metabolic mechanisms of failure of intraportally transplanted pancreatic beta-cells in rats: Role of lipotoxicity and prevention by leptin. Diabetes 2007, 56, 2295–2301. [Google Scholar] [CrossRef] [Green Version]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohashi, K.; Shibata, R.; Murohara, T.; Ouchi, N. Role of anti-inflammatory adipokines in obesity-related diseases. Trends Endocrinol. Metab. 2014, 25, 348–355. [Google Scholar] [CrossRef]
- Ohashi, K.; Ouchi, N.; Matsuzawa, Y. Anti-inflammatory and anti-atherogenic properties of adiponectin. Biochimie 2012, 94, 2137–2142. [Google Scholar] [CrossRef]
- Antoniades, C.; Antonopoulos, A.S.; Tousoulis, D.; Stefanadis, C. Adiponectin: From obesity to cardiovascular disease. Obes. Rev. 2009, 10, 269–279. [Google Scholar] [CrossRef]
- Du, X.; He, S.; Jiang, Y.; Wei, L.; Hu, W. Adiponectin prevents islet ischemia-reperfusion injury through the COX2-TNFalpha-NF-kappaB-dependent signal transduction pathway in mice. J. Endocrinol. 2013, 218, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Delaune, V.; Berney, T.; Lacotte, S.; Toso, C. Intraportal islet transplantation: The impact of the liver microenvironment. Transpl. Int. 2017, 30, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Investig. 2007, 117, 175–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, L.; Lumeng, C.N. Properties and functions of adipose tissue macrophages in obesity. Immunology 2018, 155, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Stice, M.J.; Dunn, T.B.; Bellin, M.D.; Skube, M.E.; Beilman, G.J. Omental Pouch Technique for Combined Site Islet Autotransplantation Following Total Pancreatectomy. Cell Transplant. 2018, 27, 1561–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baidal, D.A.; Ricordi, C.; Berman, D.M.; Alvarez, A.; Padilla, N.; Ciancio, G.; Linetsky, E.; Pileggi, A.; Alejandro, R. Bioengineering of an Intraabdominal Endocrine Pancreas. N. Engl. J. Med. 2017, 376, 1887–1889. [Google Scholar] [CrossRef] [Green Version]
- Sacks, H.; Symonds, M.E. Anatomical locations of human brown adipose tissue: Functional relevance and implications in obesity and type 2 diabetes. Diabetes 2013, 62, 1783–1790. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Xie, R.; Lin, X.; Jia, J.; Zeng, N.; Li, W.; Xiao, D.; Du, T. Brown adipose tissue: A potential site for islet transplantation. Transplantation 2020. [Google Scholar] [CrossRef]
Figure 1.
Characteristics of white adipose tissue as the transplant site for islets.


{kind=link}
{kind=link}
Table 1.
Candidates for transplant site for islets and their characteristics.
| Advantages | Disadvantages | |
|---|---|---|
| Liver |
|
|
| Kidney |
|
|
| Spleen |
|
|
| Muscle/ subcutaneous tissue |
|
|
| Omentum (white adipose tissue) |
|
|
| Mesentery (white adipose tissue) |
|
|
IBMIR, instant blood-mediated inflammatory reaction.
Table 2.
Representative studies regarding the omental pouch.
| Author (Year) | Transplant Model (Animal) | Number of Transplanted Islets | Additional Treatment | Outcome | Reference |
|---|---|---|---|---|---|
| Kasoju (2020) | Syngeneic (rat) | Not described | Using biomaterial spacer and growth factor | Islet engraftment | [39] |
| Lu (2019) | Not described | 450–500 islets | Using hydrogel | Normoglycemia achieved immediately | [24] |
| Ibarra (2016) | Allogeneic (rat) | 800–1000 beads (accurate number not described) | Encapsulation | Transplant efficacy was unclear | [25] |
| Montazeri (2016) | Xenogeneic (rat to nude mouse) | 250 islets (IEQs) | Using oxygenation technique with growth factor | Improved blood glucose level | [26] |
| Hajizadeh-Saffar (2015) | Allogeneic (mouse) | 200 islets (IEQs) 400 islets (IEQs) | Co-transplantation with growth factor-releasing cells (derived from mesenchymal stromal cells) | Normoglycemia rates were 80% in 200 IEQs with MSCs and 40% in 400 IEQs | [27] |
| Pareta (2014) | Syngeneic Allogeneic (rat) | 800 islets | Encapsulation | Failed to achieve normoglycemia | [28] |
| Pedraza (2013) | Syngeneic (rat) | 1800 islets | With or without bioscaffold | Achieved normoglycemia over 110 days | [29] |
| Kriz (2012) | Syngeneic (rat) | 10,000 islets (IEQs)/kg (2000–3000 IEQs) | Using biomaterial spacer | Normoglycemia rate was 70% at 100 days after transplantation | [30] |
| Gupta (2011) | Syngeneic Allogeneic (rat) | 2000 islets | Using endothelialized modules | Normoglycemia rate was 40% (syngeneic) | [31] |
| Berman (2009) | Autologous Allogeneic (monkey) | 5093 IEQ/kg (autologous) 4200–14,544 IEQ/kg (allogeneic) | Bioscaffold Immunosuppressants (allogeneic) | Achieved normoglycemia in autologous islet transplantation. Therapeutic effect was similar to that of intrahepatic islet transplantation | [34] |
| Kobayashi (2006) | Syngeneic (mouse) | 1500 islets | Encapsulation | The normoglycemia rate was 90% over 100 days after transplantation | [36] |
| Kin (2003) | Syngeneic (rat) | 2000 islets | No additional treatment | Achieving normoglycemia at 56 days after transplantation | [23] |
| Guan (1998) | Syngeneic (rat) | Approximately 3000 islets | No additional treatment | Achieving normoglycemia at 2 months after transplantation | [37] |
IEQ, islet equivalent.
Table 3.
Representative studies regarding the epididymal fat pad.
| Author (Year) | Transplant Model | Number of Transplanted Islets | Additional Treatment | Outcome | Reference |
|---|---|---|---|---|---|
| Nguyen (2020) | Xenogeneic (rat to nude mouse) | 500 islets (IEQs) | With or without bioscaffold | Achieved normoglycemia (88% with bioscaffold, 44% without bioscaffold) | [61] |
| Minardi (2019) | Syngeneic (mouse) | 70 islets | With bioscaffold | Achieved normoglycemia at 1 month after transplantation | [49] |
| Liu (2018) | Syngeneic Allogeneic (mouse) | 250 islets | Using immunomodulation technique and bioscaffolds | Achieved normoglycemia in all mice | [51] |
| Manzoli (2018) | Allogeneic (mouse) | 750–1000 islets (IEQs) | Encapsulation | Achieved normoglycemia at 10 days after transplantation | [58] |
| Weaver (2017) | Syngeneic (mice) | 600 islets (IEQs) | Using bioscaffold with growth factor | The normoglycemia rates were 75% and 60% using bioscaffold with and without growth factor, respectively | [42] |
| Wang (2017) | Syngeneic (mouse) | 150–500 islets | Using bioscaffold | Mice achieved normoglycemia: 10/12 (500 islets + scaffold), 10/15 (250 islets + scaffold), 9/19 (150 islets + scaffold), 3/10 (250 islets) | [62,63] |
| Mao (2017) | Syngeneic (mouse) | 300 islets | Using bioscaffold | No mice achieved normoglycemia in ITx only. All mice achieved normoglycemia with bioscaffold | [64] |
| Buitinga (2017) | Syngeneic (mouse) | 300 islets | Using bioscaffold | Achieved normoglycemia (75% with bioscaffold, 29% without bioscaffold) | [65] |
| Villa (2017) | Allogeneic (mouse) Xeneogeneic (baboon to NOD/scid mouse) | 750 islets (IEQs) | Encapsulation | Allogeneic: Normoglycemia achieved and maintained in all the mice (7/7) for 100 days after transplantation Xenogeneic: Normoglycemia achieved and maintained in all the mice (4/4) for 30 days after transplantation | [59] |
| Rios (2016) | Syngeneic (mouse) | 300 and 500 islets | Using bioscaffold | Achieved normoglycemia (100% in 500 islets, 25% in 300 islets) | [55] |
| Liu (2016) | Syngeneic (mouse) | 250 islets | Using bioscaffold | Normoglycemia achieved and maintained for 80 days after transplantation | [52] |
| Najjar (2015) | Syngeneic (mouse) | 250 islets (IEQs) | Using bioscaffold | Achieved normoglycemia (60% with bioscaffold, 10% without bioscaffold) | [50] |
| Gibly (2013) | Xenogeneic (Human to NOD/scid mice) | 2000 islets (IEQs) | Using bioscaffold | Normoglycemia achieved and maintained over 140 days after transplantation | [66] |
| Brady (2013) | Syngeneic (mouse) | 250 islets (IEQs) | Using bioscaffold | Achieved normoglycemia (100% with bioscaffold, 87.5% without bioscaffold) | [67] |
| Gibly (2011) | Syngeneic (mouse) | 75 islets | Using bioscaffold | Normoglycemia achieved and maintained for 42 days after transplantation | [53] |
| Kheradmand (2011) | Allogeneic (mouse) | 500 islets | Using bioscaffold with splenocytes | Achieved normoglycemia for 150 days (80% with splenocytes) | [54] |
| Brubaker (2010) | Syngeneic (mouse) | 150 islets | Using bioscaffold | Normoglycemia achieved and maintained for 110 days after transplantation (both with and without bioscaffold) | [56] |
| Salvay (2008) | Syngeneic (mouse) | 125 islets | Using bioscaffold | Normoglycemia achieved and maintained for 300 days after transplantation | [57] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sakata, N.; Yoshimatsu, G.; Kodama, S. White Adipose Tissue as a Site for Islet Transplantation. Transplantology 2020, 1, 55-70. https://0-doi-org.brum.beds.ac.uk/10.3390/transplantology1020006
AMA Style
Sakata N, Yoshimatsu G, Kodama S. White Adipose Tissue as a Site for Islet Transplantation. Transplantology. 2020; 1(2):55-70. https://0-doi-org.brum.beds.ac.uk/10.3390/transplantology1020006
Chicago/Turabian StyleSakata, Naoaki, Gumpei Yoshimatsu, and Shohta Kodama. 2020. "White Adipose Tissue as a Site for Islet Transplantation" Transplantology 1, no. 2: 55-70. https://0-doi-org.brum.beds.ac.uk/10.3390/transplantology1020006