Nonstructural Proteins of Alphavirus—Potential Targets for Drug Development


1
Singapore Immunology Network, Agency for Science, Technology and Research (A*STAR), Singapore 138648, Singapore
2
School of Biological Sciences, Nanyang Technological University, Singapore 636921, Singapore
3
Department of Biochemistry, Yong Loo Lin School of Medicine, National University of Singapore, Singapore 117596, Singapore
4
Institute of Infection and Global Health, University of Liverpool, Liverpool L69 7BE, UK
*
Author to whom correspondence should be addressed.
Viruses 2018, 10(2), 71; https://0-doi-org.brum.beds.ac.uk/10.3390/v10020071
Submission received: 7 January 2018
/
Revised: 2 February 2018
/
Accepted: 6 February 2018
/
Published: 9 February 2018
(This article belongs to the Special Issue Advances in Alphavirus Research)
Abstract
:Alphaviruses are enveloped, positive single-stranded RNA viruses, typically transmitted by arthropods. They often cause arthralgia or encephalitic diseases in infected humans and there is currently no targeted antiviral treatment available. The re-emergence of alphaviruses in Asia, Europe, and the Americas over the last decade, including chikungunya and o’nyong’nyong viruses, have intensified the search for selective inhibitors. In this review, we highlight key molecular determinants within the alphavirus replication complex that have been identified as viral targets, focusing on their structure and functionality in viral dissemination. We also summarize recent structural data of these viral targets and discuss how these could serve as templates to facilitate structure-based drug design and development of small molecule inhibitors.
1. Introduction
Alphaviruses belong to the Togaviridae family [1]. They are arboviruses that are transmitted to humans through the mosquito species Aedes aegypti and Aedes albopictus, where they cause various diseases and can be broadly divided into arthritogenic [2] and encephalitogenic [3]. Old World alphaviruses such as chikungunya virus (CHIKV), o’nyong’nyong virus (ONNV), and sindbis virus (SINV) are arthritogenic and commonly cause febrile illness accompanied by rash, polyarthralgia and chronic arthritis [4,5,6,7,8]. Infection with New World alphaviruses such as Eastern equine encephalitis (EEEV) and Venezuelan equine encephalitis virus (VEEV), which are less prevalent in humans, are mostly associated with neurological disease [9,10]. In 2005, the re-emergence of CHIKV in several Indian Ocean islands led to massive outbreaks, affecting one-third of the inhabitants in La-Réunion alone [11]. To date, millions of people have been infected in more than 40 countries in Asia, Europe and the Americas [12]. Despite the medical threat posed by CHIKV, there is currently no approved antiviral treatment or vaccine for CHIKV infection, as well as for the other less prevalent alphaviruses. Treatments are usually symptomatic, with administration of non-steroidal anti-inflammatory drugs and analgesic agents to control fever and severe joint pains.
Molecular Virology and Genome Organization
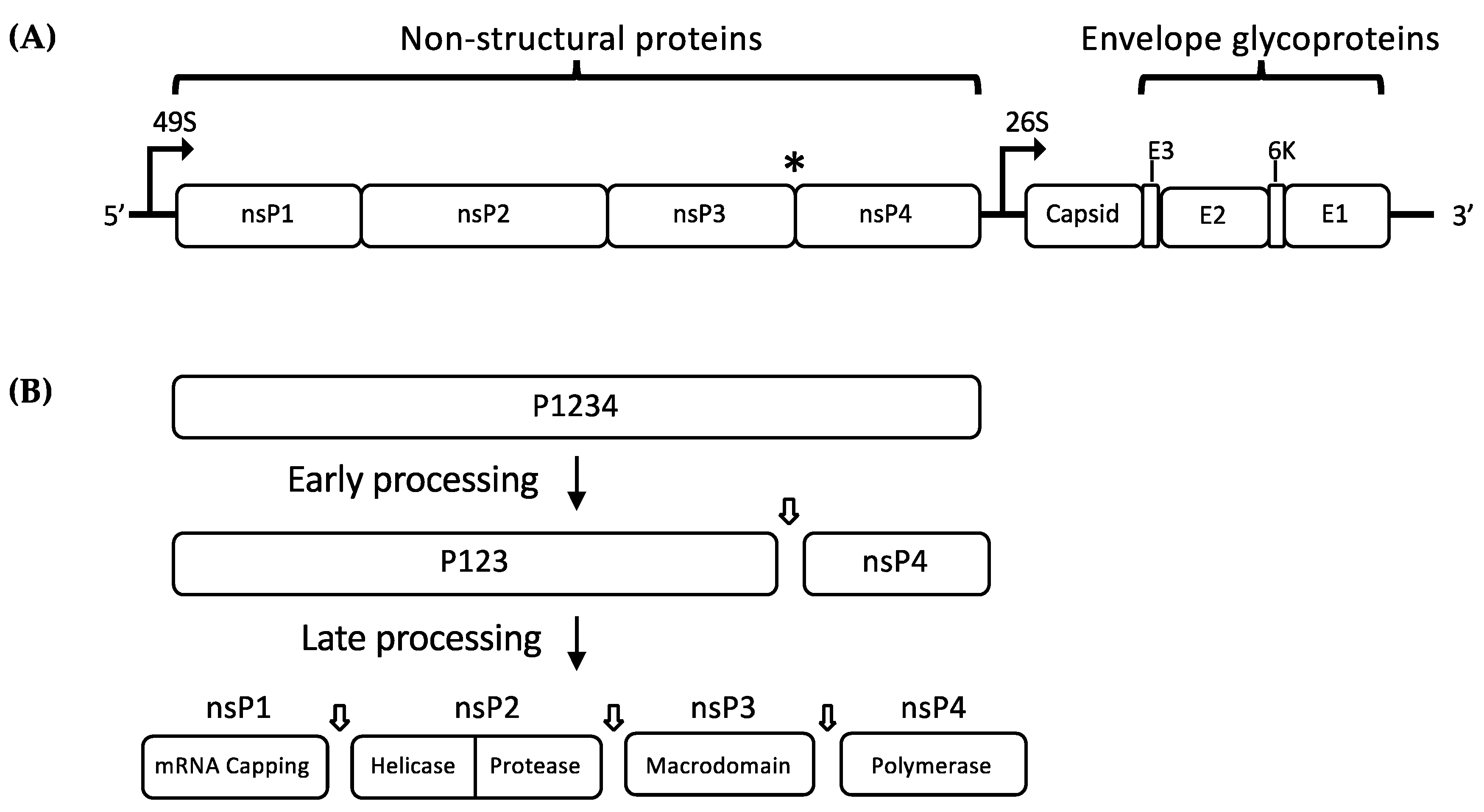
Alphaviruses are small, enveloped RNA viruses with a single-stranded, positive-sense RNA genome. The alphavirus genome is approximately 12 kb long and it consists of two open reading frames (ORFs): a 7 kb frame encoding the non-structural polyprotein and a 4 kb frame encoding the structural polyprotein (Figure 1A) [1,13]. The non-structural polyprotein is cleaved into four different proteins (nsP1, nsP2, nsP3, and nsP4) which are necessary for the transcription and translation of viral mRNA inside the cytoplasm of host cells. Figure 1B describes the stages of nonstructural polyprotein processing. Translation of the viral RNA by host cell translational machinery produces two non-structural protein (nsP) precursors (P123 or P1234). P1234 is expressed as a read-through of an opal termination codon at the end of nsP3 (Figure 1A) [14]. These precursor polyproteins are cleaved by a carboxyl-terminal protease domain of nsP2 [15]. Following translation of P1234, cleavage at the P3/4 junction occurs either in cis or trans, followed by the P1/2 junction which occurs in cis only [16]. Both P123+nsP4 and nsP1+P23+nsP4, together with some cellular proteins, form the early replication complex (RC), which preferentially synthesize negative strand viral RNA [17,18]. The final cleavage event at the P2/3 junction produces fully mature nsPs, which along with host cell proteins, forms the positive strand RC, switching RNA template for synthesis of positive-sense genomic (49S) and subgenomic (26S) RNAs. However, the correlation between P23 cleavage and the switch from negative- to positive-sense RNA production remains poorly understood.
The translation of 26S subgenomic positive sense RNA generates a single structural polyprotein, which is cleaved into five structural proteins: the Capsid (C), two major envelope glycoproteins E1 and E2, and two small cleavage products (E3, 6K) (Figure 1A). While the C protein is being autocatalytically cleaved off to encapsidate new positive sense RNA molecules, the envelope polyprotein precursor E3-E2-6k-E1 is translocated to the endoplasmic reticulum (ER). Host signalases process the polyprotein at the N- and C-terminal end of the 6k peptide, resulting in E3E2, 6k, and E1, where all anchored to the ER membrane. During export to the plasma membrane, the E3E2 precursor is cleaved by furin-like protease activity in the trans-Golgi system into E2 and E3 [19]. The nucleocapsid forms with the assembly of 120 dimers of the C protein, which buds at the cell membrane as spherical particles, acquiring a lipid envelope with embedded E1 and E2 glycoproteins [20,21]. Viral particles exhibit 80 trimeric spikes composed of heterodimers of E1 and E2, with E2 glycoprotein facilitating binding of the virus to cell surface receptors [22,23,24,25]. Upon receptor binding, the virus particle enters the host cell via clathrin-dependent endocytosis [26]. The acidic environment of the endosome results in the E1-mediated fusion of the viral envelope and endosomal membrane, followed by the release of nucleocapsid and uncoating of the viral genome in the cytosol [27,28].
In this review, the organization of alphavirus RC focusing on the function and structure of nsP2 protease will be illustrated. These proteins play important roles in the various replication stages of the viral genome. Notably, the conserved architecture of the nsP2 protease across the New and Old World alphaviruses, as determined by the recently solved structures of VEEV, SINV and CHIKV nsP2 proteases, also makes it an ideal target for designing specific and pan-alphavirus protease inhibitors [29,30,31,32].
2. Roles and Function of Non-Structural Proteins
2.1. Non-Structural Proteins (nsPs)
Key advances have been made to understand the biological aspects and pathogenesis of alphaviruses, using mainly Semliki Forest virus (SFV) and SINV as prototypes [33,34]. Many of the functions of the nsPs have been characterized on the basis of sequence comparisons and biochemical assays (Table 1), and evidence suggests their intrinsic interactions are essential for the formation of functional RCs [18,35,36,37].
2.1.1. Non-Structural Protein 1 (nsP1)
The nsP1 protein is an mRNA capping enzyme that possesses both guanine-7-methyltransferase (MTase) and guanylyltransferase (GTase) activities, where they direct the methylation and capping of newly synthesized viral genomic and subgenomic RNAs [38,39,40,41]. The MTase motif in the N-terminal domain of nsP1 catalyzes the transfer of the methyl group from S-adenosylmethionine (AdoMet) to the N7 position of a GTP molecule (m7Gppp). GTase then binds the m7Gppp, forming a covalent link with a catalytic histidine (m7Gp–GTase) and releasing PPi. The GTase then transfers the m7Gp molecule to the 5′-diphosphate RNA to create m7GpppNp-RNA [42]. The resulting cap structure is essential for viral mRNA translation and prevents the mRNA from being degraded by cellular 5′ exonucleases. Following the N-terminal domain are features that allow the association of the nsP1 protein to cellular membranes. The presence of α-helical amphipathic loop and palmitoylation sites allow the nsP1 protein and nsP1-containing RC to anchor onto the plasma membrane, possibly through nsP1 interaction with the membrane’s anionic phospholipids [43,44,45,46,47].
2.1.2. Non-Structural Protein 2 (nsP2)
The nsP2 protein possesses numerous enzymatic activities and functional roles. The N-terminal region contains a helicase domain that has seven signature motif of Superfamily 1 (SF1) helicases [48]. It functions as an RNA triphosphatase that performs the first of the viral RNA capping reactions [49,50]. It also functions as a nucleotide triphosphatase (NTPase), fueling the RNA helicase activity [50,51]. The C-terminal region of nsP2 contains a papain-like cysteine protease, which is responsible for processing the viral non-structural polyprotein (Figure 2A) [52,53,54]. The protease recognizes conserved motifs within the polyprotein (Figure 2B) [55]. This proteolytic function is highly regulated and is modulated by other domains of nsP2 [16]. The crystal structure has revealed two distinct domains. The N-terminal subdomain has a α/β-fold that is novel, unlike the structure of other known cysteine proteases. The C-terminal subdomain is an S-adenosyl-l-methionine-dependent (SAM) RNA methyltrasferase domain with a classical MTase fold, but enzymatically non-functional [29,30]. Recent crystal structure of VEEV’s enzyme surface has revealed that the predicted active site is in a major surface groove, which is likely to accommodate the substrate polyprotein to be cleaved [29,30]. The major enzyme groove may act as an enzyme mouth holding the protein to be processed. The alphavirus nsP2 protein has also been described as a virulence factor responsible for the transcriptional and translational shutoff in infected host cells and the inhibition of interferon (IFN) mediated antiviral responses contributing to the controlling of translational machinery by viral factors [56,57,58,59].
2.1.3. Non-Structural Protein 3 (nsP3)
The precise role(s) of alphavirus nsP3 protein in the RC is less clear. The nsP3 protein has three recognized domains: the N-terminal macrodomain with phosphatase activity and nucleic acid binding ability, the alphavirus unique domain (AUD) and the C-terminal hypervariable domain [60]. The crystal structures of the macrodomain from CHIKV and VEEV were found to closely resemble the homologous Escherichia coli domain [60]. To date, the most well-defined structural information available are the protease region of nsP2 and the folded N-terminal region of nsP3 (Table 1). It has been solved to 2.85 Å resolution and has a zinc coordination site within the AUD [32]. The hypervariable domain has sequence features of natively unfolded proteins. It has been demonstrated that the deletion of this domain in SFV nsP3 resulted in low viral pathogenicity, suggesting its importance in viral RNA transcription regulation [61].
2.1.4. Non-Structural Protein 4 (nsP4)
The nsP4 polymerase is the most highly conserved protein in alphaviruses, with the most divergent being >50% identical in amino acid sequence when compared with other alphaviral nsP4s [62,63]. The nsP4 contains the core RNA-dependent RNA polymerase (RdRp) domain at the C-terminal end, determined to be solely responsible for the RNA synthetic properties of the viral RC [64]. The RdRp participates in replicating the genomic RNA via a negative strand RNA and transcribing the 26S subgenomic RNA. The N-terminal domain is alphavirus-specific and may be partially disordered structurally. It appears to be important for the interaction with polyprotein P123 to form RCs that are capable of synthesizing minus strands from plus-strand templates [64,65,66]. Deletion and mutation studies of the RdRp domain of SINV nsP4 demonstrated terminal adenylyltransferase (TATase) activity, suggesting it has a potential role in maintenance of the 3′ poly-A tail at the 3′-end of positive-sense RNAs [66]. Comparison of the secondary structure of CHIKV RdRp with the structures of picornaviral polymerases showed a classical, basic RdRp architecture with well-defined fingers, palm containing the GDD active site and thumb domains [67]. A comprehensive review of the nsP4 was recently published that illustrated the fundamental functions of nsP4. It detailed the importance of P123 interaction with nsP4, the importance of RCs’ association with the cellular membrane and the possible interactions of RCs with host factors during viral replication [68].
2.2. Viral Target Proteins for Drug Development
Alphavirus replication is a delicate process, requiring specific protein-protein interactions among the nsPs and host factors to effectively form the highly organized RCs at early and late stages of infection. During infection, the formation of cytoplasmic vacuoles is induced in host cells. These vacuoles contain small cell membrane invaginations called spherules where the RC proteins nsP1 to nsP4, host factors as well as newly synthesized viral RNA localize [69,70,71,72,73]. These structures serve as compartments to facilitate virus propagation, by allowing spatial separation and regulation of RNA translation, replication and packaging of the viral genome. They protect viral RCs and genomic RNA from degradation by cellular proteases and prevent recognition by antiviral double-stranded RNA sensors, such as RIG-I and MDA-5 [74,75,76,77]. Although no 3-D reconstruction of alphavirus replication compartments has been published yet, there are multiple reports revealing the morphologies through electron tomography of other positive sense RNA viruses known to form similar replication sites [78,79,80,81]. Alphavirus RCs are widely accepted to reside on the invaginated cell membrane, with RNA replication taking place in the spherule lumen [70,82,83]. In recent years, attempts have been made to shed light into the late RC’s organization, with a number of studies adopting systems such as yeast two-hybrid [84,85], immunoprecipitation [86,87,88,89] and ELISA [85] to map the interactions among the nsPs. Six novel interactions are identified in CHIKV (nsP1-nsP1, nsP1-nsP2, nsP1-nsP3, nsP1-nsP4, nsP2-nsP4 and nsP4-nsP4) [84], some of which are similarly shown in SINV [88,89,90] and SFV [86,87]. It has also been demonstrated that nsP1 is involved in the recruitment of other nsPs into the spherules [87] and it’s membrane association is crucial for SFV replication [46]. These data suggest that nsP1′s interaction with all other nsPs is absolutely essential to keep the RC intact and functional. Therefore, nsP1 is an attractive target for drug development. Firstly, perturbing its affinity for the cell membrane could potentially inhibit it from anchoring in the spherules. This will eventually stop the recruitment of the other nsPs, thus preventing the initiation of RC formation. Secondly, disrupting the intrinsic interactions among the nsPs could prevent proper conformational arrangements of the RCs and this could ultimately impede viral replication. However, there is an information deficit in the understanding of nsP interactions and without this knowledge, targeted inhibition on RCs remains difficult. Future work will still need to address the relative importance of the arrangements between the nsPs in the RCs.
nsP2 protein is another excellent viral target for inhibition due to its role in viral replication and host evasion strategies [91]. nsP2 is multifunctional: it has RNA helicase, RNA triphosphatase, nucleoside triphosphatase and auto-protease activities. It is often described as an important co-factor for the maturation of viral RC [16,37,92]. The nsP2 protease function is especially of interest, as proteases of other viruses (such as Human Immunodeficiency Virus (HIV) and Hepatitis C Virus (HCV)) have been successfully targeted, leading to the development of FDA-approved inhibitors [93,94,95]. Although the sequence identity of the different alphavirus nsP2 proteases is very low, their active site residues are conserved [29]. Cys478 and His548 of CHIKV nsP2 protease (referred as Cys1013 and His1083 in [31]) are two residues that form the catalytic dyad (Figure 2A), where substrates with defined recognition sequences are cleaved (Figure 2B) [1,31,96,97,98]. It has been demonstrated that a Cys478 to Ala mutation produces inactive protease and completely abolishes CHIKV replication [98]. Recently, the first structure of a peptide-like E64d inhibitor-bound VEEV nsP2 protease (PDB entry 5EZS) was reported (Table 2) [30]. Although E64d is not a viable therapeutic candidate as it could only inhibit protease function and was ineffective in inhibiting viral replication, the structure provided invaluable insights into the roles of the catalytic residues and possible orientation of the substrates during catalysis. The inhibitor was shown to bind beneath a β-hairpin in the interface of the protease and SAM MTase domains. This further demonstrated that the SAM MTase domain is required for proteolysis, with at least three of its residues (Arg662, Lys705 and Lys706) being used by the cysteine protease for substrate binding and recognition. His546 (referred as His548 in CHIKV nsP2) in the protease domain is the only residue adopting a different conformation, with minimal overall structural changes observed when compared to the free enzyme. It was also suggested that the interaction of the carbonyl oxygen of the ester on E64d with the NH of Cys477 (referred as His478 in CHIKV nsP2) stabilized the transition state of the proteolytic reaction. These observations are crucial information as they emphasize the importance of residues within and around the active site cleft. Therefore, targeting these residues would be an applicable strategy to inhibit the enzyme function, which could consequentially inhibit viral replication. Moreover, nsP2 is also identified as a virulence factor. nsP2 plays a role in shutting off host cell mRNA transcription and/or translation [56,59]. A portion of the nsP2 is localized to the nucleus [99,100] and it inhibits host antiviral response by suppressing type I/II interferon-stimulated JAK/STAT signaling [101,102]. A recent study showed that mutations in the nuclear localization signal (NLS) of SFV nsP2 (649DDR651 and 649RDD651) completely blocked nsP2 from entering the nucleus and reduced SFV-induced cell death [103]. This was likely due to retention of nsP2 in the cytoplasm that prevented its association with host factors in the nucleus to shut off host antiviral functions. The NLS of nsP2 could be a good inhibitory target for preventing nsP2 translocation into the nucleus. However, this will only be effective against specific alphaviruses that use nsP2 to inhibit cellular mRNA transcription as it has been demonstrated otherwise for VEEV nsP2 [59].
3. Chemotherapeutics Targeting Viral Non-Structural Proteins
3.1. Antivirals against nsP2 Protease
The reemergence of CHIKV and its subsequent epidemics exemplify the threat alphaviruses currently have on human health. A comprehensive review of the antiviral efforts towards alphaviruses was recently published, where the various types of inhibitors discovered and developed as potential therapeutics were described [104]. It is of prime importance that resolved structures are employed in facilitating the discovery of novel drug compounds. The nsP2 protease’s function as a nonstructural polyprotein processor is absolutely essential for virus replication. Thus, inhibiting this function could potentially mitigate CHIKV infection. This strategy has successfully been used against several viruses like HCV and HIV and has led to the development of remarkably potent drugs [95,105,106,107]. With the recent availability of various alphavirus nsP2 protease crystal structures, they have become the most targeted viral domain for in silico drug design to date. Furthermore, crystal structures of the VEEV, CHIKV and SINV nsP2 protease domain Protein Data Bank (PDB) entries 2HWK [29], 5EZQ [30], 3TRK [31] and 4GUA [32] respectively] displayed highly conserved folds despite having low sequence identity, rendering it a potential target both for specific and pan-alphavirus protease inhibition. Bassetto et al. have effectively employed homology modeling and computer-aided drug design strategies to discover the first few potential compounds against CHIKV nsP2 protease, with the support of a combination of cell-based virus inhibition and cell-free protease assays [108]. Through the virtual screening of about five million commercially available compounds on the VEEV nsP2 protease structure, several were shown to selectively inhibit CHIKV-induced cell death. Compound 25, in particular, displayed the best antiviral activity (EC50 = 3.2 μM) and is predicted to target the central portion of the nsP2 protease active site (Table 2) [108]. Continuing from one of the other lead compounds by Bassetto et al., Das et al. described a set of related compounds, with a few capable of inhibiting CHIKV nsP2 protease as well as virus replication [109]. These results further prove that these compounds derived from in silico drug design are indeed nsP2 protease inhibitors. Many more potential inhibitors originating from the structure-activity relationship and molecular dynamics simulation strategies have been reported and illustrated to dock in the protease’s active site with good affinity [110,111,112]. However, the biological validation of their antiviral activity and/or target specificity is lacking. Aside from screening commercially available compound libraries, repurposing of FDA-approved protease inhibitors is another strategy that could accelerate the identification and development of specific and potent CHIKV protease inhibitors. Lopinavir and Nelfinavir, both potent HIV protease inhibitors, were validated to have modest antiviral activity on CHIKV (Table 2) (EC50 = 32 μM and 14 μM respectively). However, they are clearly toxic on the host cell as they display poor selectivity index values (CC50 = 44 μM and 22 μM respectively) [113]. Nonetheless, these inhibitors could serve as a stepping stone for the development of novel alphavirus protease inhibitors.
3.2. Inhibitors of Other nsPs
Inhibitors targeting other nsPs have also been reported, where their mechanisms of action on alphavirus replication were evaluated through reverse genetic mutation of drug-resistant mutants. A number of highly selective CHIKV and VEEV nsP1 capping enzyme inhibitors postulated to disrupt the GTase activity of nsP1 were reported recently [114,115,116]. MADTP-372 was demonstrated to be potent and selective in inhibiting the induction of cytopathic effect by CHIKV and VEEV [116]. Due to the close proximity of Pro34 residue (mutated in MADTP-372-resistant variant) to His37 residue (a putative acceptor for the guanylylation), it was postulated that MADTP-372 inhibited the GTase reaction by either blocking the binding of m7Gppp to nsP1 or the guanylylation process itself. However, due to the absence of structural information of alphavirus nsP1, other possible scenarios that involve indirect effects cannot be ruled out. Nucleoside analogs have also been proven to be effective against several alphaviruses. Notably, Ribavirin (a guanosine analog widely used against a number of other RNA viruses) demonstrated effective inhibition of CHIKV and SFV genome replication by depleting GTP pools [117,118,119]. The resulting GTP deficiency in host cells could prevent proper capping of newly synthesized viral mRNA by nsP1, and consequently allow cellular 5′ exonucleases to degrade the uncapped mRNA. The absence of the cap structure could also impede viral mRNA translation. In addition, Ribavirin has been proposed to directly inhibit nsP4 RdRp through its interaction with Cys483 residue, resulting in an increase in replication fidelity [120,121]. Another modified nucleoside analog, β-d-N4-hydroxycytidine (NHC), was effective in reducing the rates of VEEV release and its infectivity [122]. NHC is by far the most potent nsP4 inhibitor ever reported (EC50 = 0.426 µM, CC50 > 200 µM) and could potentially be a substitute for Ribavirin as it is inefficient in developing NHC resistant mutants. Another well-studied nsP4 inhibitor, Favipiravir (T-705; a purine analog approved in Japan for the treatment of influenza infections), was shown to exert broad-spectrum anti-alphavirus activity in vitro and provide protection in a mouse model of lethal CHIKV infection [123]. Favipiravir selectively inhibits CHIKV nsP4 RdRp function through its interaction with Lys291 residue. Interestingly, this particular lysine residue is conserved in the polymerases of other positive-sense RNA viruses. Thus this may provide an explanation for Favipiravir’s broad-spectrum antiviral activity. Through the screening of chemical compound libraries, Compound-A was also recently found to inhibit CHIKV infection. Compound-A was demonstrated as a specific nsP4 RdRp inhibitor that could potentially inhibit RdRp’s ribonucleotide selection function by targeting Met2295 residue [124]. It demonstrated potent CHIKV antiviral activity but exhibited very poor selectivity (EC50 = 1.29 µM, CC50 = 5.84 µM). Nonetheless, Compound-A could serve as a starting point for chemical modifications so as to reduce its toxicity.
4. Concluding Remarks and Perspectives
The current understanding of interactions through structural evaluation of the nsP2 protease domain has opened avenues for the development of specific inhibitors. It is by far the most well-defined out of the other nsP domains and its structural information has been inspirational for computational biologists and chemists alike in designing many series of compounds, mainly targeting its catalytic binding pocket. The representation of E64d inhibitor bound to the nsP2 catalytic pocket is a major advancement towards identifying the key residues that may be important for substrate binding and recognition [30]. However, the lack of structural information on the other viral nsPs makes understanding of the mode of action of inhibitors and rational designing of specific and efficient inhibitors challenging. It is worthwhile to note that for all nsP1 and nsP4 inhibitors discussed earlier, the direct interactions with their targeted residues were never demonstrated. In particular, obtaining the crystal structure of alphavirus nsP4 RdRp would allow more robust comparison of this polymerase to other viral RdRps and new relationships between RdRp-encoding viruses could be formed. Resolving the structure of the RC will also allow for a better understanding of the function of the polymerase and the entire RC.
Acknowledgments
This work is supported by core research grants provided to the Singapore Immunology Network by the Biomedical Research Council (BMRC), Agency for Science, Technology and Research (A*STAR), Singapore. We would also like to thank Kai-Er Eng from SIgN, A*STAR for critical discussion and proofreading of the manuscript.
Author Contributions
Farhana Abu Bakar and Lisa F. P. Ng wrote the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Strauss, J.H.; Strauss, E.G. The alphaviruses: Gene expression, replication, and evolution. Microbiol. Rev. 1994, 58, 491–562. [Google Scholar] [PubMed]
- Laine, M.; Luukkainen, R.; Toivanen, A. Sindbis viruses and other alphaviruses as cause of human arthritic disease. J. Intern. Med. 2004, 256, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Steele, K.E.; Twenhafel, N.A. Review paper: Pathology of animal models of alphavirus encephalitis. Vet. Pathol. 2010, 47, 790–805. [Google Scholar] [CrossRef] [PubMed]
- Borgherini, G.; Poubeau, P.; Staikowsky, F.; Lory, M.; Le Moullec, N.; Becquart, J.P.; Wengling, C.; Michault, A.; Paganin, F. Outbreak of chikungunya on Reunion Island: Early clinical and laboratory features in 157 adult patients. Clin. Infect. Dis. 2007, 44, 1401–1407. [Google Scholar] [CrossRef] [PubMed]
- Simon, F.; Parola, P.; Grandadam, M.; Fourcade, S.; Oliver, M.; Brouqui, P.; Hance, P.; Kraemer, P.; Ali Mohamed, A.; de Lamballerie, X.; et al. Chikungunya infection: An emerging rheumatism among travelers returned from Indian Ocean islands. Report of 47 cases. Medicine 2007, 86, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Pialoux, G.; Gauzere, B.A.; Jaureguiberry, S.; Strobel, M. Chikungunya, an epidemic arbovirosis. Lancet Infect. Dis. 2007, 7, 319–327. [Google Scholar] [CrossRef]
- Manimunda, S.P.; Vijayachari, P.; Uppoor, R.; Sugunan, A.P.; Singh, S.S.; Rai, S.K.; Sudeep, A.B.; Muruganandam, N.; Chaitanya, I.K.; Guruprasad, D.R. Clinical progression of chikungunya fever during acute and chronic arthritic stages and the changes in joint morphology as revealed by imaging. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Ali Ou Alla, S.; Combe, B. Arthritis after infection with Chikungunya virus. Best Prac. Res. Clin. Rheumatol. 2011, 25, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Holbrook, M.R.; Gowen, B.B. Animal models of highly pathogenic RNA viral infections: Encephalitis viruses. Antivir. Res. 2008, 78, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Zacks, M.A.; Paessler, S. Encephalitic alphaviruses. Vet. Microbiol. 2010, 140, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Renault, P.; Solet, J.L.; Sissoko, D.; Balleydier, E.; Larrieu, S.; Filleul, L.; Lassalle, C.; Thiria, J.; Rachou, E.; de Valk, H.; et al. A major epidemic of chikungunya virus infection on Reunion Island, France, 2005–2006. Am. J. Trop. Med. Hyg. 2007, 77, 727–731. [Google Scholar] [PubMed]
- World Health Organization. Chikungunya—Fact Sheet; World Health Organization: Geneva, Switzerland, 2017; pp. 1–5. [Google Scholar]
- Strauss, E.G.; Rice, C.M.; Strauss, J.H. Complete nucleotide sequence of the genomic RNA of Sindbis virus. Virology 1984, 133, 92–110. [Google Scholar] [CrossRef]
- Strauss, E.G.; Rice, C.M.; Strauss, J.H. Sequence coding for the alphavirus nonstructural proteins is interrupted by an opal termination codon. Proc. Natl. Acad. Sci. USA 1983, 80, 5271–5275. [Google Scholar] [CrossRef] [PubMed]
- De Groot, R.J.; Hardy, W.R.; Shirako, Y.; Strauss, J.H. Cleavage-site preferences of Sindbis virus polyproteins containing the non-structural proteinase. Evidence for temporal regulation of polyprotein processing in vivo. EMBO J. 1990, 9, 2631–2638. [Google Scholar] [PubMed]
- Vasiljeva, L.; Merits, A.; Golubtsov, A.; Sizemskaja, V.; Kaariainen, L.; Ahola, T. Regulation of the sequential processing of Semliki Forest virus replicase polyprotein. J. Biol. Chem. 2003, 278, 41636–41645. [Google Scholar] [CrossRef] [PubMed]
- Barton, D.J.; Sawicki, S.G.; Sawicki, D.L. Solubilization and immunoprecipitation of alphavirus replication complexes. J. Virol. 1991, 65, 1496–1506. [Google Scholar] [PubMed]
- Shirako, Y.; Strauss, J.H. Regulation of Sindbis virus RNA replication: Uncleaved P123 and nsP4 function in minus-strand RNA synthesis, whereas cleaved products from P123 are required for efficient plus-strand RNA synthesis. J. Virol. 1994, 68, 1874–1885. [Google Scholar] [PubMed]
- Tang, B.L. The cell biology of Chikungunya virus infection. Cell. Microbiol. 2012, 14, 1354–1363. [Google Scholar] [CrossRef] [PubMed]
- Perera, R.; Owen, K.E.; Tellinghuisen, T.L.; Gorbalenya, A.E.; Kuhn, R.J. Alphavirus nucleocapsid protein contains a putative coiled coil alpha-helix important for core assembly. J. Virol. 2001, 75, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ekstrom, M.; Liljestrom, P.; Garoff, H. Membrane protein lateral interactions control Semliki Forest virus budding. EMBO J. 1994, 13, 1058–1064. [Google Scholar] [PubMed]
- Paredes, A.M.; Brown, D.T.; Rothnagel, R.; Chiu, W.; Schoepp, R.J.; Johnston, R.E.; Prasad, B.V. Three-dimensional structure of a membrane-containing virus. Proc. Natl. Acad. Sci. USA 1993, 90, 9095–9099. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.H.; Kuhn, R.J.; Olson, N.H.; Rossmann, M.G.; Choi, H.K.; Smith, T.J.; Baker, T.S. Nucleocapsid and glycoprotein organization in an enveloped virus. Cell 1995, 80, 621–630. [Google Scholar] [CrossRef]
- Mancini, E.J.; Clarke, M.; Gowen, B.E.; Rutten, T.; Fuller, S.D. Cryo-electron microscopy reveals the functional organization of an enveloped virus, Semliki Forest virus. Mol. Cell 2000, 5, 255–266. [Google Scholar] [CrossRef]
- Lescar, J.; Roussel, A.; Wien, M.W.; Navaza, J.; Fuller, S.D.; Wengler, G.; Wengler, G.; Rey, F.A. The Fusion glycoprotein shell of Semliki Forest virus: An icosahedral assembly primed for fusogenic activation at endosomal pH. Cell 2001, 105, 137–148. [Google Scholar] [CrossRef]
- Sourisseau, M.; Schilte, C.; Casartelli, N.; Trouillet, C.; Guivel-Benhassine, F.; Rudnicka, D.; Sol-Foulon, N.; Le Roux, K.; Prevost, M.C.; Fsihi, H.; et al. Characterization of reemerging chikungunya virus. PLoS Pathog. 2007, 3, e89. [Google Scholar] [CrossRef] [PubMed]
- Kielian, M.; Rey, F.A. Virus membrane-fusion proteins: More than one way to make a hairpin. Nat. Rev. Microbiol. 2006, 4, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Marsh, M.; Helenius, A. Virus entry: Open sesame. Cell 2006, 124, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.T.; White, M.A.; Watowich, S.J. The crystal structure of the Venezuelan equine encephalitis alphavirus nsP2 protease. Structure 2006, 14, 1449–1458. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Compton, J.R.; Leary, D.H.; Olson, M.A.; Lee, M.S.; Cheung, J.; Ye, W.; Ferrer, M.; Southall, N.; Jadhav, A.; et al. Kinetic, Mutational, and Structural Studies of the Venezuelan Equine Encephalitis Virus Nonstructural Protein 2 Cysteine Protease. Biochemistry 2016, 55, 3007–3019. [Google Scholar] [CrossRef] [PubMed]
- Cheung, J.; Franklin, M.; Mancia, F.; Rudolph, M.; Cassidy, M.; Gary, E.; Burshteyn, F.; Love, J. Structure of the Chikungunya virus nsP2 protease. Unpublished work. 2011. [Google Scholar]
- Shin, G.; Yost, S.A.; Miller, M.T.; Elrod, E.J.; Grakoui, A.; Marcotrigiano, J. Structural and functional insights into alphavirus polyprotein processing and pathogenesis. Proc. Natl. Acad. Sci. USA 2012, 109, 16534–16539. [Google Scholar] [CrossRef] [PubMed]
- Atkins, G.J.; Sheahan, B.J.; Liljestrom, P. The molecular pathogenesis of Semliki Forest virus: A model virus made useful? J. Gen. Virol. 1999, 80 Pt 9, 2287–2297. [Google Scholar] [CrossRef] [PubMed]
- Rupp, J.C.; Sokoloski, K.J.; Gebhart, N.N.; Hardy, R.W. Alphavirus RNA synthesis and non-structural protein functions. J. Gen. Virol. 2015, 96, 2483–2500. [Google Scholar] [CrossRef] [PubMed]
- Gorchakov, R.; Frolova, E.; Sawicki, S.; Atasheva, S.; Sawicki, D.; Frolov, I. A new role for ns polyprotein cleavage in Sindbis virus replication. J. Virol. 2008, 82, 6218–6231. [Google Scholar] [CrossRef] [PubMed]
- Lemm, J.A.; Rice, C.M. Assembly of functional Sindbis virus RNA replication complexes: Requirement for coexpression of P123 and P34. J. Virol. 1993, 67, 1905–1915. [Google Scholar] [PubMed]
- Lemm, J.A.; Rumenapf, T.; Strauss, E.G.; Strauss, J.H.; Rice, C.M. Polypeptide requirements for assembly of functional Sindbis virus replication complexes: A model for the temporal regulation of minus- and plus-strand RNA synthesis. EMBO J. 1994, 13, 2925–2934. [Google Scholar] [PubMed]
- Cross, R.K. Identification of a unique guanine-7-methyltransferase in Semliki Forest virus (SFV) infected cell extracts. Virology 1983, 130, 452–463. [Google Scholar] [CrossRef]
- Laakkonen, P.; Hyvönen, M.; Peränen, J.; Kääriäinen, L. Expression of Semliki Forest virus nsP1-specific methyltransferase in insect cells and in Escherichia coli. J. Virol. 1994, 68, 7418–7425. [Google Scholar] [PubMed]
- Ahola, T.; Kääriäinen, L. Reaction in alphavirus mRNA capping: Formation of a covalent complex of nonstructural protein nsP1 with 7-methyl-GMP. Proc. Natl. Acad. Sci. USA 1995, 92, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Mi, S.; Stollar, V. Expression of Sindbis virus nsP1 and methyltransferase activity in Escherichia coli. Virology 1991, 184, 423–427. [Google Scholar] [CrossRef]
- Decroly, E.; Ferron, F.; Lescar, J.; Canard, B. Conventional and unconventional mechanisms for capping viral mRNA. Nat. Rev. Microbiol. 2011, 10, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Laakkonen, P.; Ahola, T.; Kaariainen, L. The effects of palmitoylation on membrane association of Semliki forest virus RNA capping enzyme. J. Biol. Chem. 1996, 271, 28567–28571. [Google Scholar] [CrossRef] [PubMed]
- Ahola, T.; Lampio, A.; Auvinen, P.; Kaariainen, L. Semliki Forest virus mRNA capping enzyme requires association with anionic membrane phospholipids for activity. EMBO J. 1999, 18, 3164–3172. [Google Scholar] [CrossRef] [PubMed]
- Lampio, A.; Kilpelainen, I.; Pesonen, S.; Karhi, K.; Auvinen, P.; Somerharju, P.; Kaariainen, L. Membrane binding mechanism of an RNA virus-capping enzyme. J. Biol. Chem. 2000, 275, 37853–37859. [Google Scholar] [CrossRef] [PubMed]
- Spuul, P.; Salonen, A.; Merits, A.; Jokitalo, E.; Kaariainen, L.; Ahola, T. Role of the amphipathic peptide of Semliki forest virus replicase protein nsP1 in membrane association and virus replication. J. Virol. 2007, 81, 872–883. [Google Scholar] [CrossRef] [PubMed]
- Ahola, T.; Kujala, P.; Tuittila, M.; Blom, T.; Laakkonen, P.; Hinkkanen, A.; Auvinen, P. Effects of palmitoylation of replicase protein nsP1 on alphavirus infection. J. Virol. 2000, 74, 6725–6733. [Google Scholar] [CrossRef] [PubMed]
- Gorbalenya, A.E.; Koonin, E.V.; Donchenko, A.P.; Blinov, V.M. Two related superfamilies of putative helicases involved in replication, recombination, repair and expression of DNA and RNA genomes. Nucleic Acids Res. 1989, 17, 4713–4730. [Google Scholar] [CrossRef] [PubMed]
- Vasiljeva, L.; Merits, A.; Auvinen, P.; Kaariainen, L. Identification of a novel function of the alphavirus capping apparatus. RNA 5′-triphosphatase activity of nsP2. J. Biol. Chem. 2000, 275, 17281–17287. [Google Scholar] [CrossRef] [PubMed]
- Karpe, Y.A.; Aher, P.P.; Lole, K.S. NTPase and 5′-RNA triphosphatase activities of Chikungunya virus nsP2 protein. PLoS ONE 2011, 6, e22336. [Google Scholar] [CrossRef] [PubMed]
- Rikkonen, M.; Peränen, J.; Kääriäinen, L. ATPase and GTPase activities associated with Semliki Forest virus nonstructural protein nsP2. J. Virol. 1994, 68, 5804–5810. [Google Scholar] [PubMed]
- Kim, K.H.; Rumenapf, T.; Strauss, E.G.; Strauss, J.H. Regulation of Semliki Forest virus RNA replication: A model for the control of alphavirus pathogenesis in invertebrate hosts. Virology 2004, 323, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Vasiljeva, L.; Valmu, L.; Kääriäinen, L.; Merits, A. Site-specific protease activity of the carboxyl-terminal domain of Semliki Forest virus replicase protein nsP2. J. Biol. Chem. 2001, 276, 30786–30793. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, C.; Kutumbarao, N.H.V.; Suhitha, S.; Velmurugan, D. Structure–function relationship of Chikungunya nsP2 protease: A comparative study with papain. Chem. Biol. Drug Des. 2017, 89, 772–782. [Google Scholar] [CrossRef] [PubMed]
- Lulla, A.; Lulla, V.; Tints, K.; Ahola, T.; Merits, A. Molecular Determinants of Substrate Specificity for Semliki Forest Virus Nonstructural Protease. J. Virol. 2006, 80, 5413–5422. [Google Scholar] [CrossRef] [PubMed]
- Gorchakov, R.; Frolova, E.; Frolov, I. Inhibition of transcription and translation in Sindbis virus-infected cells. J. Virol. 2005, 79, 9397–9409. [Google Scholar] [CrossRef] [PubMed]
- Breakwell, L.; Dosenovic, P.; Karlsson Hedestam, G.B.; D’Amato, M.; Liljestrom, P.; Fazakerley, J.; McInerney, G.M. Semliki Forest virus nonstructural protein 2 is involved in suppression of the type I interferon response. J. Virol. 2007, 81, 8677–8684. [Google Scholar] [CrossRef] [PubMed]
- Frolov, I.; Garmashova, N.; Atasheva, S.; Frolova, E.I. Random insertion mutagenesis of sindbis virus nonstructural protein 2 and selection of variants incapable of downregulating cellular transcription. J. Virol. 2009, 83, 9031–9044. [Google Scholar] [CrossRef] [PubMed]
- Bhalla, N.; Sun, C.; Metthew Lam, L.K.; Gardner, C.L.; Ryman, K.D.; Klimstra, W.B. Host translation shutoff mediated by non-structural protein 2 is a critical factor in the antiviral state resistance of Venezuelan equine encephalitis virus. Virology 2016, 496, 147–165. [Google Scholar] [CrossRef] [PubMed]
- Malet, H.; Coutard, B.; Jamal, S.; Dutartre, H.; Papageorgiou, N.; Neuvonen, M.; Ahola, T.; Forrester, N.; Gould, E.A.; Lafitte, D.; et al. The crystal structures of Chikungunya and Venezuelan equine encephalitis virus nsP3 macro domains define a conserved adenosine binding pocket. J. Virol. 2009, 83, 6534–6545. [Google Scholar] [CrossRef] [PubMed]
- Tuittila, M.T.; Santagati, M.G.; Röyttä, M.; Määttä, J.A.; Hinkkanen, A.E. Replicase complex genes of Semliki Forest virus confer lethal neurovirulence. J. Virol. 2000, 74, 4579–4589. [Google Scholar] [CrossRef] [PubMed]
- Forrester, N.L.; Guerbois, M.; Seymour, R.L.; Spratt, H.; Weaver, S.C. Vector-borne transmission imposes a severe bottleneck on an RNA virus population. PLoS Pathog. 2012, 8, e1002897. [Google Scholar] [CrossRef] [PubMed]
- Weston, J.; Villoing, S.; Bremont, M.; Castric, J.; Pfeffer, M.; Jewhurst, V.; McLoughlin, M.; Rodseth, O.; Christie, K.E.; Koumans, J.; et al. Comparison of two aquatic alphaviruses, salmon pancreas disease virus and sleeping disease virus, by using genome sequence analysis, monoclonal reactivity, and cross-infection. J. Virol. 2002, 76, 6155–6163. [Google Scholar] [CrossRef] [PubMed]
- Rubach, J.K.; Wasik, B.R.; Rupp, J.C.; Kuhn, R.J.; Hardy, R.W.; Smith, J.L. Characterization of purified Sindbis virus nsP4 RNA-dependent RNA polymerase activity in vitro. Virology 2009, 384, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Rupp, J.C.; Jundt, N.; Hardy, R.W. Requirement for the Amino-Terminal Domain of Sindbis Virus nsP4 during Virus Infection. J. Virol. 2011, 85, 3449–3460. [Google Scholar] [CrossRef] [PubMed]
- Tomar, S.; Hardy, R.W.; Smith, J.L.; Kuhn, R.J. Catalytic core of alphavirus nonstructural protein nsP4 possesses terminal adenylyltransferase activity. J. Virol. 2006, 80, 9962–9969. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.W.; Tan, Y.B.; Zheng, J.; Zhao, Y.; Lim, B.T.; Cornvik, T.; Lescar, J.; Ng, L.F.P.; Luo, D. Chikungunya virus nsP4 RNA-dependent RNA polymerase core domain displays detergent-sensitive primer extension and terminal adenylyltransferase activities. Antivir. Res. 2017, 143, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Pietila, M.K.; Hellstrom, K.; Ahola, T. Alphavirus polymerase and RNA replication. Virus Res. 2017, 234, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Grimley, P.M.; Berezesky, I.K.; Friedman, R.M. Cytoplasmic structures associated with an arbovirus infection: Loci of viral ribonucleic acid synthesis. J. Virol. 1968, 2, 1326–1338. [Google Scholar] [PubMed]
- Kujala, P.; Ikaheimonen, A.; Ehsani, N.; Vihinen, H.; Auvinen, P.; Kaariainen, L. Biogenesis of the Semliki Forest virus RNA replication complex. J. Virol. 2001, 75, 3873–3884. [Google Scholar] [CrossRef] [PubMed]
- Frolova, E.I.; Gorchakov, R.; Pereboeva, L.; Atasheva, S.; Frolov, I. Functional Sindbis virus replicative complexes are formed at the plasma membrane. J. Virol. 2010, 84, 11679–11695. [Google Scholar] [CrossRef] [PubMed]
- Atasheva, S.; Fish, A.; Fornerod, M.; Frolova, E.I. Venezuelan equine Encephalitis virus capsid protein forms a tetrameric complex with CRM1 and importin α/β that obstructs nuclear pore complex function. J. Virol. 2010, 84, 4158–4171. [Google Scholar] [CrossRef] [PubMed]
- Neuvonen, M.; Kazlauskas, A.; Martikainen, M.; Hinkkanen, A.; Ahola, T.; Saksela, K. SH3 domain-mediated recruitment of host cell amphiphysins by alphavirus nsP3 promotes viral RNA replication. PLoS Pathog. 2011, 7, e1002383. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Hur, S. Helicases in Antiviral Immunity: Dual Properties as Sensors and Effectors. Trends Biochem. Sci. 2015, 40, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Andrejeva, J.; Childs, K.S.; Young, D.F.; Carlos, T.S.; Stock, N.; Goodbourn, S.; Randall, R.E. The V proteins of paramyxoviruses bind the IFN-inducible RNA helicase, mda-5, and inhibit its activation of the IFN-β promoter. Proc. Natl. Acad. Sci. USA 2004, 101, 17264–17269. [Google Scholar] [CrossRef] [PubMed]
- Diao, F.; Li, S.; Tian, Y.; Zhang, M.; Xu, L.G.; Zhang, Y.; Wang, R.P.; Chen, D.; Zhai, Z.; Zhong, B.; et al. Negative regulation of MDA5- but not RIG-I-mediated innate antiviral signaling by the dihydroxyacetone kinase. Proc. Natl. Acad. Sci. USA 2007, 104, 11706–11711. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Kikuchi, M.; Natsukawa, T.; Shinobu, N.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Akira, S.; Fujita, T. The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat. Immunol. 2004, 5, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Ertel, K.J.; Benefield, D.; Castano-Diez, D.; Pennington, J.G.; Horswill, M.; den Boon, J.A.; Otegui, M.S.; Ahlquist, P. Cryo-electron tomography reveals novel features of a viral RNA replication compartment. eLife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Kopek, B.G.; Perkins, G.; Miller, D.J.; Ellisman, M.H.; Ahlquist, P. Three-dimensional analysis of a viral RNA replication complex reveals a virus-induced mini-organelle. PLoS Biol. 2007, 5, e220. [Google Scholar] [CrossRef] [PubMed]
- Fontana, J.; Lopez-Iglesias, C.; Tzeng, W.P.; Frey, T.K.; Fernandez, J.J.; Risco, C. Three-dimensional structure of Rubella virus factories. Virology 2010, 405, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Welsch, S.; Miller, S.; Romero-Brey, I.; Merz, A.; Bleck, C.K.; Walther, P.; Fuller, S.D.; Antony, C.; Krijnse-Locker, J.; Bartenschlager, R. Composition and three-dimensional architecture of the dengue virus replication and assembly sites. Cell Host Microbe 2009, 5, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Froshauer, S.; Kartenbeck, J.; Helenius, A. Alphavirus RNA replicase is located on the cytoplasmic surface of endosomes and lysosomes. J. Cell Biol. 1988, 107, 2075–2086. [Google Scholar] [CrossRef] [PubMed]
- Hellstrom, K.; Kallio, K.; Merilainen, H.M.; Jokitalo, E.; Ahola, T. Ability of minus strands and modified plus strands to act as templates in Semliki Forest virus RNA replication. J. Gen. Virol. 2016, 97, 1395–1407. [Google Scholar] [CrossRef] [PubMed]
- Rana, J.; Rajasekharan, S.; Gulati, S.; Dudha, N.; Gupta, A.; Chaudhary, V.K.; Gupta, S. Network mapping among the functional domains of Chikungunya virus nonstructural proteins. Proteins 2014, 82, 2403–2411. [Google Scholar] [CrossRef] [PubMed]
- Sreejith, R.; Rana, J.; Dudha, N.; Kumar, K.; Gabrani, R.; Sharma, S.K.; Gupta, A.; Vrati, S.; Chaudhary, V.K.; Gupta, S. Mapping interactions of Chikungunya virus nonstructural proteins. Virus Res. 2012, 169, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Lulla, V.; Sawicki, D.L.; Sawicki, S.G.; Lulla, A.; Merits, A.; Ahola, T. Molecular defects caused by temperature-sensitive mutations in Semliki Forest virus nsP1. J. Virol. 2008, 82, 9236–9244. [Google Scholar] [CrossRef] [PubMed]
- Salonen, A.; Vasiljeva, L.; Merits, A.; Magden, J.; Jokitalo, E.; Kaariainen, L. Properly folded nonstructural polyprotein directs the semliki forest virus replication complex to the endosomal compartment. J. Virol. 2003, 77, 1691–1702. [Google Scholar] [CrossRef] [PubMed]
- Shirako, Y.; Strauss, J.H. Requirement for an aromatic amino acid or histidine at the N terminus of Sindbis virus RNA polymerase. J. Virol. 1998, 72, 2310–2315. [Google Scholar] [PubMed]
- Sawicki, D.L.; Sawicki, S.G. A second nonstructural protein functions in the regulation of alphavirus negative-strand RNA synthesis. J. Virol. 1993, 67, 3605–3610. [Google Scholar] [PubMed]
- Fata, C.L.; Sawicki, S.G.; Sawicki, D.L. Modification of Asn374 of nsP1 suppresses a Sindbis virus nsP4 minus-strand polymerase mutant. J. Virol. 2002, 76, 8641–8649. [Google Scholar] [CrossRef] [PubMed]
- Fros, J.J.; Pijlman, G.P. Alphavirus Infection: Host Cell Shut-Off and Inhibition of Antiviral Responses. Viruses 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Lemm, J.A.; Bergqvist, A.; Read, C.M.; Rice, C.M. Template-dependent initiation of Sindbis virus RNA replication in vitro. J. Virol. 1998, 72, 6546–6553. [Google Scholar] [PubMed]
- Manns, M.P.; von Hahn, T. Novel therapies for hepatitis C—One pill fits all? Nat. Rev. Drug Discov. 2013, 12, 595–610. [Google Scholar] [CrossRef] [PubMed]
- Pokorna, J.; Machala, L.; Rezacova, P.; Konvalinka, J. Current and Novel Inhibitors of HIV Protease. Viruses 2009, 1, 1209–1239. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, E. The design of drugs for HIV and HCV. Nat. Rev. Drug Discov. 2007, 6, 1001–1018. [Google Scholar] [CrossRef] [PubMed]
- Strauss, E.G.; de Groot, R.J.; Levinson, R.; Strauss, J.H. Identification of the active site residues in the nsP2 proteinase of Sindbis virus. Virology 1992, 191, 932–940. [Google Scholar] [CrossRef]
- Golubtsov, A.; Kääriäinen, L.; Caldentey, J. Characterization of the cysteine protease domain of Semliki Forest virus replicase protein nsP2 by in vitro mutagenesis. FEBS Lett. 2006, 580, 1502–1508. [Google Scholar] [CrossRef] [PubMed]
- Rausalu, K.; Utt, A.; Quirin, T.; Varghese, F.S.; Zusinaite, E.; Das, P.K.; Ahola, T.; Merits, A. Chikungunya virus infectivity, RNA replication and non-structural polyprotein processing depend on the nsP2 protease’s active site cysteine residue. Sci. Rep. 2016, 6, 37124. [Google Scholar] [CrossRef] [PubMed]
- Rikkonen, M.; Peranen, J.; Kaariainen, L. Nuclear and nucleolar targeting signals of Semliki Forest virus nonstructural protein nsP2. Virology 1992, 189, 462–473. [Google Scholar] [CrossRef]
- Rikkonen, M.; Peranen, J.; Kaariainen, L. Nuclear targeting of Semliki Forest virus nsP2. Arch. Virol. Suppl. 1994, 9, 369–377. [Google Scholar] [PubMed]
- Fros, J.J.; van der Maten, E.; Vlak, J.M.; Pijlman, G.P. The C-terminal domain of chikungunya virus nsP2 independently governs viral RNA replication, cytopathicity, and inhibition of interferon signaling. J. Virol. 2013, 87, 10394–10400. [Google Scholar] [CrossRef] [PubMed]
- Bourai, M.; Lucas-Hourani, M.; Gad, H.H.; Drosten, C.; Jacob, Y.; Tafforeau, L.; Cassonnet, P.; Jones, L.M.; Judith, D.; Couderc, T.; et al. Mapping of Chikungunya virus interactions with host proteins identified nsP2 as a highly connected viral component. J. Virol. 2012, 86, 3121–3134. [Google Scholar] [CrossRef] [PubMed]
- Tamm, K.; Merits, A.; Sarand, I. Mutations in the nuclear localization signal of nsP2 influencing RNA synthesis, protein expression and cytotoxicity of Semliki Forest virus. J. Gen. Virol. 2008, 89, 676–686. [Google Scholar] [CrossRef] [PubMed]
- Ching, K.-C.; Ng, L.F.P.; Chai, C.L.L. A compendium of small molecule direct-acting and host-targeting inhibitors as therapies against alphaviruses. J. Antimicrob. Chemother. 2017, 72, 2973–2989. [Google Scholar] [CrossRef] [PubMed]
- Kwong, A.D.; Kauffman, R.S.; Hurter, P.; Mueller, P. Discovery and development of telaprevir: An NS3-4A protease inhibitor for treating genotype 1 chronic hepatitis C virus. Nat. Biotechnol. 2011, 29, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Howe, A.Y.; Venkatraman, S. The Discovery and Development of Boceprevir: A Novel, First-generation Inhibitor of the Hepatitis C Virus NS3/4A Serine Protease. J. Clin. Transl. Hepatol. 2013, 1, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Chu, Y.; Wang, Y. HIV protease inhibitors: A review of molecular selectivity and toxicity. HIV AIDS 2015, 7, 95–104. [Google Scholar] [CrossRef]
- Bassetto, M.; de Burghgraeve, T.; Delang, L.; Massarotti, A.; Coluccia, A.; Zonta, N.; Gatti, V.; Colombano, G.; Sorba, G.; Silvestri, R.; et al. Computer-aided identification, design and synthesis of a novel series of compounds with selective antiviral activity against chikungunya virus. Antivir. Res. 2013, 98, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Das, P.K.; Puusepp, L.; Varghese, F.S.; Utt, A.; Ahola, T.; Kananovich, D.G.; Lopp, M.; Merits, A.; Karelson, M. Design and Validation of Novel Chikungunya Virus Protease Inhibitors. Antimicrob. Agents Chemother. 2016, 60, 7382–7395. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.D.; Kirubakaran, P.; Nagarajan, S.; Sakkiah, S.; Muthusamy, K.; Velmurgan, D.; Jeyakanthan, J. Homology modeling, molecular dynamics, e-pharmacophore mapping and docking study of Chikungunya virus nsP2 protease. J. Mol. Model. 2012, 18, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.T.; Yu, H.; Keller, P.A. Identification of chikungunya virus nsP2 protease inhibitors using structure-base approaches. J. Mol. Graph. Model. 2015, 57, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Byler, K.G.; Collins, J.T.; Ogungbe, I.V.; Setzer, W.N. Alphavirus protease inhibitors from natural sources: A homology modeling and molecular docking investigation. Comput. Biol. Chem. 2016, 64, 163–184. [Google Scholar] [CrossRef] [PubMed]
- Bhakat, S.; Delang, L.; Kaptein, S.; Neyts, J.; Leyssen, P.; Jayaprakash, V. Reaching beyond HIV/HCV: Nelfinavir as a potential starting point for broad-spectrum protease inhibitors against dengue and chikungunya virus. RSC Adv. 2015, 5, 85938–85949. [Google Scholar] [CrossRef]
- Gigante, A.; Canela, M.D.; Delang, L.; Priego, E.M.; Camarasa, M.J.; Querat, G.; Neyts, J.; Leyssen, P.; Perez-Perez, M.J. Identification of 1,2,3 triazolo[4,5-d]pyrimidin-7(6H)-ones as novel inhibitors of Chikungunya virus replication. J. Med. Chem. 2014, 57, 4000–4008. [Google Scholar] [CrossRef] [PubMed]
- Gigante, A.; Gomez-SanJuan, A.; Delang, L.; Li, C.; Bueno, O.; Gamo, A.M.; Priego, E.M.; Camarasa, M.J.; Jochmans, D.; Leyssen, P.; et al. Antiviral activity of 1,2,3 triazolo[4,5-d]pyrimidin-7(6H)-ones against chikungunya virus targeting the viral capping nsP1. Antivir. Res. 2017, 144, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Delang, L.; Li, C.; Tas, A.; Quérat, G.; Albulescu, I.C.; de Burghgraeve, T.; Guerrero, N.A.S.; Gigante, A.; Piorkowski, G.; Decroly, E.; et al. The viral capping enzyme nsP1: A novel target for the inhibition of chikungunya virus infection. Sci. Rep. 2016, 6, 31819. [Google Scholar] [CrossRef] [PubMed]
- Briolant, S.; Garin, D.; Scaramozzino, N.; Jouan, A.; Crance, J.M. In vitro inhibition of Chikungunya and Semliki Forest viruses replication by antiviral compounds: Synergistic effect of interferon-α and ribavirin combination. Antivir. Res. 2004, 61, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Rothan, H.A.; Bahrani, H.; Mohamed, Z.; Teoh, T.C.; Shankar, E.M.; Rahman, N.A.; Yusof, R. A combination of doxycycline and ribavirin alleviated chikungunya infection. PLoS ONE 2015, 10, e0126360. [Google Scholar] [CrossRef] [PubMed]
- Scholte, F.E.; Tas, A.; Martina, B.E.; Cordioli, P.; Narayanan, K.; Makino, S.; Snijder, E.J.; van Hemert, M.J. Characterization of synthetic Chikungunya viruses based on the consensus sequence of recent E1-226V isolates. PLoS ONE 2013, 8, e71047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coffey, L.L.; Beeharry, Y.; Borderia, A.V.; Blanc, H.; Vignuzzi, M. Arbovirus high fidelity variant loses fitness in mosquitoes and mice. Proc. Natl. Acad. Sci. USA 2011, 108, 16038–16043. [Google Scholar] [CrossRef] [PubMed]
- Rozen-Gagnon, K.; Stapleford, K.A.; Mongelli, V.; Blanc, H.; Failloux, A.-B.; Saleh, M.-C.; Vignuzzi, M. Alphavirus Mutator Variants Present Host-Specific Defects and Attenuation in Mammalian and Insect Models. PLoS Pathog. 2014, 10, e1003877. [Google Scholar] [CrossRef] [PubMed]
- Urakova, N.; Kuznetsova, V.; Crossman, D.K.; Sokratian, A.; Guthrie, D.B.; Kolykhalov, A.A.; Lockwood, M.A.; Natchus, M.G.; Crowley, M.R.; Painter, G.R.; et al. β-d-N(4)-hydroxycytidine is a potent anti-alphavirus compound that induces high level of mutations in viral genome. J. Virol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Delang, L.; Guerrero, N.S.; Tas, A.; Quérat, G.; Pastorino, B.; Froeyen, M.; Dallmeier, K.; Jochmans, D.; Herdewijn, P.; Bello, F.; et al. Mutations in the chikungunya virus non-structural proteins cause resistance to favipiravir (T-705), a broad-spectrum antiviral. J. Antimicrob. Chemother. 2014, 69, 2770–2784. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y.; Orba, Y.; Sasaki, M.; Kobayashi, S.; Carr, M.J.; Nobori, H.; Sato, A.; Hall, W.W.; Sawa, H. Discovery of a novel antiviral agent targeting the nonstructural protein 4 (nsP4) of chikungunya virus. Virology 2017, 505, 102–112. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(A) Schematic representation of the alphavirus genome showing the RNA sequence open reading frames (ORFs). The (*) indicates the position of opal termination codon; (B) schematic representation of non-structural polyprotein (nsP2) processing by nsP2 protease. Early processing of P1234 produces P123 and nsP4 which associate to form the early replication complex (RC), which performs negative-sense RNA synthesis. P123 is further processed to produce the individual nsPs, which associate to form mature RC that regulates positive-sense RNA synthesis and transcription of subgenomic 26S RNA.
Figure 1.
(A) Schematic representation of the alphavirus genome showing the RNA sequence open reading frames (ORFs). The (*) indicates the position of opal termination codon; (B) schematic representation of non-structural polyprotein (nsP2) processing by nsP2 protease. Early processing of P1234 produces P123 and nsP4 which associate to form the early replication complex (RC), which performs negative-sense RNA synthesis. P123 is further processed to produce the individual nsPs, which associate to form mature RC that regulates positive-sense RNA synthesis and transcription of subgenomic 26S RNA.

Figure 2.
(A) Illustration of superposed structures of papain and chikungunya virus (CHIKV) nsP2 proteases (Protein Data Bank (PDB) 9PAD and 3TRK respectively). The structure of papain protease is presented as a solid, blue ribbon. The structure of CHIKV nsP2 protease is presented as solid, green ribbon. The domain common for both proteases is highlighted in dark colors and is enlarged subsequently to show the conserved catalytic dyad; (B) alignment of nsP2 cleavage sites. The nomenclature of Berger and Schechter is used to identify residues on the amino (P1, P2, etc.) or carboxy (P1′, P2′, etc.) termini of the scissile bond. The arrow indicates the location of the cleavage site. Cleavage sites between the non-structural proteins contain a common motif, AG(A/C)↓(G/Y/A) [55]. Abbreviations: Chikungunya virus (CHIKV), o’nyong’nyong virus (ONNV), semliki forest virus (SFV), ross river virus (RRV), sindbis virus (SINV) and Venezuelan equine encephalitis virus (VEEV).
Figure 2.
(A) Illustration of superposed structures of papain and chikungunya virus (CHIKV) nsP2 proteases (Protein Data Bank (PDB) 9PAD and 3TRK respectively). The structure of papain protease is presented as a solid, blue ribbon. The structure of CHIKV nsP2 protease is presented as solid, green ribbon. The domain common for both proteases is highlighted in dark colors and is enlarged subsequently to show the conserved catalytic dyad; (B) alignment of nsP2 cleavage sites. The nomenclature of Berger and Schechter is used to identify residues on the amino (P1, P2, etc.) or carboxy (P1′, P2′, etc.) termini of the scissile bond. The arrow indicates the location of the cleavage site. Cleavage sites between the non-structural proteins contain a common motif, AG(A/C)↓(G/Y/A) [55]. Abbreviations: Chikungunya virus (CHIKV), o’nyong’nyong virus (ONNV), semliki forest virus (SFV), ross river virus (RRV), sindbis virus (SINV) and Venezuelan equine encephalitis virus (VEEV).


Table 1.
Non-structural protein (nsP) domain functions and available crystal structures.
| Non-Structural Protein | Domain Function | Virus | PDB ID | Reference |
|---|---|---|---|---|
| nsP1 | mRNA capping | SFV | 1FW5 | [45] |
| nsP2 | NTPase/HelicaseProtease | - VEEV CHIKV SINV | - 2HWK, 5EZQ 3TRK 4GUA | - [29,30] [31] [32] |
| nsP3 | Macrodomain | VEEV CHIKV SINV | 3GQE 3GPG 4GUA | [60] [60] [32] |
| nsP4 | RNA-dependent RNA Polymerase | - | - | [32] |
PDB: Protein data bank; SFV: Semliki Forest virus; VEEV: Venezuelan equine encephalitis virus; CHIKV: chikungunya virus ; SINV: sindbis virus.




{kind=link}
{kind=link}
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Abu Bakar, F.; Ng, L.F.P. Nonstructural Proteins of Alphavirus—Potential Targets for Drug Development. Viruses 2018, 10, 71. https://0-doi-org.brum.beds.ac.uk/10.3390/v10020071
AMA Style
Abu Bakar F, Ng LFP. Nonstructural Proteins of Alphavirus—Potential Targets for Drug Development. Viruses. 2018; 10(2):71. https://0-doi-org.brum.beds.ac.uk/10.3390/v10020071
Chicago/Turabian StyleAbu Bakar, Farhana, and Lisa F. P. Ng. 2018. "Nonstructural Proteins of Alphavirus—Potential Targets for Drug Development" Viruses 10, no. 2: 71. https://0-doi-org.brum.beds.ac.uk/10.3390/v10020071
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.