The Emergence of Avian Orthoavulavirus 13 in Wild Migratory Waterfowl in China Revealed the Existence of Diversified Trailer Region Sequences and HN Gene Lengths within this Serotype

 ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical States
2.2. Sample Collection and Virus Isolation
2.3. RNA Extraction, cNDA Synthesis, PCR and Whole Genome Sequencing
2.4. Pathogenicity Test and Virus Infection of Cells
2.5. Virus Infection of Chickens
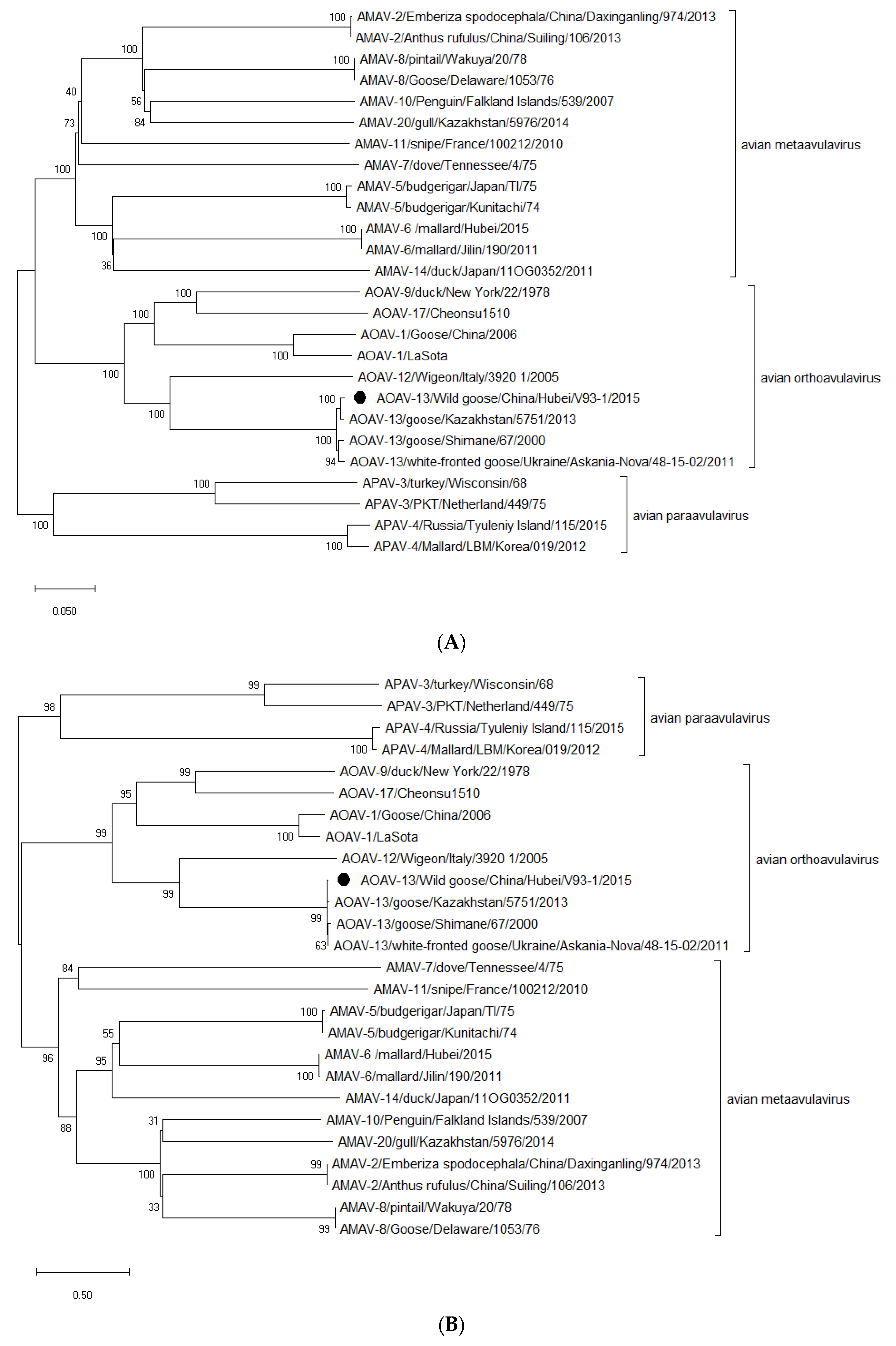
2.6. Phylogenetic Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Samal, S.K. The Biology of Paramyxoviruses; Caister Academic Press: Norfolk, UK, 2011; 469p. [Google Scholar]
- Kolakofsky, D.; Roux, L. The molecular biology of paramyxoviruses. In The Molecular Basis of Viral Replication; Bercoff, R.P., Ed.; Springer: Boston, MA, USA, 1987; pp. 277–297. [Google Scholar]
- Saif, Y.M.; Barnes, H.J. Diseases of Poultry, 12th ed.; Saif, Y.M., Fadly, A.M., Eds.; Blackwell: Oxford, UK, 2008; pp. 75–116. [Google Scholar]
- Amarasinghe, G.K.; Bao, Y.; Basler, C.F.; Bavari, S.; Beer, M.; Bejerman, N.; Blasdell, K.R.; Bochnowski, A.; Briese, T.; Bukreyev, A.; et al. Taxonomy of the order mononegavirales: Update 2017. Arch. Virol. 2017, 162, 2493–2504. [Google Scholar] [CrossRef] [PubMed]
- Thomazelli, L.M.; de Araujo, J.; Fabrizio, T.; Walker, D.; Reischak, D.; Ometto, T.; Barbosa, C.M.; Petry, M.V.; Webby, R.J.; Durigon, E.L. Novel avian paramyxovirus (apmv-15) isolated from a migratory bird in south america. PLoS ONE 2017, 12, e0177214. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, E.; Ito, T.; Ito, H. Completion of full length genome sequence of novel avian paramyxovirus strain apmv/shimane67 isolated from migratory wild geese in japan. J. Vet. Med. Sci. 2016, 78, 1583–1594. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Kim, J.Y.; Lee, Y.J.; Lee, E.K.; Song, B.M.; Lee, H.S.; Choi, K.S. A novel avian paramyxovirus (putative serotype 15) isolated from wild birds. Front. Microbiol. 2017, 8, 786. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Kim, Y.; An, I.; Wang, S.J.; Kim, Y.; Lee, H.J.; Choi, K.S.; Im, S.P.; Min, W.; Oem, J.K.; et al. Complete genome sequence of a novel avian paramyxovirus isolated from wild birds in south Korea. Arch. Virol. 2018, 163, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Thampaisarn, R.; Bui, V.N.; Trinh, D.Q.; Nagai, M.; Mizutani, T.; Omatsu, T.; Katayama, Y.; Gronsang, D.; Le, D.H.T.; Ogawa, H.; et al. Characterization of avian paramyxovirus serotype 14, a novel serotype, isolated from a duck fecal sample in Japan. Virus Res. 2017, 228, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Wille, M.; Aban, M.; Wang, J.; Moore, N.; Shan, S.; Marshall, J.; Acuna, D.G.; Vijaykrishna, D.; Butler, J.; Wang, J.; et al. Antarctic penguins as reservoirs of diversity for avian avulaviruses. J. Virol. 2019, 93, e00271-19. [Google Scholar] [CrossRef] [PubMed]
- Amarasinghe, G.K.; Ayllon, M.A.; Bao, Y.; Basler, C.F.; Bavari, S.; Blasdell, K.R.; Briese, T.; Brown, P.A.; Bukreyev, A.; Balkema-Buschmann, A.; et al. Taxonomy of the order mononegavirales: Update 2019. Arch. Virol. 2019, 164, 1967–1980. [Google Scholar] [CrossRef] [PubMed]
- Umali, D.V.; Ito, H.; Katoh, H.; Ito, T. Surveillance of avian paramyxovirus in migratory waterfowls in the san-in region of western Japan from 2006 to 2012. J. Vet. Med. Sci. 2014, 76, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Cardenas-Garcia, S.; Diel, D.G.; Susta, L.; Lucio-Decanini, E.; Yu, Q.; Brown, C.C.; Miller, P.J.; Afonso, C.L. Development of an improved vaccine evaluation protocol to compare the efficacy of newcastle disease vaccines. Biol. J. Int. Assoc. Biol. Stand. 2015, 43, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Yamamo, E.; Ito, H.; Tomioka, Y.; Ito, T. Characterization of novel avian paramyxovirus strain apmv/shimane67 isolated from migratory wild geese in japan. J. Vet. Med. Sci. 2015, 77, 1079–1085. [Google Scholar] [CrossRef] [PubMed]
- Goraichuk, I.; Poonam, S.; Dimitrov, K.; Stegniy, B.; Muzyka, D.; Pantin-Jackwood, M.; Gerilovych, A.; Solodiankin, O.; Bolotin, V.; Rula, O.; et al. Phylogenetic analysis of the complete genome of the apmv-13 isolate from Ukraine. Int. J. Infect. Dis. 2016, 45, 459. [Google Scholar] [CrossRef]
- Karamendin, K.; Kydyrmanov, A.; Seidalina, A.; Asanova, S.; Sayatov, M.; Kasymbekov, E.; Khan, E.; Daulbayeva, K.; Harrison, S.M.; Carr, I.M.; et al. Complete genome sequence of a novel avian paramyxovirus (apmv-13) isolated from a wild bird in Kazakhstan. Genome Announc. 2016, 4, e00167-16. [Google Scholar] [CrossRef] [PubMed]
- Karamendin, K.; Kydyrmanov, A.; Seidalina, A.; Asanova, S.; Daulbayeva, K.; Kasymbekov, Y.; Khan, E.; Fereidouni, S.; Starick, E.; Zhumatov, K.; et al. Circulation of avian paramyxoviruses in wild birds of Kazakhstan in 2002–2013. Virol. J. 2016, 13, 23. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Chen, Q.; Wang, Q.; Chen, J.; Jin, T.; Wong, G.; Quan, C.; Liu, J.; Wu, J.; Yin, R.; et al. Genesis, evolution and prevalence of h5n6 avian influenza viruses in China. Cell Host Microbe 2016, 20, 810–821. [Google Scholar] [CrossRef]
- Yin, R.; Zhang, P.; Liu, X.; Chen, Y.; Tao, Z.; Ai, L.; Li, J.; Yang, Y.; Li, M.; Xue, C.; et al. Dispersal and transmission of avian paramyxovirus serotype 4 among wild birds and domestic poultry. Front. Cell Infect. Microbiol. 2017, 7, 212. [Google Scholar] [CrossRef]
- Chen, Y.; Ding, Z.; Liu, X.; Chen, J.; Li, J.; Fei, Y.; Liu, Z.; Stoeger, T.; Bi, Y.; Yin, R. Biological and phylogenetic characterization of a novel hemagglutination-negative avian avulavirus 6 isolated from wild waterfowl in China. Transbound Emerg. Dis. 2018, 65, 1421–1428. [Google Scholar] [CrossRef]
- Zhang, P.; Xie, G.; Liu, X.; Ai, L.; Chen, Y.; Meng, X.; Bi, Y.; Chen, J.; Sun, Y.; Stoeger, T.; et al. High genetic diversity of newcastle disease virus in wild and domestic birds in northeastern China from 2013 to 2015 reveals potential epidemic trends. Appl. Environ. Microbiol. 2015, 82, 1530–1536. [Google Scholar] [CrossRef]
- Newcastle Disease. Chapter 2.3.14. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; The World Organisation for Animal Health: Paris, France, 2012; pp. 555–574. Available online: http://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.03.14_NEWCASTLE_DIS.pdf (accessed on 12 July 2019).
- Tong, S.; Chern, S.W.; Li, Y.; Pallansch, M.A.; Anderson, L.J. Sensitive and broadly reactive reverse transcription-pcr assays to detect novel paramyxoviruses. J. Clin. Microbiol. 2008, 46, 2652–2658. [Google Scholar] [CrossRef]
- Dimitrov, K.M.; Sharma, P.; Volkening, J.D.; Goraichuk, I.V.; Wajid, A.; Rehmani, S.F.; Basharat, A.; Shittu, I.; Joannis, T.M.; Miller, P.J.; et al. A robust and cost-effective approach to sequence and analyze complete genomes of small RNA viruses. Virol. J. 2017, 14, 72. [Google Scholar] [CrossRef]
- Afgan, E.; Baker, D.; van den Beek, M.; Blankenberg, D.; Bouvier, D.; Cech, M.; Chilton, J.; Clements, D.; Coraor, N.; Eberhard, C.; et al. The galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2016 update. Nucleic Acids Res. 2016, 44, W3–W10. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega x: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, H.L.; Taylor, T.L.; Dimitrov, K.M.; Sabra, M.; Afonso, C.L.; Suarez, D.L. Virulent newcastle disease viruses from chicken origin are more pathogenic and transmissible to chickens than viruses normally maintained in wild birds. Vet. Microbiol. 2019, 235, 25–34. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fei, Y.; Liu, X.; Mu, J.; Li, J.; Yu, X.; Chang, J.; Bi, Y.; Stoeger, T.; Wajid, A.; Muzyka, D.; et al. The Emergence of Avian Orthoavulavirus 13 in Wild Migratory Waterfowl in China Revealed the Existence of Diversified Trailer Region Sequences and HN Gene Lengths within this Serotype. Viruses 2019, 11, 646. https://0-doi-org.brum.beds.ac.uk/10.3390/v11070646
Fei Y, Liu X, Mu J, Li J, Yu X, Chang J, Bi Y, Stoeger T, Wajid A, Muzyka D, et al. The Emergence of Avian Orthoavulavirus 13 in Wild Migratory Waterfowl in China Revealed the Existence of Diversified Trailer Region Sequences and HN Gene Lengths within this Serotype. Viruses. 2019; 11(7):646. https://0-doi-org.brum.beds.ac.uk/10.3390/v11070646
Chicago/Turabian StyleFei, Yidong, Xinxin Liu, Jiaqi Mu, Junjiao Li, Xibing Yu, Jin Chang, Yuhai Bi, Tobias Stoeger, Abdul Wajid, Denys Muzyka, and et al. 2019. "The Emergence of Avian Orthoavulavirus 13 in Wild Migratory Waterfowl in China Revealed the Existence of Diversified Trailer Region Sequences and HN Gene Lengths within this Serotype" Viruses 11, no. 7: 646. https://0-doi-org.brum.beds.ac.uk/10.3390/v11070646