Identification of Immunogenic Epitopes That Permit the Detection of Antigen-Specific T Cell Responses in Multiple Serotypes of Group B Coxsackievirus Infections

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Peptide Synthesis and Immunization Procedures
2.3. Virus Propagation and Infection
2.4. Proliferative Response
2.5. MHC Class II Binding
2.6. Derivation of MHC Class II Dextramers and Tetramers
2.7. Staining with Dextramers and Tetramers
2.8. Cytokine Analysis
2.9. Vβ Usage
2.10. Histology
2.11 Statistics
3. Results
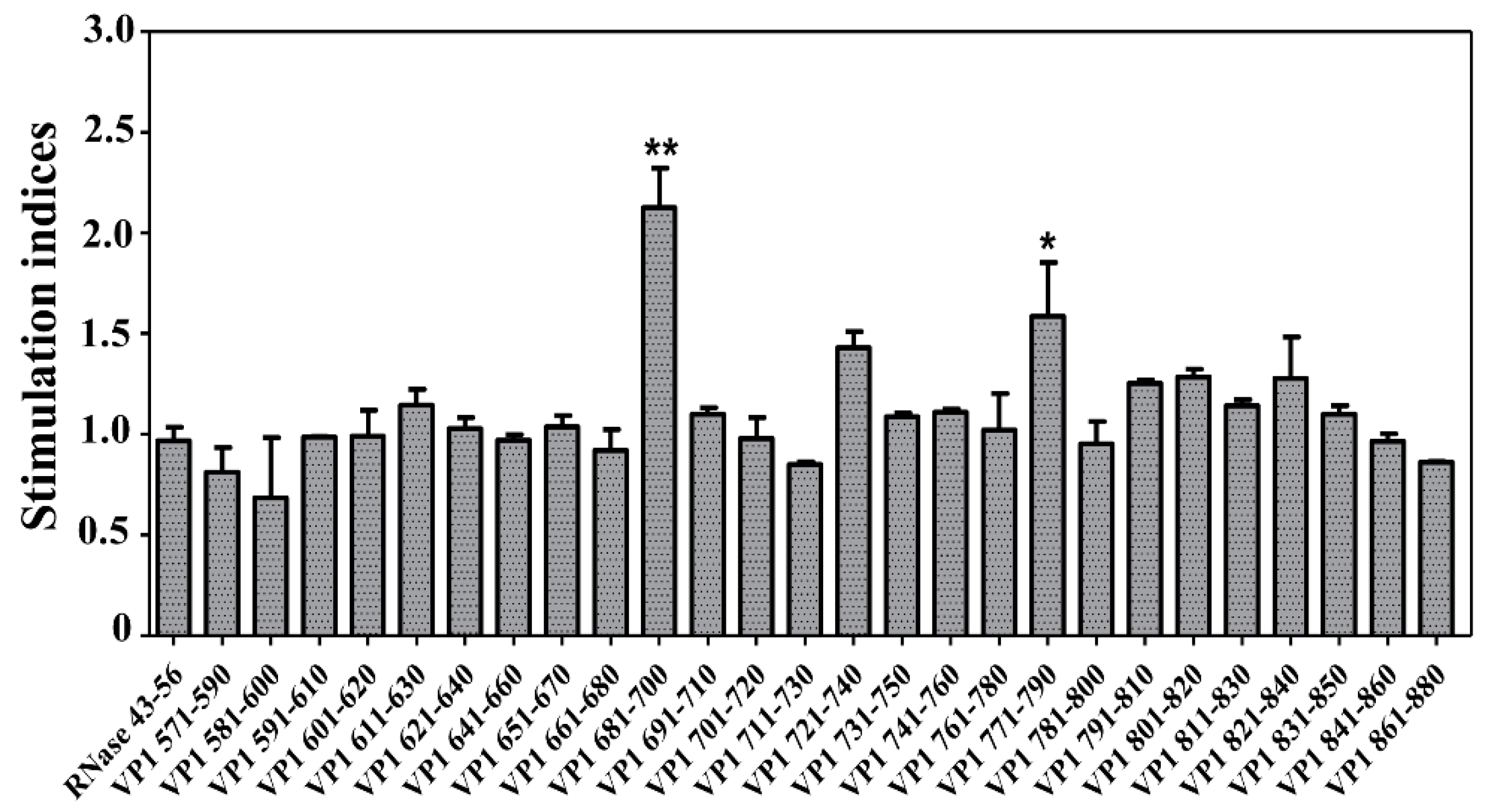
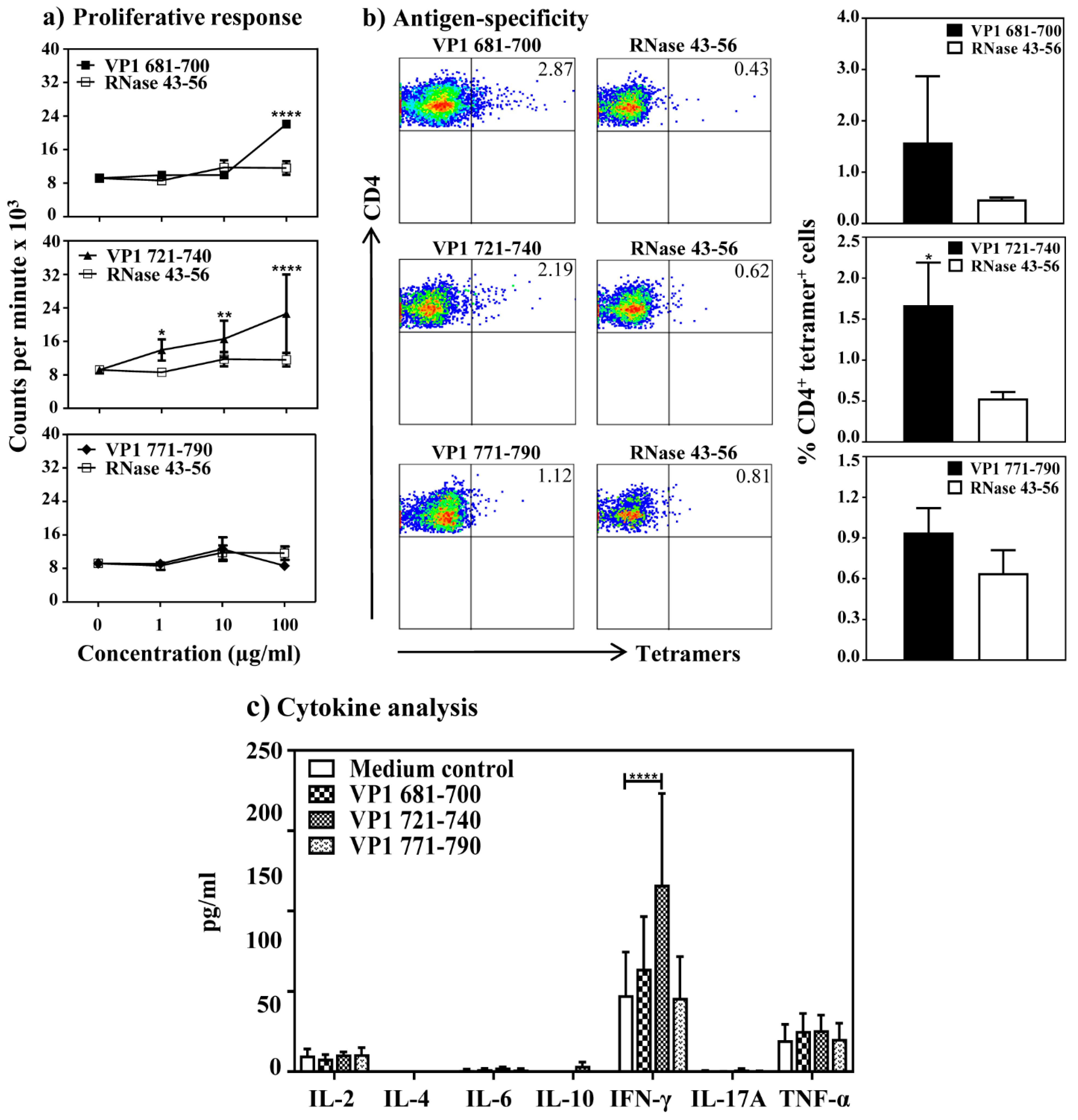
3.1. CVB3 VP1 Contains Multiple Immunogenic Epitopes
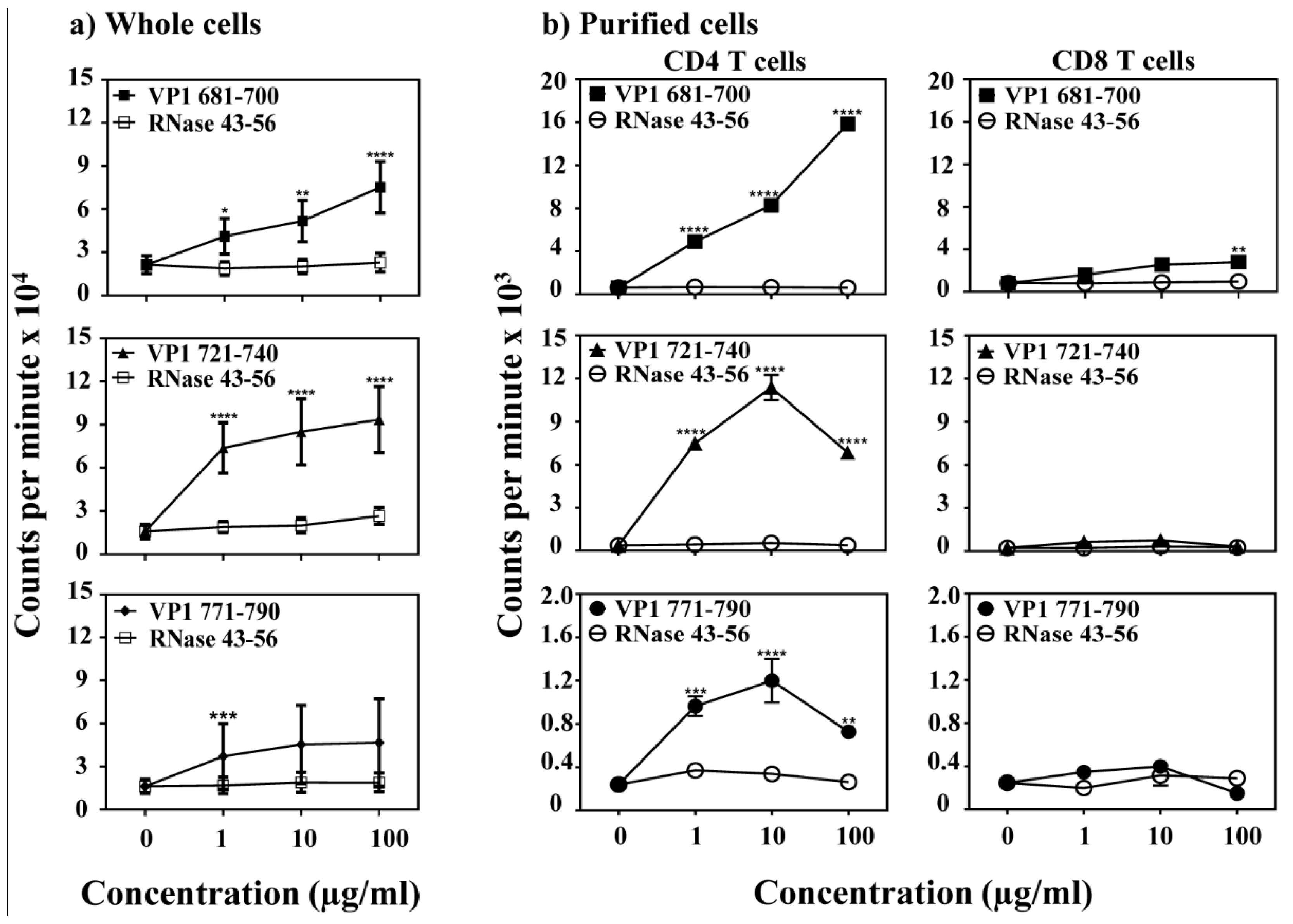
3.2. VP1 Epitopes Induce Mainly CD4 T Cell Responses
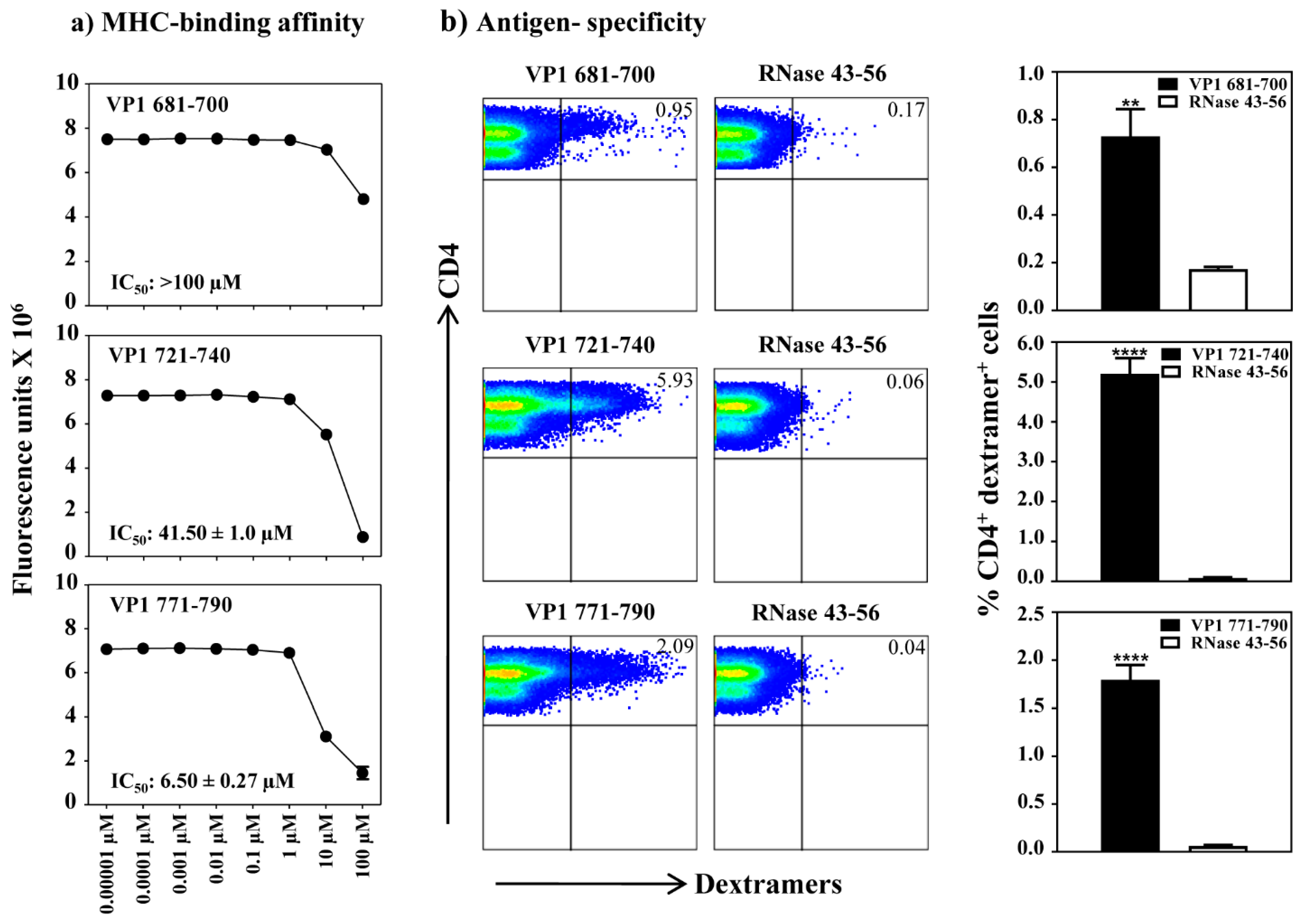
3.3. The VP1 Epitopes Bind IAk Molecules Differentially and the T Cell Responses Are Antigen-Specific
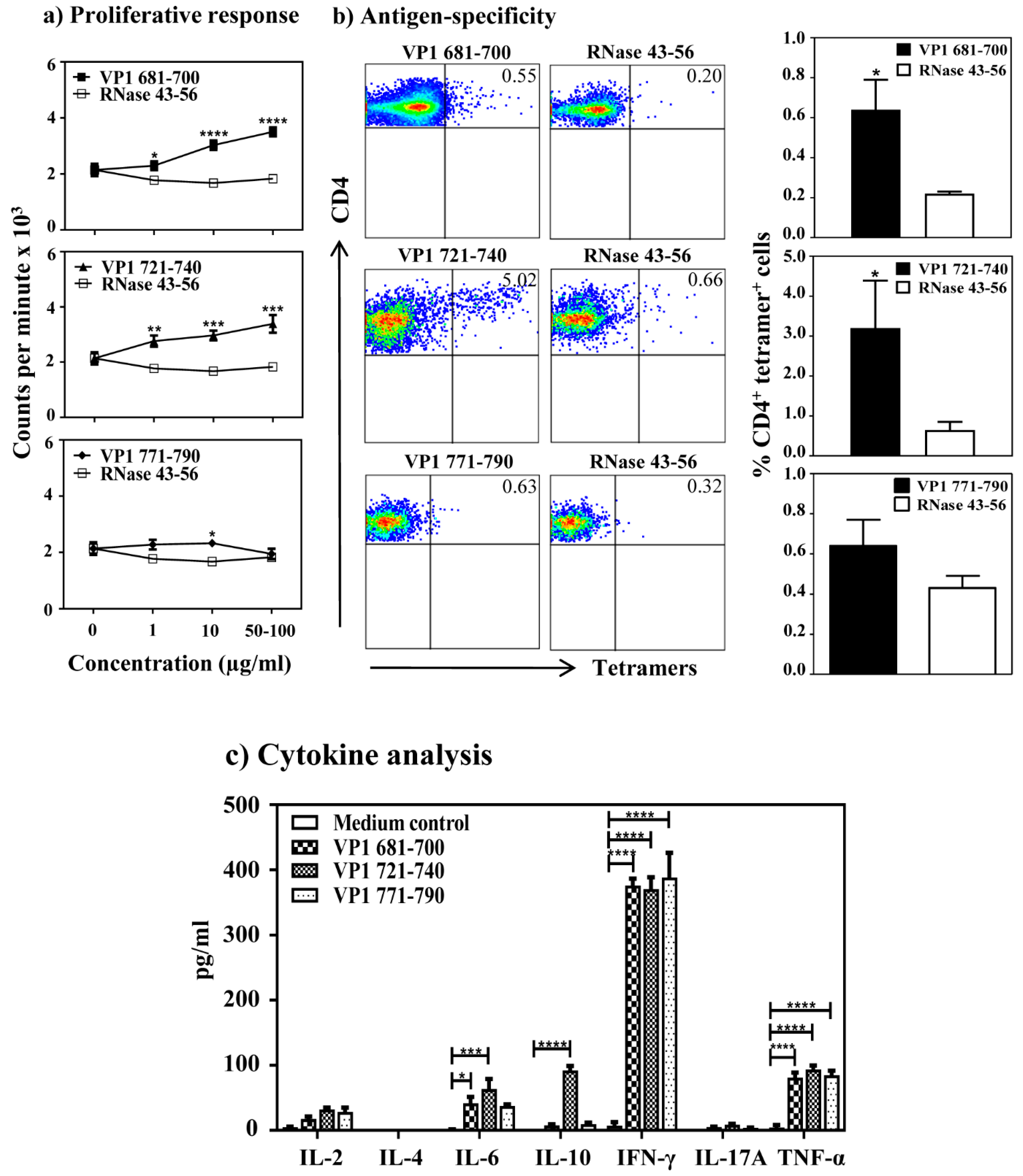
3.4. CVB3 Infection Leads to the Induction of VP1 Virus-Specific, CD4 T Cell Responses
3.5. Multiple Serotypes of CVB Can Potentially Induce Similar T Cell Responses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DCM | Dilated cardiomyopathy |
| CVB | Coxsackievirus B |
| T1D | Type 1 diabetes |
| MHC | major histocompatibility complex |
| CPE | cytopathic effect |
| RNase | bovine ribonuclease |
| HEL | hen egg lysozyme |
| APCs | antigen presenting cells |
| PBS | phosphate-buffered saline |
| LNCs | lymph node cells |
| EMEM | Eagle’s minimum essential medium |
| FBS | fetal bovine serum |
| 3[H] | tritiated-thymidine |
| Cpm | counts per minute |
| HLA | human leukocyte antigen |
| IEDB | immune epitope database |
| IL | Interleukin |
| 7-AAD | 7-aminoactinomycin-D |
| IFN | interferon |
| VP | viral protein |
| TMEV | Theiler’s murine encephalomyelitis virus |
| DELFIA | dissociation-enhanced lanthanide fluoro immuno assay |
| Th | T helper |
| CLIP | class II invariant chain-associated peptide |
| TNF | tumor necrosis factor |
| MBP | myelin basic protein |
| SA | streptavidin |
| CFA | Complete Freund’s adjuvant |
References
- Zaoutis, T.; Klein, J.D. Enterovirus infections. Pediatr. Rev. Am. Acad. Pediatr. 1998, 19, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Rhoades, R.E.; Tabor-Godwin, J.M.; Tsueng, G.; Feuer, R. Enterovirus infections of the central nervous system. Virology 2011, 411, 288–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tracy, S.; Hofling, K.; Pirruccello, S.; Lane, P.H.; Reyna, S.M.; Gauntt, C.J. Group B coxsackievirus myocarditis and pancreatitis: Connection between viral virulence phenotypes in mice. J. Med. Virol. 2000, 62, 70–81. [Google Scholar] [CrossRef]
- Archard, L.C.; Bowles, N.E.; Cunningham, L.; Freeke, C.A.; Olsen, E.G.; Rose, M.L.; Meany, B.; Why, H.J.; Richardson, P.J. Molecular probes for detection of persisting enterovirus infection of human heart and their prognostic value. Eur. Heart J. 1991, 12 (Suppl. D), 56–59. [Google Scholar] [CrossRef]
- Cihakova, D.; Rose, N.R. Pathogenesis of myocarditis and dilated cardiomyopathy. Adv. Immunol. 2008, 99, 95–114. [Google Scholar]
- Kuhl, U.; Pauschinger, M.; Noutsias, M.; Seeberg, B.; Bock, T.; Lassner, D.; Poller, W.; Kandolf, R.; Schultheiss, H.P. High prevalence of viral genomes and multiple viral infections in the myocardium of adults with “idiopathic” left ventricular dysfunction. Circulation 2005, 111, 887–893. [Google Scholar] [CrossRef] [Green Version]
- Martino, T.; Liu, P.; Sole, M.J. Enteroviral myocarditis and dialted cardiomyopathy: A review of clinical and experimental studies. In Human Enterovirus Infections; ASM: Washington, DC, USA, 1995. [Google Scholar]
- Chapman, N.M.; Kim, K.S. Persistent coxsackievirus infection: Enterovirus persistence in chronic myocarditis and dilated cardiomyopathy. Curr. Top. Microbiol. Immunol. 2008, 323, 275–292. [Google Scholar]
- Rose, N.R.; Wolfgram, L.J.; Herskowitz, A.; Beisel, K.W. Postinfectious autoimmunity: Two distinct phases of coxsackievirus B3-induced myocarditis. Ann. N. Y. Acad. Sci. 1986, 475, 146–156. [Google Scholar] [CrossRef]
- Yoon, J.W.; Austin, M.; Onodera, T.; Notkins, A.L. Isolation of a virus from the pancreas of a child with diabetic ketoacidosis. N. Engl. J. Med. 1979, 300, 1173–1179. [Google Scholar] [CrossRef]
- Dotta, F.; Censini, S.; van Halteren, A.G.; Marselli, L.; Masini, M.; Dionisi, S.; Mosca, F.; Boggi, U.; Muda, A.O.; Del Prato, S.; et al. Coxsackie B4 virus infection of beta cells and natural killer cell insulitis in recent-onset type 1 diabetic patients. Proc. Natl. Acad. Sci. USA 2007, 104, 5115–5120. [Google Scholar] [CrossRef] [Green Version]
- Clements, G.B.; Galbraith, D.N.; Taylor, K.W. Coxsackie B virus infection and onset of childhood diabetes. Lancet 1995, 346, 221–223. [Google Scholar] [CrossRef]
- Gamble, D.R.; Kinsley, M.L.; FitzGerald, M.G.; Bolton, R.; Taylor, K.W. Viral antibodies in diabetes mellitus. Br. Med. J. 1969, 3, 627–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drescher, K.M.; Tracy, S.M. The CVB and etiology of type 1 diabetes. Curr. Top. Microbiol. Immunol. 2008, 323, 259–274. [Google Scholar] [PubMed]
- Tracy, S.; Drescher, K.M. Coxsackievirus infections and NOD mice: Relevant models of protection from, and induction of, type 1 diabetes. Ann. N. Y. Acad. Sci. 2007, 1103, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Tracy, S.; Drescher, K.M.; Chapman, N.M.; Kim, K.S.; Carson, S.D.; Pirruccello, S.; Lane, P.H.; Romero, J.R.; Leser, J.S. Toward testing the hypothesis that group B coxsackieviruses (CVB) trigger insulin-dependent diabetes: Inoculating nonobese diabetic mice with CVB markedly lowers diabetes incidence. J. Virol. 2002, 76, 12097–12111. [Google Scholar] [CrossRef] [Green Version]
- Filippi, C.M.; Estes, E.A.; Oldham, J.E.; von Herrath, M.G. Immunoregulatory mechanisms triggered by viral infections protect from type 1 diabetes in mice. J. Clin. Investig. 2009, 119, 1515–1523. [Google Scholar] [CrossRef] [Green Version]
- Cihakova, D.; Barin, J.G.; Afanasyeva, M.; Kimura, M.; Fairweather, D.; Berg, M.; Talor, M.V.; Baldeviano, G.C.; Frisancho, S.; Gabrielson, K. Interleukin-13 protects against experimental autoimmune myocarditis by regulating macrophage differentiation. Am. J. Pathol. 2008, 172, 1195–1208. [Google Scholar] [CrossRef] [Green Version]
- Massilamany, C.; Gangaplara, A.; Basavalingappa, R.H.; Rajasekaran, R.A.; Khalilzad-Sharghi, V.; Han, Z.; Othman, S.; Steffen, D.; Reddy, J. Localization of CD8 T cell epitope within cardiac myosin heavy chain-α 334–352 that induces autoimmune myocarditis in A/J mice. Int. J. Cardiol. 2016, 202, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Basavalingappa, R.H.; Massilamany, C.; Krishnan, B.; Gangaplara, A.; Kang, G.; Khalilzad-Sharghi, V.; Han, Z.; Othman, S.; Li, Q.; Riethoven, J.-J. Identification of an Epitope from Adenine Nucleotide Translocator 1 That Induces Inflammation in Heart in A/J Mice. Am. J. Pathol. 2016, 186, 3160–3175. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, B.; Massilamany, C.; Basavalingappa, R.H.; Gangaplara, A.; Kang, G.; Li, Q.; Uzal, F.A.; Strande, J.L.; Delhon, G.A.; Riethoven, J.J. Branched chain α-ketoacid dehydrogenase kinase 111–130, a T cell epitope that induces both autoimmune myocarditis and hepatitis in A/J mice. Immun. Inflamm. Dis. 2017, 5, 421–434. [Google Scholar] [CrossRef] [Green Version]
- Massilamany, C.; Gangaplara, A.; Steffen, D.; Reddy, J. Identification of novel mimicry epitopes for cardiac myosin heavy chain-α that induce autoimmune myocarditis in A/J mice. Cell. Immunol. 2011, 271, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Massilamany, C.; Gangaplara, A.; Chapman, N.; Rose, N.; Reddy, J. Detection of cardiac myosin heavy chain-alpha-specific CD4 cells by using MHC class II/IA(k) tetramers in A/J mice. J. Immunol. Methods 2011, 372, 107–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, J.; Bettelli, E.; Nicholson, L.; Waldner, H.; Jang, M.H.; Wucherpfennig, K.W.; Kuchroo, V.K. Detection of autoreactive myelin proteolipid protein 139-151-specific T cells by using MHC II (IAs) tetramers. J. Immunol. 2003, 170, 870–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, C.L.; Seth, N.P.; Lucas, M.; Appel, H.; Gauthier, L.; Lauer, G.M.; Robbins, G.K.; Szczepiorkowski, Z.M.; Casson, D.R.; Chung, R.T.; et al. Ex vivo analysis of human memory CD4 T cells specific for hepatitis C virus using MHC class II tetramers. J. Clin. Investig. 2003, 112, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Hausmann, D.H.; Yu, B.; Hausmann, S.; Wucherpfennig, K.W. pH-dependent peptide binding properties of the type I diabetes-associated I-Ag7 molecule: Rapid release of CLIP at an endosomal pH. J. Exp. Med. 1999, 189, 1723–1734. [Google Scholar] [CrossRef] [Green Version]
- Fugger, L.; Liang, J.; Gautam, A.; Rothbard, J.B.; McDevitt, H.O. Quantitative analysis of peptides from myelin basic protein binding to the MHC class II protein, I-Au, which confers susceptibility to experimental allergic encephalomyelitis. Mol. Med. 1996, 2, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Jang, M.H.; Seth, N.P.; Wucherpfennig, K.W. Ex vivo analysis of thymic CD4 T cells in nonobese diabetic mice with tetramers generated from I-A(g7)/class II-associated invariant chain peptide precursors. J. Immunol. 2003, 171, 4175–4186. [Google Scholar] [CrossRef] [Green Version]
- Massilamany, C.; Upadhyaya, B.; Gangaplara, A.; Kuszynski, C.; Reddy, J. Detection of autoreactive CD4 T cells using major histocompatibility complex class II dextramers. BMC Immunol. 2011, 12, 40. [Google Scholar] [CrossRef] [Green Version]
- Massilamany, C.; Steffen, D.; Reddy, J. An epitope from Acanthamoeba castellanii that cross-react with proteolipid protein 139-151-reactive T cells induces autoimmune encephalomyelitis in SJL mice. J. Neuroimmunol. 2010, 219, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Massilamany, C.; Thulasingam, S.; Steffen, D.; Reddy, J. Gender differences in CNS autoimmunity induced by mimicry epitope for PLP 139-151 in SJL mice. J. Neuroimmunol. 2011, 230, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Daniel, W.W. Applied Nonparametric Statistics; Houghton Mifflin: Boston, MA, USA, 1978; pp. xiii, 503. [Google Scholar]
- Barnard, G.A. A New Test for 2 × 2 Tables. Nature 1945, 156, 177. [Google Scholar] [CrossRef]
- Yauch, R.L.; Kim, B.S. A predominant viral epitope recognized by T cells from the periphery and demyelinating lesions of SJL/J mice infected with Theiler’s virus is located within VP1(233-244). J. Immunol. 1994, 153, 4508–4519. [Google Scholar] [PubMed]
- Zurbriggen, A.; Hogle, J.M.; Fujinami, R.S. Alteration of amino acid 101 within capsid protein VP-1 changes the pathogenicity of Theiler’s murine encephalomyelitis virus. J. Exp. Med. 1989, 170, 2037–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, E.J.; Jeffrey, P.D. Sequence homology between encephalomyocarditis virus protein VPI and histidyl-tRNA synthetase supports a hypothesis of molecular mimicry in polymyositis. Med. Hypotheses 1988, 25, 21–25. [Google Scholar] [CrossRef]
- Babbitt, B.P.; Allen, P.M.; Matsueda, G.; Haber, E.; Unanue, E.R. Binding of immunogenic peptides to Ia histocompatibility molecules. Nature 1985, 317, 359–361. [Google Scholar] [CrossRef]
- Basavalingappa, R.H.; Massilamany, C.; Krishnan, B.; Gangaplara, A.; Rajasekaran, R.A.; Afzal, M.Z.; Riethoven, J.J.; Strande, J.L.; Steffen, D.; Reddy, J. beta1-Adrenergic Receptor Contains Multiple IA(k) and IE(k) Binding Epitopes That Induce T Cell Responses with Varying Degrees of Autoimmune Myocarditis in A/J Mice. Front. Immunol. 2017, 8, 1567. [Google Scholar] [CrossRef] [Green Version]
- Massilamany, C.; Gangaplara, A.; Jia, T.; Elowsky, C.; Kang, G.; Riethoven, J.-J.; Li, Q.; Zhou, Y.; Reddy, J. Direct staining with major histocompatibility complex class II dextramers permits detection of antigen-specific, autoreactive CD4 T cells in situ. PLoS ONE 2014, 9, e87519. [Google Scholar] [CrossRef]
- Muckelbauer, J.K.; Kremer, M.; Minor, I.; Tong, L.; Zlotnick, A.; Johnson, J.E.; Rossmann, M.G. Structure determination of coxsackievirus B3 to 3.5 A resolution. Acta Cryst. D Biol. Cryst. 1995, 51, 871–887. [Google Scholar] [CrossRef]
- Jaidane, H.; Hober, D. Role of coxsackievirus B4 in the pathogenesis of type 1 diabetes. Diabetes Metab. 2008, 34, 537–548. [Google Scholar] [CrossRef]
- Huber, S.; Polgar, J.; Moraska, A.; Cunningham, M.; Schwimmbeck, P.; Schultheiss, P. T lymphocyte responses in CVB3-induced murine myocarditis. Scand. J. Infect. Dis. Suppl. 1993, 88, 67–78. [Google Scholar]
- Blay, R.; Simpson, K.; Leslie, K.; Huber, S. Coxsackievirus-induced disease. CD4+ cells initiate both myocarditis and pancreatitis in DBA/2 mice. Am. J. Pathol. 1989, 135, 899–907. [Google Scholar] [PubMed]
- Vella, C.; Brown, C.L.; McCarthy, D.A. Coxsackievirus B4 infection of the mouse pancreas: Acute and persistent infection. J. Gen. Virol. 1992, 73, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Baxter, N.J.; Roetzer, A.; Liebig, H.D.; Sedelnikova, S.E.; Hounslow, A.M.; Skern, T.; Waltho, J.P. Structure and dynamics of coxsackievirus B4 2A proteinase, an enyzme involved in the etiology of heart disease. J. Virol. 2006, 80, 1451–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairweather, D.; Stafford, K.A.; Sung, Y.K. Update on coxsackievirus B3 myocarditis. Curr. Opin. Rheumatol. 2012, 24, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, M.; Azuma, E.; Jiang, Q.; Kobayashi, M.; Iwamoto, S.; Kumamoto, T.; Kisenge, R.; Yamamoto, H.; Komada, Y. The reconstitution of CD45RBhiCD4+ naive T cells is inversely correlated with donor age in murine allogeneic haematopoietic stem cell transplantation. Br. J. Haematol. 2000, 111, 700–707. [Google Scholar] [CrossRef]
- Lee, C.; Liang, M.N.; Tate, K.M.; Rabinowitz, J.D.; Beeson, C.; Jones, P.P.; McConnell, H.M. Evidence that the autoimmune antigen myelin basic protein (MBP) Ac1-9 binds towards one end of the major histocompatibility complex (MHC) cleft. J. Exp. Med. 1998, 187, 1505–1516. [Google Scholar] [CrossRef] [Green Version]
- Boyman, O. Bystander activation of CD4+ T cells. Eur. J. Immunol. 2010, 40, 936–939. [Google Scholar] [CrossRef]
- Varela-Calvino, R.; Ellis, R.; Sgarbi, G.; Dayan, C.M.; Peakman, M. Characterization of the T-cell response to coxsackievirus B4: Evidence that effector memory cells predominate in patients with type 1 diabetes. Diabetes 2002, 51, 1745–1753. [Google Scholar] [CrossRef] [Green Version]
- Vita, R.; Mahajan, S.; Overton, J.A.; Dhanda, S.K.; Martini, S.; Cantrell, J.R.; Wheeler, D.K.; Sette, A.; Peters, B. The Immune Epitope Database (IEDB): 2018 update. Nucleic Acids Res. 2019, 47, D339–D343. [Google Scholar] [CrossRef] [Green Version]
- Sin, J.; Mangale, V.; Thienphrapa, W.; Gottlieb, R.A.; Feuer, R. Recent progress in understanding coxsackievirus replication, dissemination, and pathogenesis. Virology 2015, 484, 288–304. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | Peptide | Sequence |
|---|---|---|---|
| VP1 571–590 | GPVEDAITAAIGRVADTVGT | VP1 721–740 | PDKVDSYVWQTSTNPSVFWT |
| VP1 581–600 | IGRVADTVGTGPNNSEAIPA | VP1 731–750 | TSTNPSVFWTEGNAPPRMSI |
| VP1 591–610 | GPNNSEAIPALTAAETGHTS | VP1 741–760 | EGNAPPRMSIPFLSIGNAYS |
| VP1 601–620 | LTAAETGHTSQVVPGDTMQT | VP1 751–770 | PFLSIGNAYSNFYDGWSEFS |
| VP1 611–630 | QVVPGDTMQTRHVKNYHSRS | VP1 761–780 | NFYDGWSEFSRNGVYGINTL |
| VP1 621–640 | RHVKNYHSRSESTIENFLCR | VP1 771–790 | RNGVYGINTLNNMGTLYARH |
| VP1 631–650 | ESTIENFLCRSACVYFTEYE | VP1 781–800 | NNMGTLYARHVNAGSTGPIK |
| VP1 641–660 | SACVYFTEYENSGAKRYAEW | VP1 791–810 | VNAGSTGPIKSTIRIYFKPK |
| VP1 651–670 | NSGAKRYAEWVLTPRQAAQL | VP1 801–820 | STIRIYFKPKHVKAWIPRPP |
| VP1 661–680 | VLTPRQAAQLRRKLEFFTYV | VP1 811–830 | HVKAWIPRPPRLCQYEKAKN |
| VP1 671–690 | RRKLEFFTYVRFDLELTFVI | VP1 821–840 | RLCQYEKAKNVNFQPSGVTT |
| VP1 681–700 | RFDLELTFVITSTQQPSTTQ | VP1 831–850 | VNFQPSGVTTTRQSITTMTN |
| VP1 691–710 | TSTQQPSTTQNQDAQILTHQ | VP1 841–860 | TRQSITTMTNTGAIWTTIRG |
| VP1 701–720 | NQDAQILTHQIMYVPPGGPV | VP1 851–870 | TGAIWTTIRGSVCGDYRVVN |
| VP1 711–730 | IMYVPPGGPVPDKVDSYVWQ | VP1 861–880 | SVCGDYRVVNRHSATSADWQ |
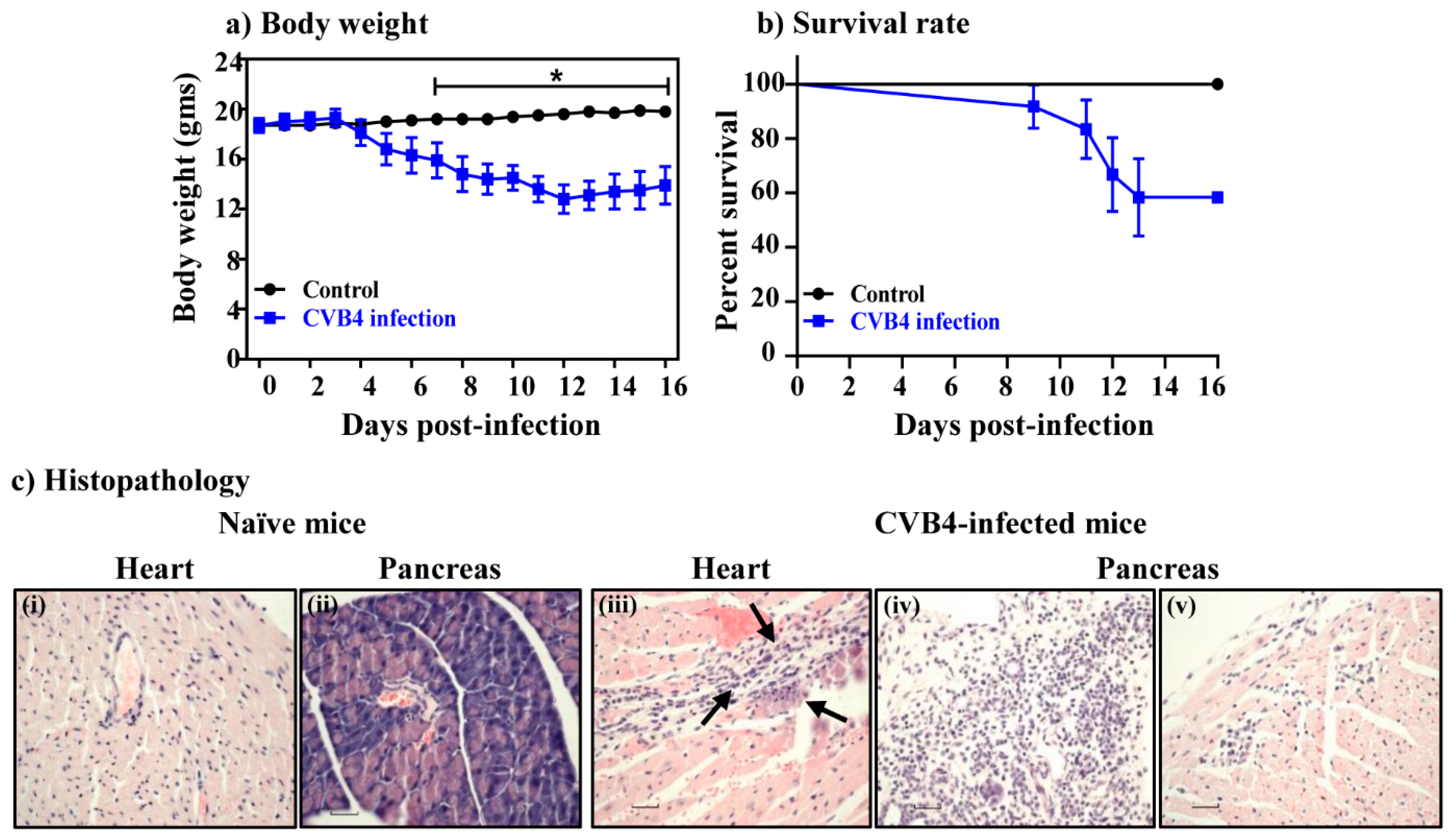
| Parameters | Naïve | CVB4 |
|---|---|---|
| Heart | ||
| Incidence | 0/4 (0.0) | 4/12 (33.3) |
| Mortality | 0/4 (0.0) | 5/12 (41.7) |
| Myocardial lesions | 0.0 (0.0) | 10.0 ± 1.2 |
| Pancreas | ||
| Incidence | 0 (0.0) | 11/12 (91.7) |
| Atrophy | 0 (0.0) | 10/12 (83.3) |
| Inflammation | 0 (0.0) | 6/12 (50.0) |
| Necrosis | 0 (0.0) | 2/12 (16.7) |
| Mineralization | 0 (0.0) | 0/12 (0.0) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lasrado, N.; Gangaplara, A.; Arumugam, R.; Massilamany, C.; Pokal, S.; Zhou, Y.; Xiang, S.-H.; Steffen, D.; Reddy, J. Identification of Immunogenic Epitopes That Permit the Detection of Antigen-Specific T Cell Responses in Multiple Serotypes of Group B Coxsackievirus Infections. Viruses 2020, 12, 347. https://0-doi-org.brum.beds.ac.uk/10.3390/v12030347
Lasrado N, Gangaplara A, Arumugam R, Massilamany C, Pokal S, Zhou Y, Xiang S-H, Steffen D, Reddy J. Identification of Immunogenic Epitopes That Permit the Detection of Antigen-Specific T Cell Responses in Multiple Serotypes of Group B Coxsackievirus Infections. Viruses. 2020; 12(3):347. https://0-doi-org.brum.beds.ac.uk/10.3390/v12030347
Chicago/Turabian StyleLasrado, Ninaad, Arunakumar Gangaplara, Rajkumar Arumugam, Chandirasegaran Massilamany, Sayli Pokal, Yuzhen Zhou, Shi-Hua Xiang, David Steffen, and Jay Reddy. 2020. "Identification of Immunogenic Epitopes That Permit the Detection of Antigen-Specific T Cell Responses in Multiple Serotypes of Group B Coxsackievirus Infections" Viruses 12, no. 3: 347. https://0-doi-org.brum.beds.ac.uk/10.3390/v12030347