1. Introduction
Hepatitis C virus (HCV) is a member of the
Flaviviridae family and was identified in 1989 as the infectious agent of non-A, non-B hepatitis. Currently, HCV is the leading cause of end-stage liver disease as a result of cirrhosis and/or hepatocellular carcinoma (HCC). An estimated 71 million people are chronically infected and approximately 400,000 associated deaths occur each year worldwide [
1,
2]. Although safe, tolerable and curative therapies for HCV infection have emerged in recent years, the prevention, clinical management and access to treatment remain important determinants in the control of HCV infection. Despite the recent therapeutic advances, HCV pathophysiology is still not entirely elucidated justifying continued research in this field [
3].
HCV-infected hepatocytes are exposed to several stressors that may affect their function and viability. These stressors include viral replication and viral protein production within hepatocytes, as well as the inflammatory response of the host. It is known that HCV infection leads to increased oxidative stress in the liver and in particular in the hepatocytes [
4]. Since HCV replication and viral protein production are closely linked to the endoplasmic reticulum (ER), both ER stress and oxidative stress occur and contribute to the progression of chronic HCV-related liver disease; therefore, hepatocytes should adapt to injury insult [
5,
6,
7,
8,
9]. However, little is known about the consequences of this adaptation upon HCV infection.
HCV contains a positive sense single-stranded RNA (ssRNA+) genome that encodes for a polyprotein of approximately 3100 amino acids, depending on the genotype, that is cleaved co- and post-translationally by cellular and viral proteases to produce 10 viral proteins with various structural and biochemical functions (Core, E1, E2, p7, NS2, NS3, NS4A, NS4B, NS5A and NS5B) [
10]. The role of HCV proteins in the generation of oxidative stress and ER stress has been demonstrated and Core and the non-structural proteins NS3/4A and NS5A are the most potent inducers [
11,
12,
13].
In mammalian cells, different signaling pathways have evolved to mediate the cellular stress response. One of the most conserved regulatory events activated in response to stress is the phosphorylation of the α subunit of eukaryotic translation Initiation Factor 2 (eIF2α) at serine 51 and subsequent ATF4 (Activation Transcription Factor 4) activation [
14]. It has been demonstrated that activation of the eIF2α/ATF4 pathway directs an autophagy gene transcriptional program to overcome cellular stress. Furthermore, the transcription factors ATF4 and CCAAT/Enhancer-Binding Protein Homologous Protein (CHOP) are involved in the transcriptional activation of other autophagy genes, including p62/SQSTM1 (Sequestosome 1) [hereafter referred to as p62] [
15].
Autophagy can also be induced via activation of the Unfolded Protein Response (UPR) through phosphorylation of the ER stress sensors: Protein Kinase R (PKR)-like endoplasmic reticulum kinase (PERK) and Inositol-Requiring protein 1 (IRE1, or Endoplasmic reticulum to nucleus signaling 1, human homologue). Both sensors can induce and activate Beclin-1, as well as the expression of autophagy-related genes (ATGs),
ATG5 and
ATG12 [
16,
17]. Additionally, PERK phosphorylation is known to couple distinct upstream stress signals to eIF2α/ATF4 pathway activation and further autophagy gene expression [
15]. In Huh7 hepatoma cells, infection with HCV leads to induction of the UPR and subsequently to an inhibition of the phosphatidylinositol 3-kinase (PI3K)/AKT/mammalian Target of the Rapamycin (mTOR) signaling pathway, resulting in induction of autophagy [
16]. HCV-induced autophagy can also be mediated via an UPR-independent mechanism, since silencing of the UPR-sensor IRE1 in HCV-infected Huh7.5 cells did not affect HCV replication or the induction of autophagy [
18].
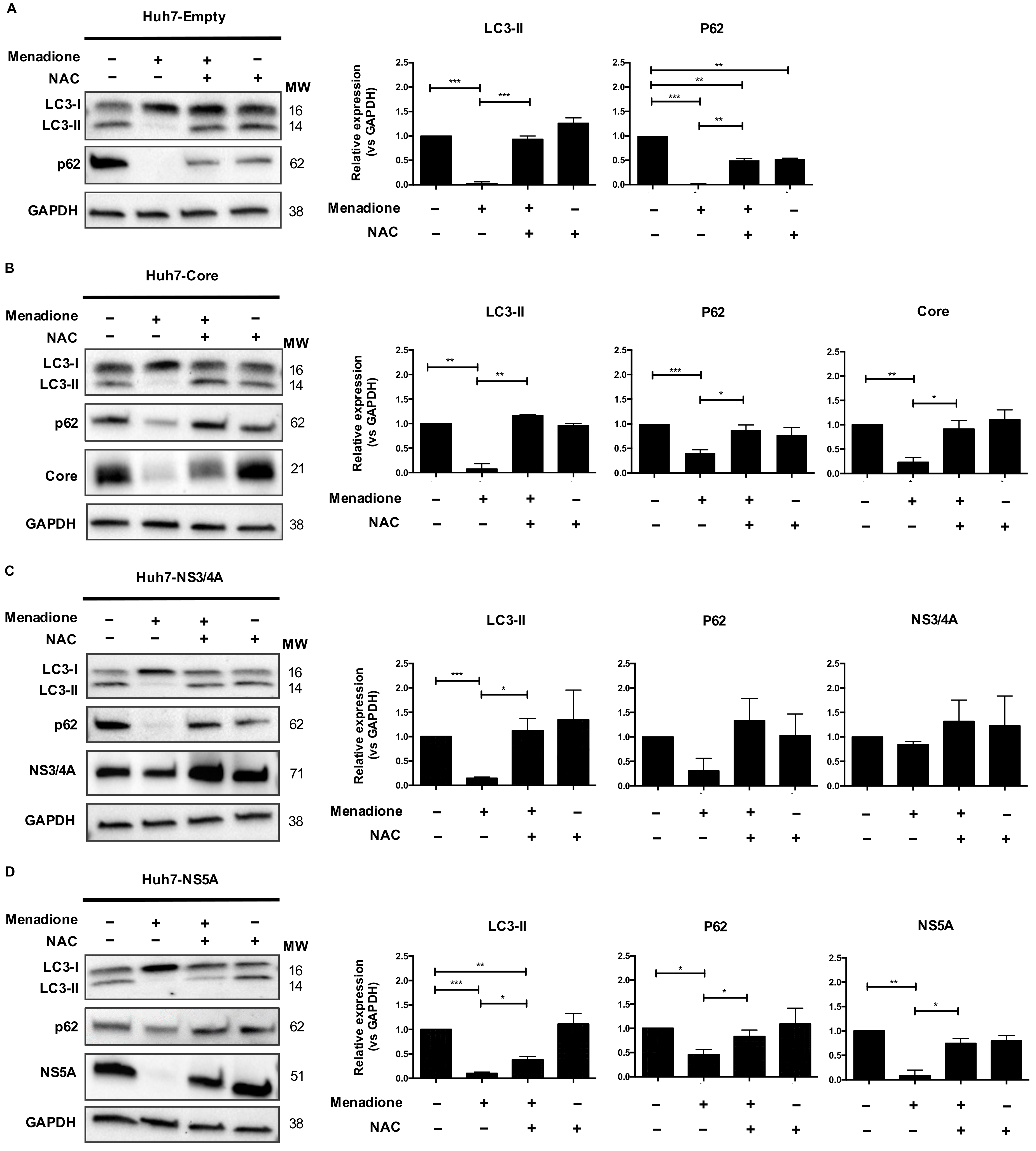
In a previous study, we established a model in Huh7 cells, as well as in primary rat hepatocytes, to investigate the adaptive responses activated under HCV protein expression and external cellular oxidative stress induction. HCV Core or NS3/4A were transiently expressed in hepatocytes and subsequently treated with menadione, a superoxide anion donor. We observed that under induction of external oxidative stress and HCV protein expression, both pro-oxidants, hepatocytes adapt to oxidative stress via a reduction in reactive oxygen species (ROS) production as well as reduction of oxidative stress-induced apoptosis [
19]. In addition, we observed an increased degradation of the HCV Core protein together with the autophagy adaptor protein p62 in hepatocytes resistant to oxidative stress [
19].
In the present study, we investigated the adaptive response to stress in cells overexpressing HCV proteins. Autophagy-related proteins and changes in their steady state protein levels were investigated and related to protection against cell death in the context of HCV protein overexpression, oxidative stress and ER stress. We observed that selective degradation of HCV Core and NS5A proteins via activation of the eIF2α/ATF4 pathway plays an important role in the adaptive response of hepatocytes to stress via suppression of pro-oxidant agents.
2. Material and Methods
2.1. Cell Lines and Culture
Huh7 cells were maintained in Dulbecco’s Modified Eagle Medium (1X) + GlutaMAX
TM- I (DMEM; Gibco, Landsmeer, The Netherlands) supplemented with 10% fetal bovine serum (FBS, Gibco, Landsmeer, The Netherlands) and 1% penicillin-streptomycin (Gibco, Landsmeer, The Netherlands) at 37 °C and 5% CO
2. The stable cell lines Huh7_puro (3 µg/mL puromycin) (containing the empty vector), Huh7_Core_Jc1_bla (10 µg/mL blasticidin), Huh7_NS3/4A_Con1 (0.75 µg/mL G418) and Huh7_NS5A_JFH1_puro (3 µg/mL puromycin) were generated and kindly provided by Prof. Dr. Ralf Bartenschlager from the University of Heidelberg, Germany [
20].
2.2. Reagents and Treatments
Cells (3.0 × 105) were grown until 80% confluence in 6-well plates 24 h (h) prior to treatment. Cells were treated for 6 h with 50 µmol/L menadione (Sigma, Zwijndrecht, The Netherlands) or 5 mmol/L hydrogen peroxide (H2O2) (Sigma, Zwijndrecht, The Netherlands) to induce oxidative stress. Subsequently, cells were harvested and viability was determined by trypan blue exclusion staining. For the kinetic assays cells were collected after menadione treatment every hour for 6 h. In control experiments, 5 mmol/L N-acetyl-l-cysteine (NAC, Sigma, Zwijndrecht, The Netherlands) was added 30 min prior to menadione treatment. In some experiments 50 µmol/L chloroquine (CQ) diphosphate salt (Sigma, Zwijndrecht, The Netherlands) was used to inhibit autophagic flux and 100 nmol/L bafilomycin A1 (Sigma, Zwijndrecht, The Netherlands) or a mixture of 20 mmol/L ammoniumchloride (NH4Cl)/100 µmol/L leupeptin/100 µmol/L pepstatin was used to inhibit the lysosomal degradation pathway. For proteasome inhibition 10 µmol/L MG132 (Sigma, Zwijndrecht, The Netherlands) was added 3 h prior to menadione treatment. Experiments were conducted in duplicate and the results are expressed as the average of three independent experiments.
2.3. Tissue Samples
Liver tissue samples from patients submitted for liver transplantation were obtained from the tissue bank of the Gastrohepatology Group, University of Antioquia, Colombia. The samples were selected according to etiology and used for RNA isolation and quantitative polymerase chain reaction (qPCR) analysis. The liver tissues were from patients with cirrhosis associated to HCV infection (6 patient samples), HCV-associated hepatocellular carcinoma (HCC) (7 patient samples), hepatitis B virus (HBV)-associated cirrhosis (4 patient samples) and non-viral liver disease (4 patient samples). The demographic and clinical characteristics of the samples are presented in
Table 1. For total RNA isolation 100 mg of tissue was processed using TriZOL reagent (Invitrogen, Landsmeer, The Netherlands) following the manufacturer’s instructions. Total RNA (2.5 µg) was used for reverse transcription (RT). Complementary DNA (cDNA) was diluted 20 X in nuclease free water and stored at −20 °C until use.
2.4. Cell Culture, RNA Isolation and Quantitative Polymerase Chain Reaction (qPCR)
After treatment, Huh7 cells stably expressing the empty vector, Core, NS3/4A and NS5A were harvested on ice and washed three times with ice-cold 1X Hank’s Balanced Salt Solution (HBSS) with Ca
2+ and Mg
2+ (Gibco, Landsmeer, The Netherlands). TRI reagent (Sigma, Zwijndrecht, The Netherlands) was added to the cells for total RNA isolation according to the manufacturer’s instructions. Total RNA (2.5 µg) was used for RT in 1X RT buffer (500 mmol/L Tris-HCl -pH 8.3-; 500 mmol/L KCl; 30 mmol/L MgCl
2; and 50 mmol/L DTT), 1 mmol/L deoxynucleotides triphosphate (dNTPs, Sigma, Zwijndrecht, The Netherlands), 10 ng/µL random nanomers (Sigma, Zwijndrecht, The Netherlands), 0.6 U/µL RNaseOUT
TM (Invitrogen, Landsmeer, The Netherlands) and 4 U/µL Moloney-Murine Leukemia Virus (M-MLV) reverse transcriptase (Invitrogen, Landsmeer, The Netherlands) in a final volume of 50 µL. cDNA was diluted 20 X in nuclease free water and qPCR was carried out in a StepOnePlus™ (96-well) PCR System (Applied Biosystems, Landsmeer, The Netherlands) using TaqMan probes; the sequences of the probes and primers are described in
Supplementary Table S1. For qPCR, 2X reaction buffer (dNTPs, HotGoldStar DNA polymerase, 5 mmol/L MgCl
2) (Eurogentech, Maastricht, The Netherlands), 5 µmol/L fluorogenic probe and 50 µmol/L of primers sense and antisense (Invitrogen, Landsmeer, The Netherlands) were used. mRNA levels were normalized to 18S gene expression and compared between groups [
21]. The experiments were performed in duplicate and presented as the average of three independent experiments.
2.5. Determination of Cellular Oxidative Stress
Total cytoplasmic ROS was quantified using the fluorogenic probe CellROX® Deep Red (Invitrogen, Landsmeer, The Netherlands) following the manufacturer‘s instructions. After induction of oxidative stress, 5 µmol/L of CellROX reagent was added to the cells and cells were subsequently incubated at 37 °C and 5% CO2 for 30 min. Media was removed and cells were washed three times with 1X HBSS with Ca2+, Mg2+ (Gibco, Landsmeer, The Netherlands) and subsequently harvested using 1X trypsin (Gibco, Landsmeer, The Netherlands) and analyzed by flow cytometry using a BD FACSVerse system and a 635nm laser. Three independent experiments were carried out and the results are expressed as an average.
2.6. Caspase 3 Activity Determination and Flow Cytometry
After treatment, cells were scraped on ice and lysed by three cycles of freezing (liquid nitrogen) and thawing (37 °C) in lysis buffer (25 mmol/L HEPES, 150 mmol/L KAc, 2 mmol/L EDTA, 0.1% NP-40) supplemented with protease and phosphatase inhibitors (10 mmol/L NaF, 50 mmol/L PMSF, 1 µg/µL of α-protenin/pepstatin/leupeptin and 1 mmol/L DTT) followed by centrifugation for 10 min at 12,000 rpm. For the caspase 3 activity assay [
22], 30 µg of protein was mixed with the synthetic fluorogenic caspase 3 substrate, Ac-DEVD-AMC and the release of fluorogenic AMC was quantified in a spectrofluorometer at an excitation wavelength of 380 nm and emission wavelength of 430 nm. The arbitrary units of fluorescence (AUF) from three independent experiments were used to depict the results. Multiparametric apoptosis assay by flow cytometry was performed using MitoProbe
TM DilC(5) assay kit in combination with propidium iodide (PI) staining following the manufacturer’s protocol (ThermoFisher Scientific, Landsmeer, The Netherlands). Cells were harvested after menadione treatment (50 µmol/L) using trypsin. Then, 10 µM DilC(5) and 100 µg/mL PI were added to the cells.
2.7. Transfection of siRNA
For silencing of p62, 4 × 104 cells were seeded in 12-well plates pre-treated with 1.5 µL Lipofectamine 3000 (Invitrogen, Landsmeer, The Netherlands) and 50 μmol/L esiRNA human p62/SQSTM 1 (esiRNA1) (Sigma, Zwijndrecht, The Netherlands, Cat. #EHU027651) or scrambled siRNA (esiRNA Egfp Cat. #EHUEGFP-20UG, Sigma, Zwijndrecht, The Netherlands) as a control. The Lipofectamine 3000 and the esiRNAs were prepared in 75 μL OPTI-MEMTM I (1X) reduced serum medium (Gibco, Landsmeer, The Netherlands) following the manufacturer’s instructions then wells were cover with the mixture (reverse transfection). After cells were added, media was completed to 750 μL final volume per well. 12 h post-transfection, media were replaced and a second transfection round was performed using the same amounts of Lipofectamine 3000 and esiRNA for 12 h. Subsequently, cells were treated with 50 μmol/L menadione for 6 h. After treatment cells were scraped and lysed by freezing and thawing cycles in lysis buffer containing protease and phosphatase inhibitors as described above, followed by centrifugation for 10 min at 12,000 rpm. Supernatant was collected and stored at −20 °C until use. Three independent experiments were performed and the results are expressed as means ±S.D.
2.8. Western Blotting
Cell lysates (20 μg) were resolved on Mini-PROTEAN
® TGX Stain-Free
TM Precast Gels (BioRad, Veenendaal, The Netherlands) and semi-dry blotting transfer was performed using Trans-Blot Turbo Midi Nitrocellulose Membrane with Trans-Blot Turbo System (BioRad, Veenendaal, The Netherlands). To confirm the electrophoretic transfer, Ponceau S 0.1%
w/
v (Sigma, Zwijndrecht, The Netherlands) staining was used. The monoclonal antibodies human anti-HCV Core B12-F8 (kindly provided by prof. Dr. Mondelli, University of Pavia, Italy) [
23], mouse anti-HCV NS3/4A (8 G-2) (Abcam, Cambridge, UK) and mouse anti-HCV NS5A (9E10) (kindly provided by prof. Dr. Charles M. Rice), were used at a dilution of 1:1000 and mouse anti-glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (Calbiochem, Amsterdam, The Netherlands) at a dilution of 1:10,000. ER stress markers were also determined using the polyclonal rabbit antibodies anti-peIF2α (Cell Signaling, Leiden, The Netherlands), anti-eIF2α (total) (Cell Signaling, Leiden, The Netherlands), anti-ATF4 (Cell Signaling, Leiden, The Netherlands) and anti-glucose-regulated orotein of 78kDa (GRP78) (Cell Signaling, Leiden, The Netherlands) at 1:1000 dilution. Polyclonal rabbit anti-Microtubule Associated Protein 1 Light Chain 3 Beta (LC3B) (Cell Signaling, Leiden, The Netherlands) and anti-p62 (Cell Signaling, Leiden, The Netherlands) were also used at 1:1000 dilution. For detection of ubiquitinated proteins mouse anti-α-Ubiquitin (1/1000) (Hycult, Biotech, Uden, The Netherlands) was used. Secondary horseradish peroxidase (HRP)-bound antibodies were used. The blots were analyzed by chemiluminescence in a ChemiDoc XRS system (Bio-Rad, Veenendaal, The Netherlands). Protein band intensities were quantified by ImageLab software (BioRad, Veenendaal, The Netherlands).
2.9. Statistical Analysis
All experiments were performed at least three times and the mean ± standard deviation (s.d.) is depicted. The Graphpad Prism 5 software (GraphPad Software) was used and comparisons were evaluated by unpaired, two-tailed t-test. For the group analysis two tails Anova and Bonferroni post-test were performed. A p value of <0.05 was considered statistically significant.
4. Discussion
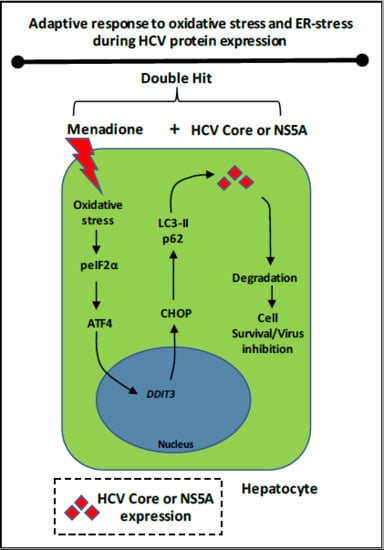
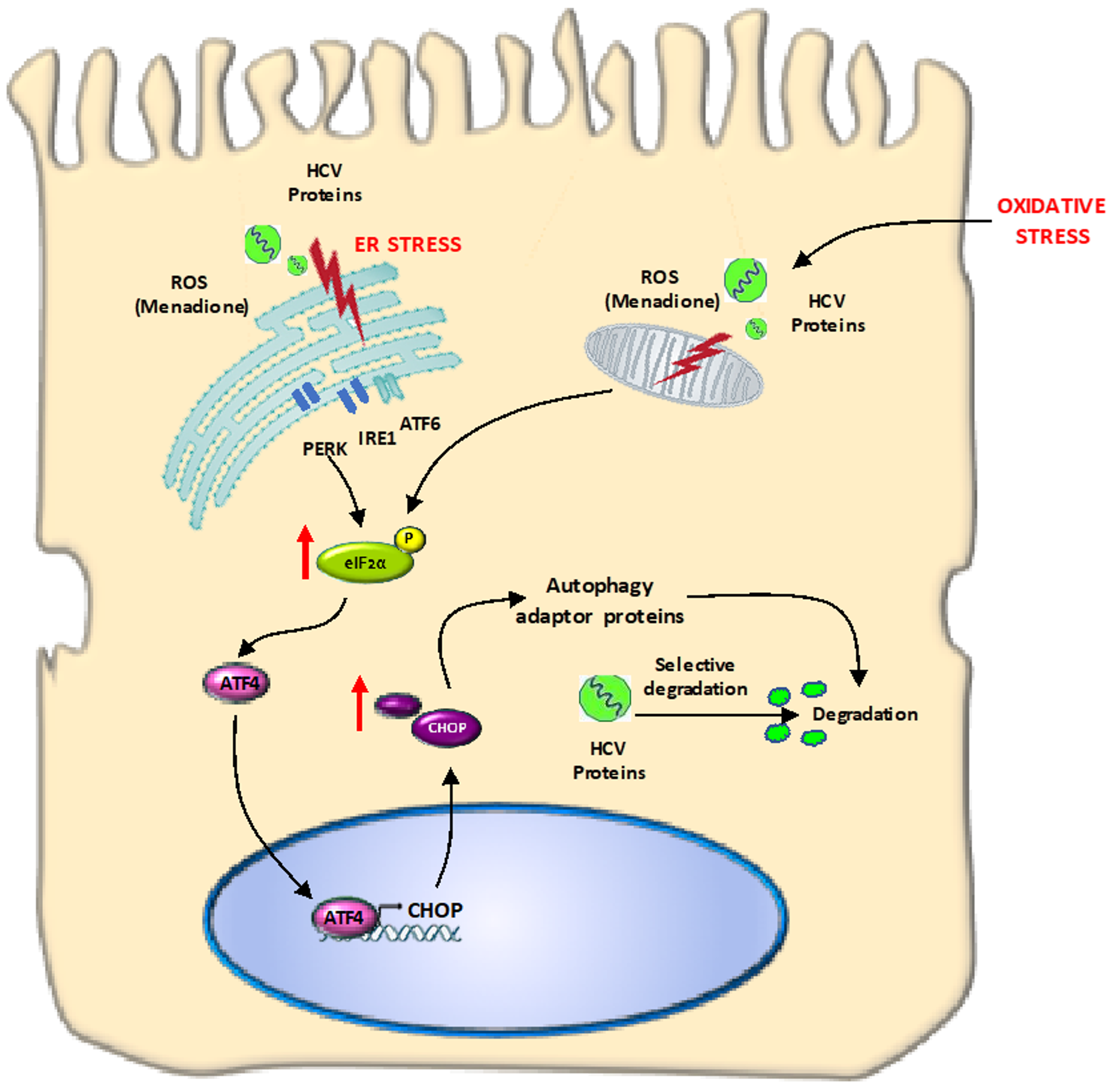
The goal of the present study was to investigate the adaptive response of hepatocyte-like cells to multiple stressors including oxidative stress and overexpression of pro-oxidant HCV proteins. Our findings can have relevance for other liver diseases accompanied by increased oxidative stress, e.g., non-alcoholic steatohepatitis (NASH) and alcoholic steatohepatitis (ASH). We found that Huh7 cells expressing HCV Core and NS5A resist the deleterious effects from additional oxidative stress via selective degradation of these viral proteins involving autophagy adaptor proteins such as p62 and LC3. The degradation occurs in response to activation of the eIF2α/ATF4 pathway and suggest elimination of harmful viral proteins that could increase the cytotoxic effects from oxidative stress (
Figure 9).
Infection of cells by viruses constitutes an important stress to the host cells. Cells must support viral replication and synthesis and shedding of newly synthesized virions. In addition, in most viral infections, cells are also exposed to immune and/or inflammatory response to the viral infection. Therefore, cells have to adapt to multiple stressors in order to survive and thus also sustain the viral replication cycle. HCV infection of hepatocytes is an example in which cells are subjected to multiple stressors: viral protein synthesis may lead to ER stress and HCV infection is also accompanied by oxidative stress [
4,
25]. In a previous study, we observed increased resistance to menadione-induced oxidative stress using Huh7 cells and primary rat hepatocytes transiently expressing HCV Core or NS3/4A proteins. Both mitochondrial ROS production and menadione-induced apoptosis were significantly decreased together with reduced levels of the ER stress markers GRP78 and sXBP1 in Huh7 cells expressing HCV Core and NS3/4A proteins. This increased resistance was accompanied by increased degradation of the HCV Core protein in these cells, suggesting that selective degradation of one stressor may be in part responsible for this increased resistance [
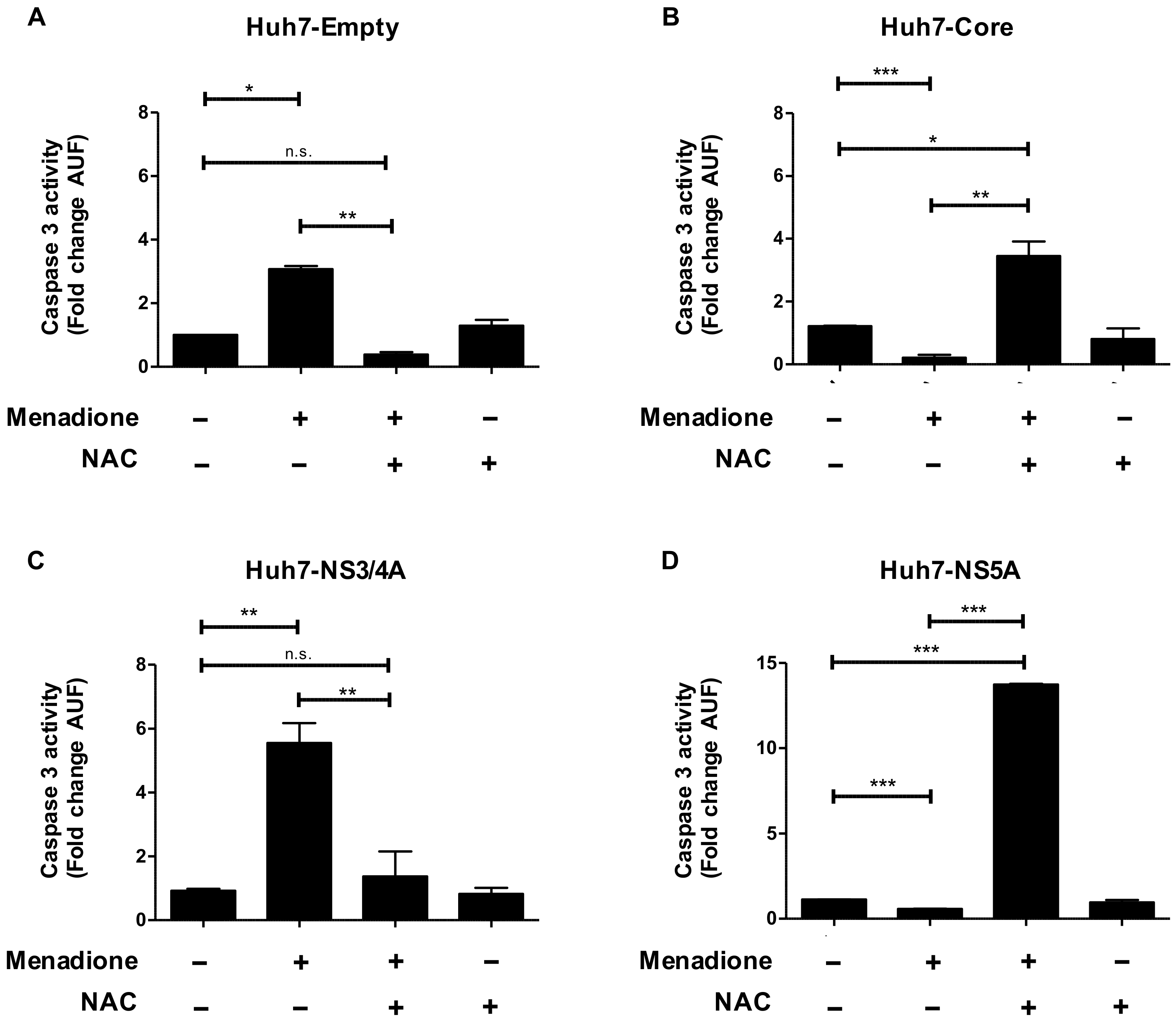
19]. In the present study, using stably transfected Huh7 cells, we confirm increased resistance to oxidative stress and the selective degradation of HCV proteins Core and NS5A, as well as the activation of the stress pathway eIF2α/ATF4. Although the infectious HCV culture system (HCVcc) was established in 2005 and is widely used in the field for the study of the cellular responses to HCV infection, we decided to use a simplified version of the system, since our aim was to focus on the exploration of adaptive mechanisms in hepatocyte-like cells exposed to multiple sources of damage and to investigate the role of individual HCV proteins in this adaptive response.
The increased resistance to oxidative stress, demonstrated by reduced apoptosis, observed in Huh7 cells expressing the HCV proteins Core and NS5A is probably not caused by altered levels of anti-oxidant enzymes. Gene expression of prominent anti-oxidant genes like SOD1 and CAT were not changed by any of the treatments, although we cannot rule out a regulation at the level of antioxidant enzyme activity.
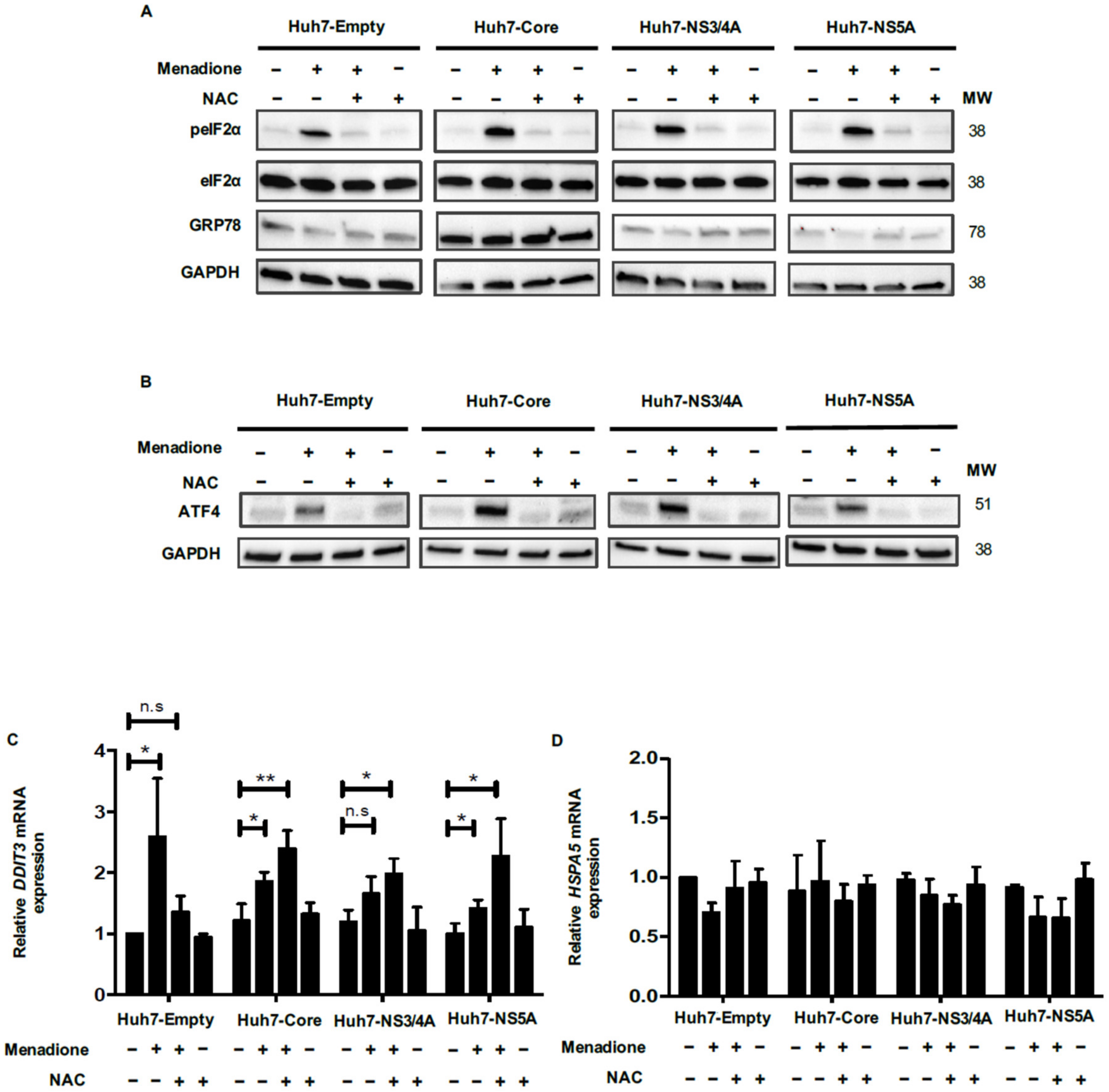
Autophagy has been described as an important adaptive survival mechanism to cope with cellular stress, e.g., ER stress and oxidative stress [
26]. The initial step in activation of the autophagic program is activation of the eIF2α/ATF4 pathway [
15,
27]. In our model, we were able to demonstrate activation of the eIF2α/ATF4 pathway and downstream events like increased ATF4 and CHOP (
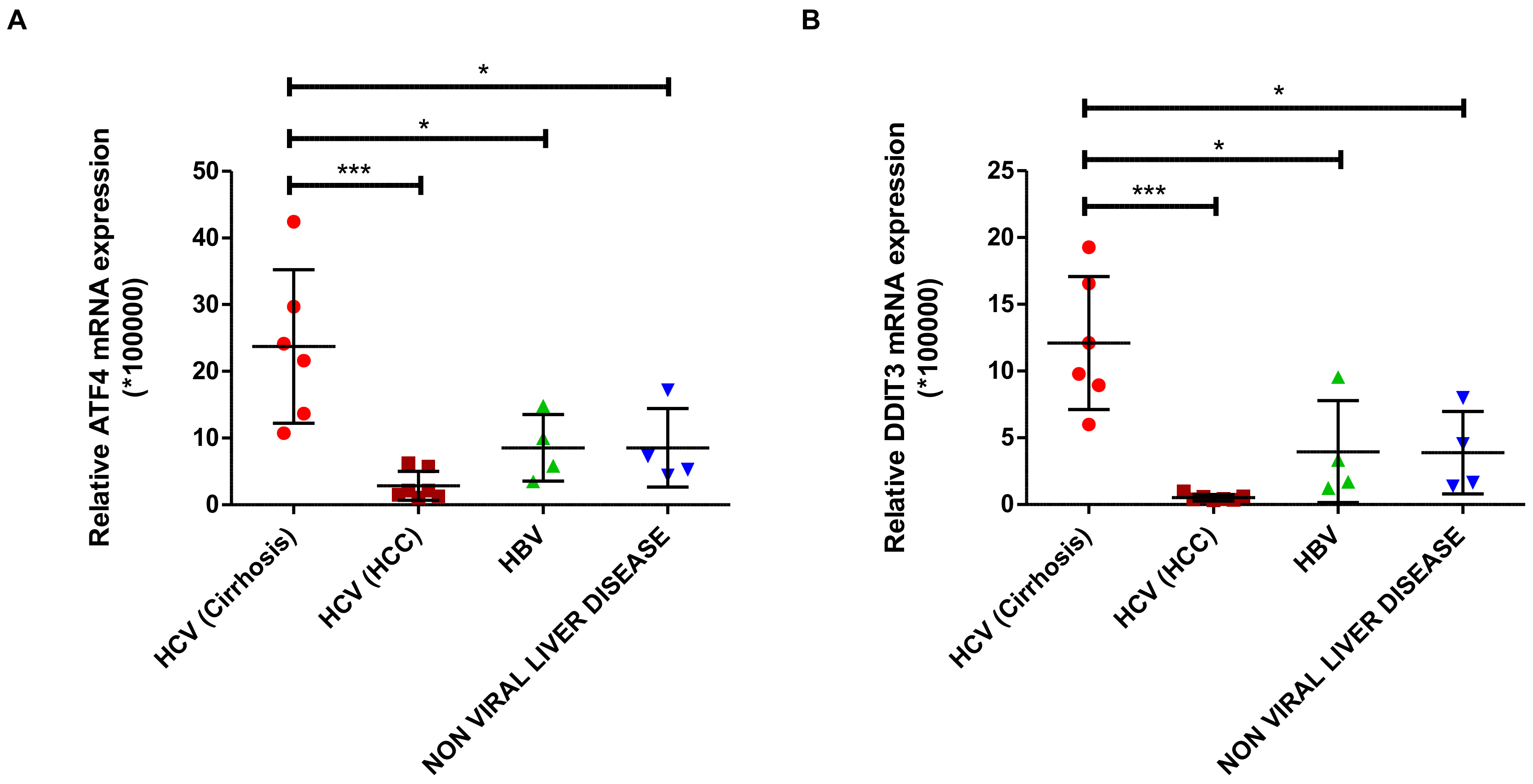
DDIT3) expression. Increased levels of autophagy markers have also been observed in liver biopsies of patients with HCV infection and in cell culture models of HCV infection [
28,
29]. In our study, we demonstrate increased expression of the transcription factors ATF4 and CHOP (
DDIT3), key elements of the eIF2α/ATF4 pathway, in liver biopsies from patients with HCV-related cirrhosis, but not in liver tumor tissue of patients with HCV-related HCC or liver tissue of non-HCV-related cirrhosis. These clinical data support our experimental data and also highlight the specificity of the observed changes for HCV during its replication at early stages of the infection and during cirrhosis but not for late stages of the infection as HCC because HCV replication was considered low.
Sir et al. demonstrated that HCV infection induces the accumulation of autophagosomes in cells without increasing autophagic protein degradation, whereas accumulation of autophagosomes and protein degradation were increased in Huh7 cells under starvation, the ‘gold standard’ for induction of autophagy [
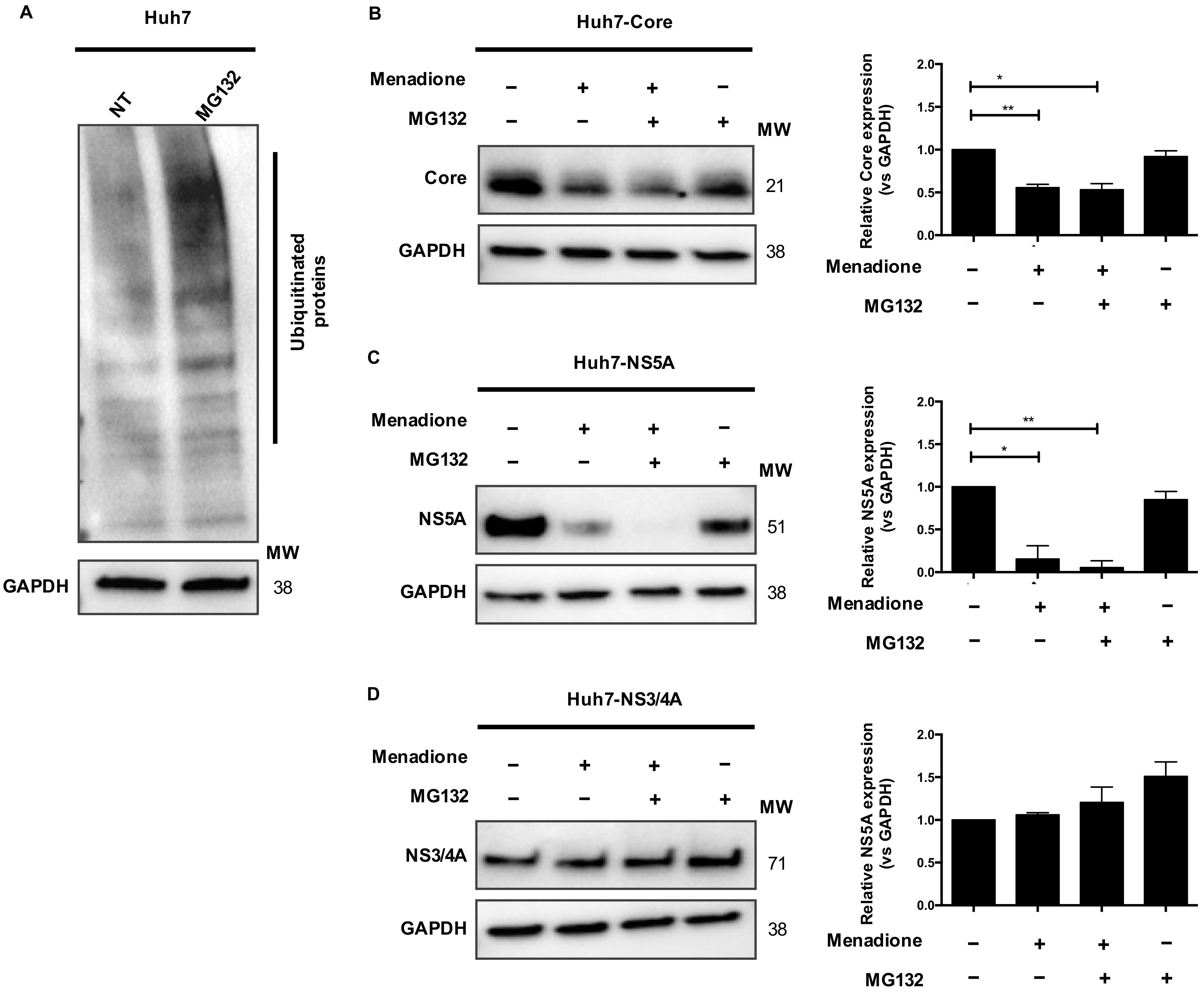
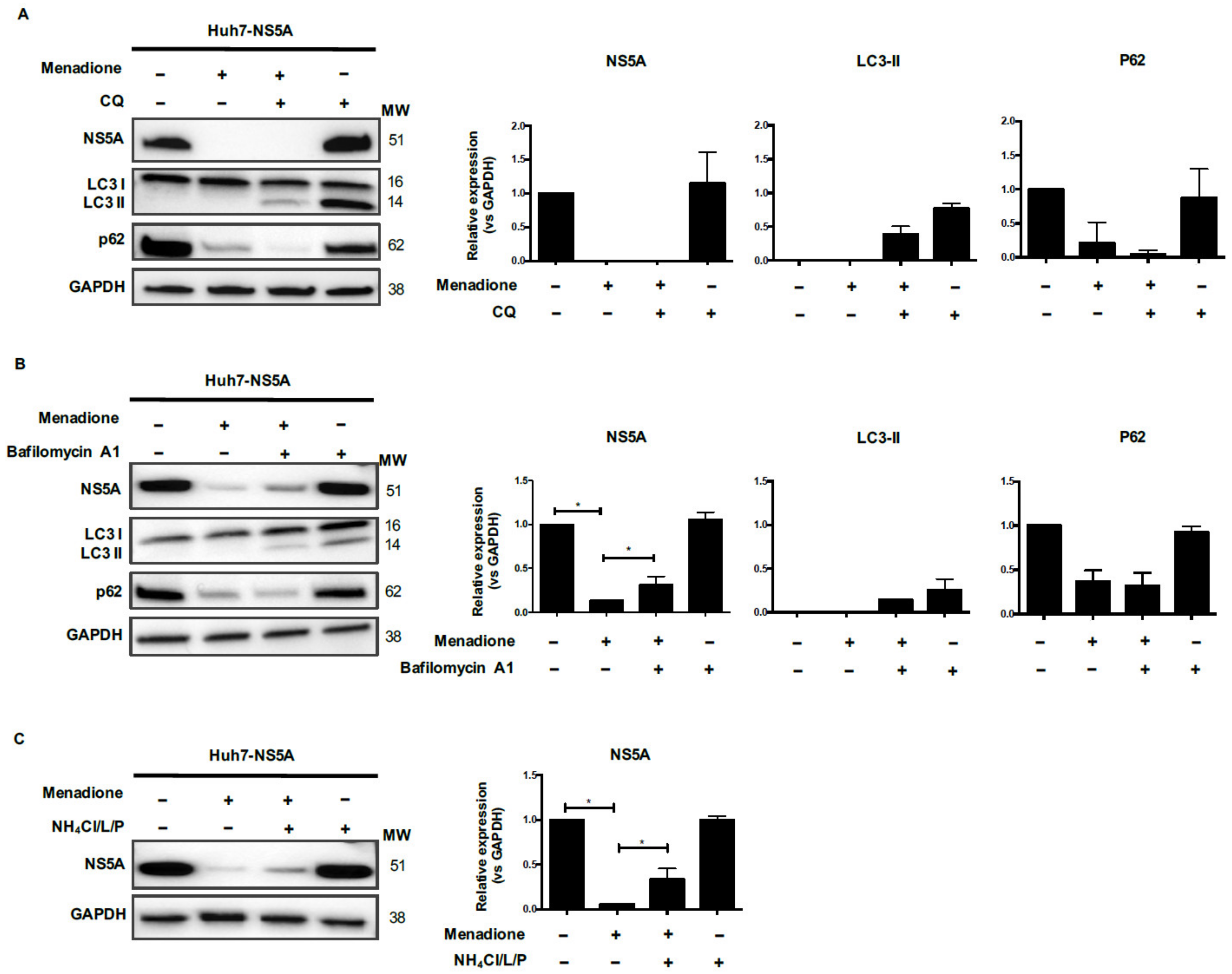
30]. We did observe increased viral protein degradation in HCV Core and NS5A-expressing Huh7 cells after exposure to external oxidative stress. However, this degradation was independent of the ubiquitin-proteasomal degradation pathway and was also not prevented by blocking autophagic flux with chloroquine. Blocking the lysosomal pathway partially restored HCV protein levels, however additional experiments are required to demonstrate the involvement of selective autophagic pathway and role of autophagy adaptor markers [
31].
Microautophagy, in which lysosomes invaginate and directly sequester cytosolic components, has been suggested to play an important role in the elimination of harmful proteins. In this pathway, LC3-II and p62 play an important role as adaptor proteins [
32]. In our study, we confirm that selective HCV Core and NS5A degradation can be induced in response to oxidative stress. The identification of autophagy receptor proteins such as p62, which also binds to ubiquitinated proteins, has provided a molecular link between the ubiquitination pathway and autophagy. p62 is a scaffold protein that has been implicated in processes like signal transduction, cell proliferation, cell survival, cell death and oxidative stress response [
33] and also plays an important role as receptor for selective autophagy [
34]. In our study, the HCV Core and NS5A degradation occurred simultaneously with a decrease of p62 protein level, suggesting the involvement of p62 as receptor protein for degradation of viral proteins after oxidative stress induction. Supporting this hypothesis is our observation that silencing of p62 allows the recovery of the expression of HCV Core and NS5A after external oxidative stress. Since we observed a partial restauration of NS5A expression after p62 silencing (
Figure 8D), other mechanism must also be considered. It is also important to demonstrate a direct interaction between HCV Core or NS5A with p62 by immunoprecipitation assay.
Wang et al. suggested that macroautophagy and chaperone-mediated autophagy (CMA) are required for hepatocyte resistance to oxidative stress, because inhibition of macroautophagy sensitized cells to apoptotic and necrotic cell death induced by menadione [
35]. Our results indicate a more prominent role for autophagy adaptor proteins like p62. The reason for this difference may be due to differences in the model systems used, but are in accordance with the suggestion of Czaja et al. who propose that two types of autophagy are better than one to face the effects of oxidative stress in hepatocytes and that adaptor proteins play an important role in this effect [
26].
In summary, our study demonstrates that hepatocyte-like cells can adapt to multiple stressors, like HCV protein expression and external oxidative stress. We conclude that activation of the eIF2α/ATF4 pathway and subsequent selective degradation, involving LC3-II and p62 contributes to the resistance of hepatocytes to oxidative stress by selective removal of one of the stressors (HCV proteins in this case). This mechanism suggests an important role for autophagy in viral replication and persistence of viral infection and may provide new leads for clinically applicable therapeutic interventions. In addition, it might be suggested to re-investigate in more detail the value of anti-oxidants in HCV patients, since anti-oxidants may abolish the adaptive response of hepatocytes and prevent the degradation of viral proteins in hepatocytes.


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}