Description of a Novel Mycovirus in the Phytopathogen Fusarium culmorum and a Related EVE in the Yeast Lipomyces starkeyi


Abstract
:1. Introduction
2. Materials and Methods
2.1. Recovery of FcV1 dsRNA
2.2. Full Genome Sequencing
2.3. Genome Sequence Analysis
2.4. Evaluation of Vertical Transmission
2.5. Chemical Treatment Assay
2.6. Horizontal Transmission Assay
2.7. Prevalence of FcV1 in Other F. culmorum Strains
3. Results
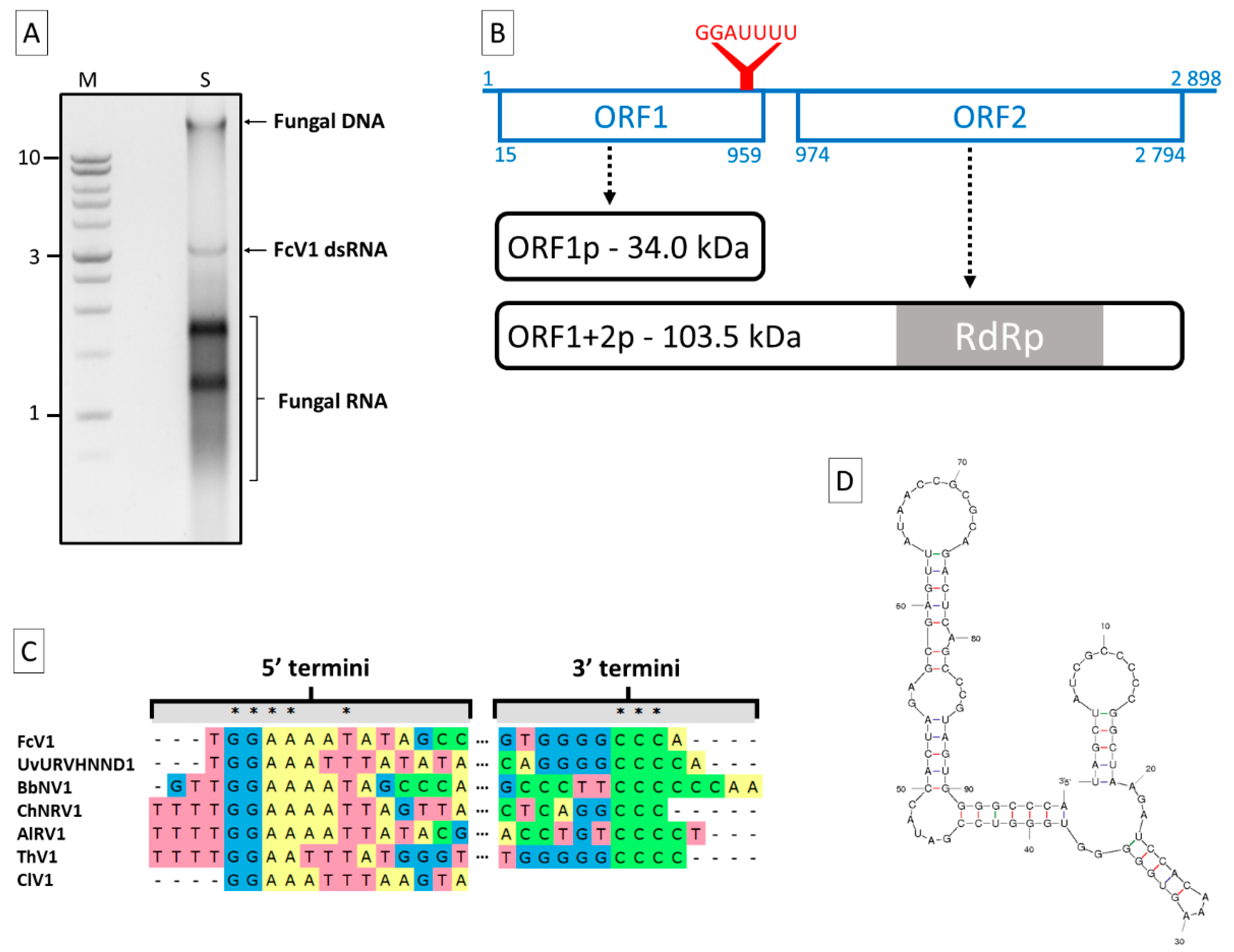
3.1. Genome Sequence Analysis of FcV1
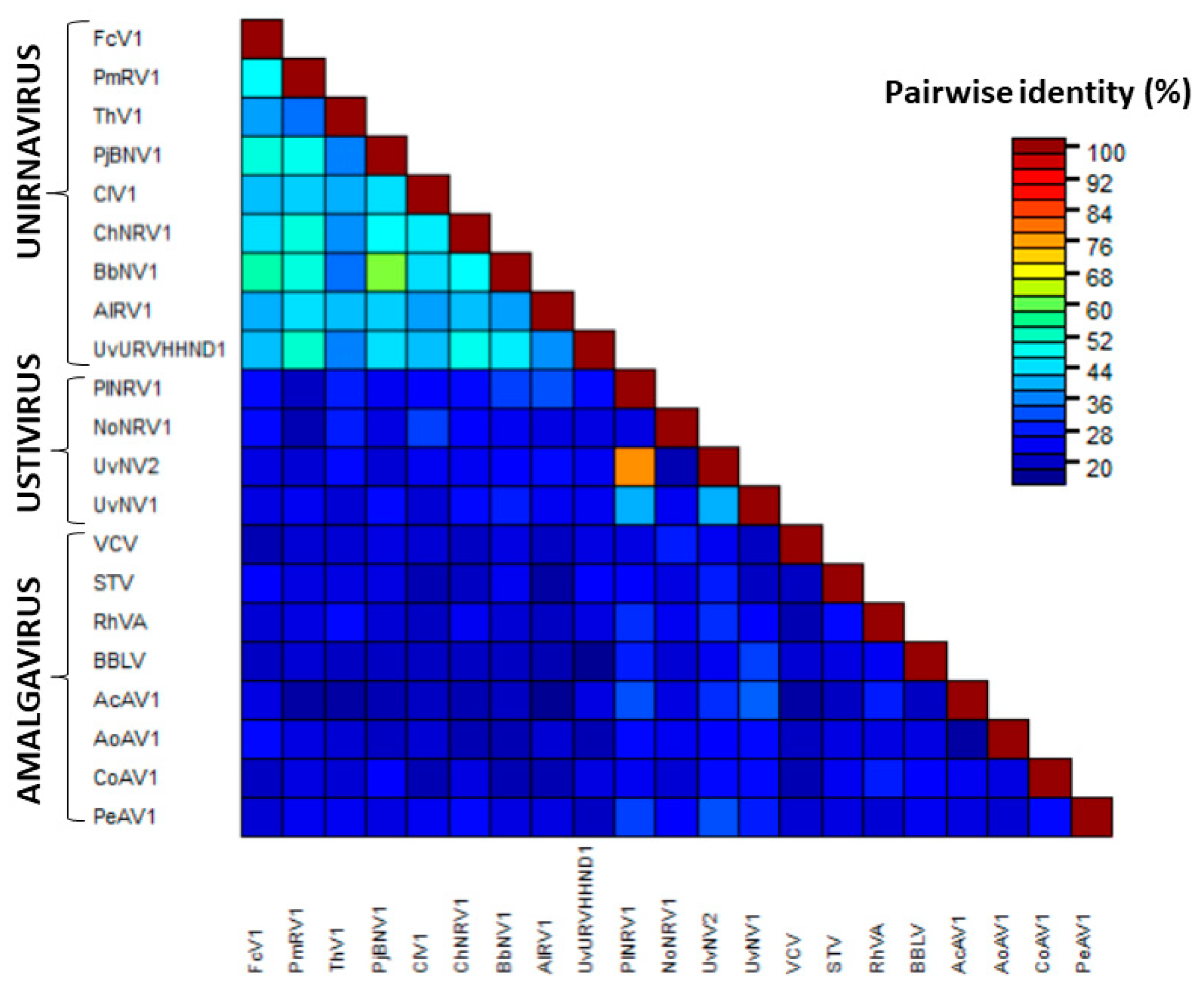
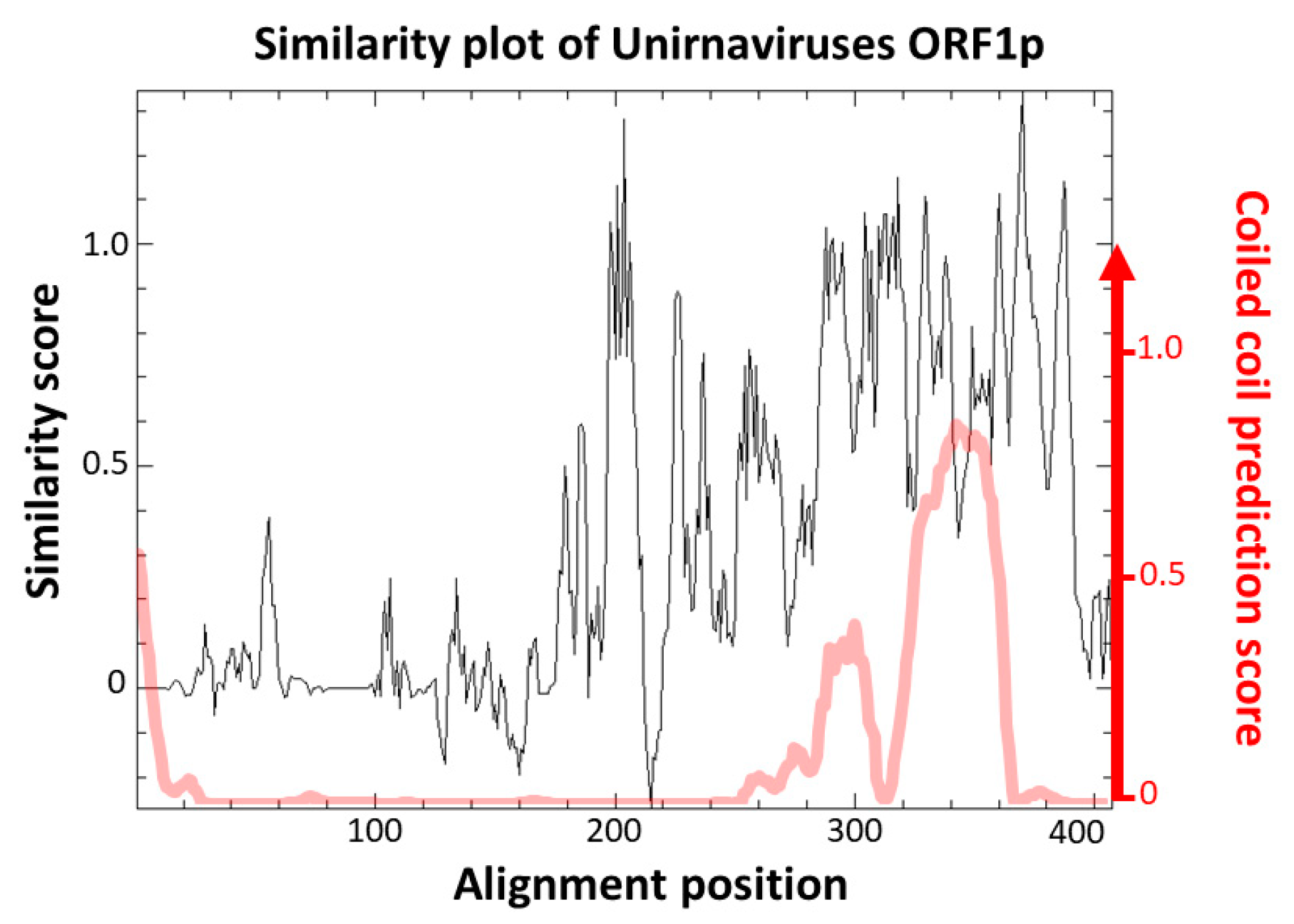
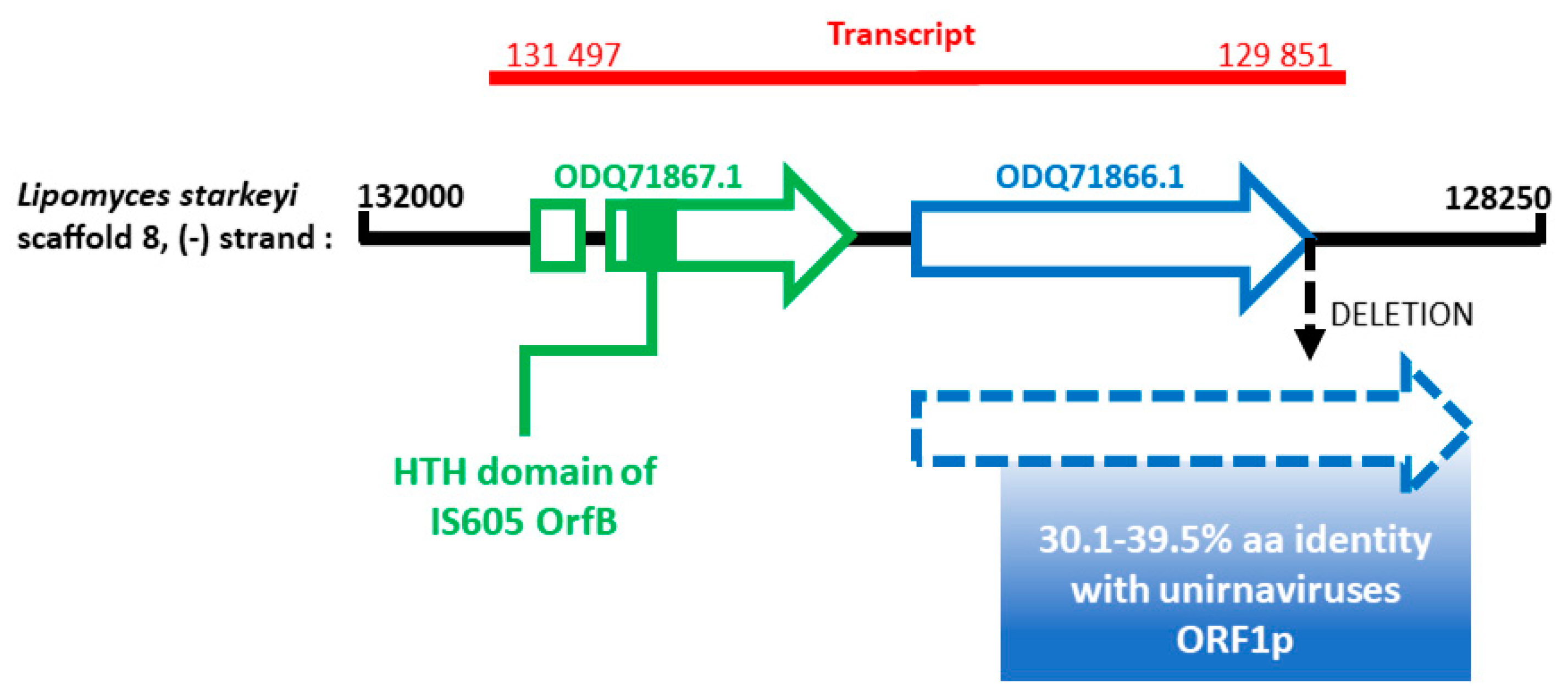
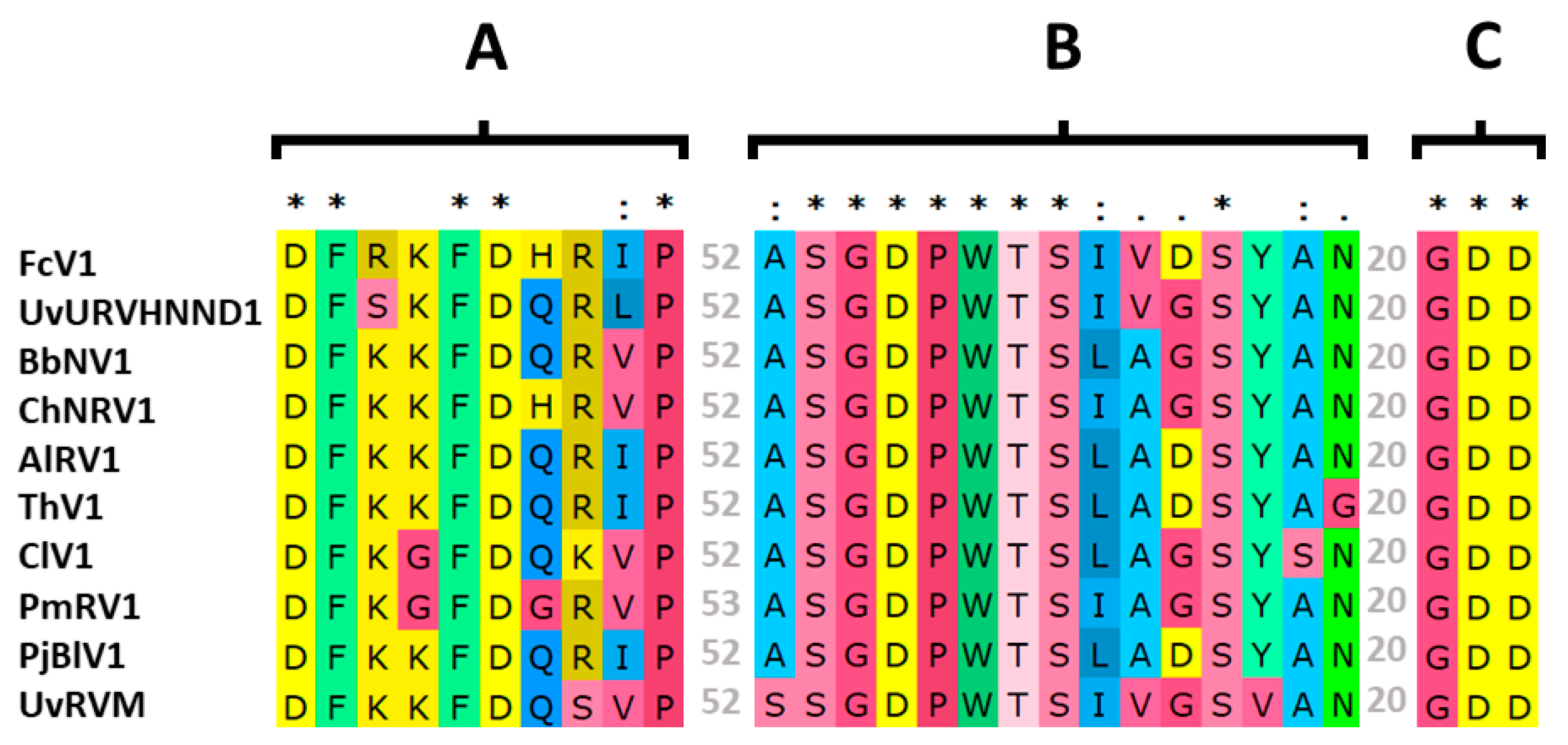
3.2. Analysis of FcV1 ORF1p and Retrieving of ORF1p Homologs
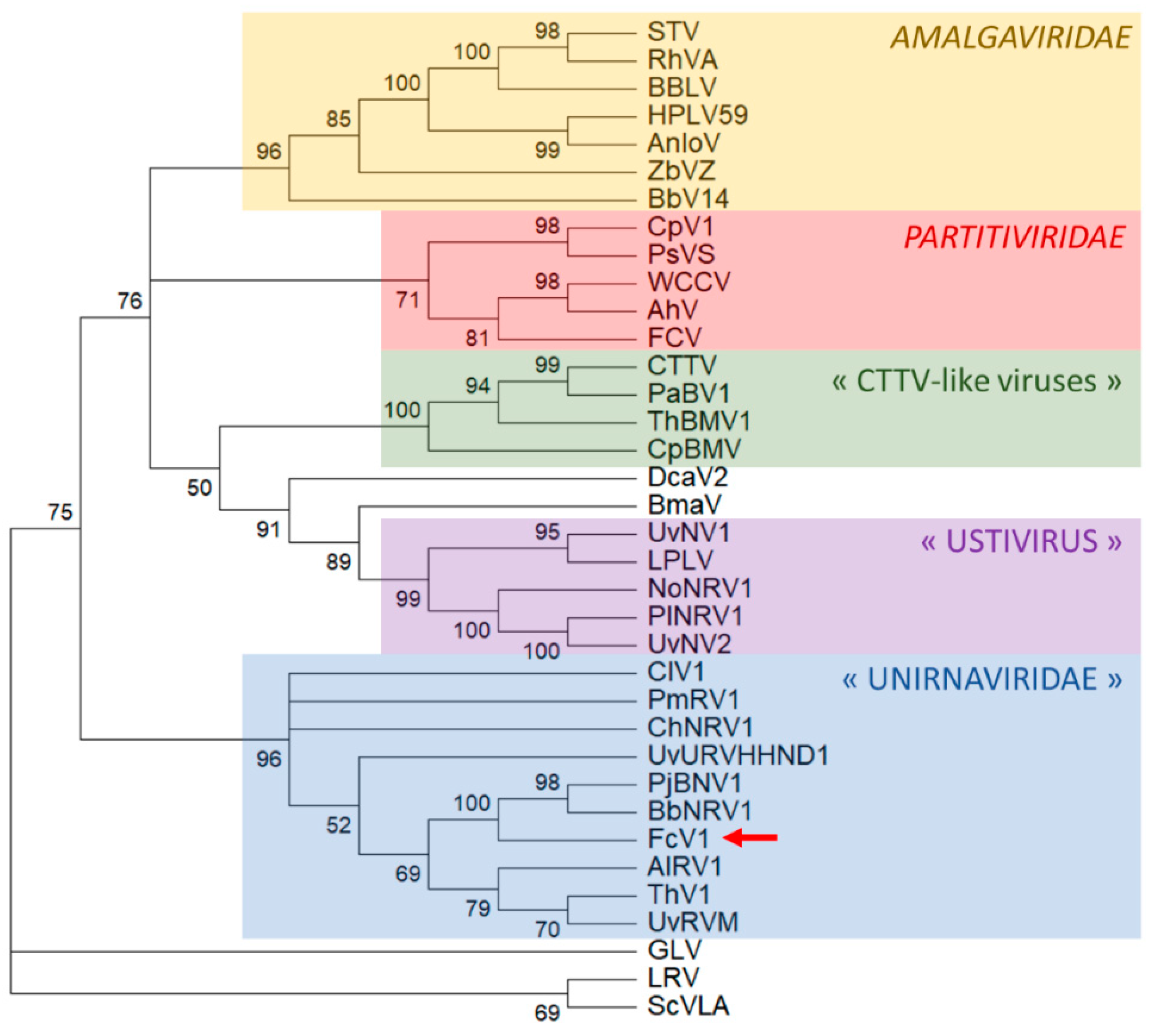
3.3. Analysis of FcV1 ORF2 and Phylogenetic Analysis
3.4. Biological Characterization and Prevalence in Other Strains
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Cho, W.K.; Lee, K.-M.; Yu, J.; Son, M.; Kim, K.-H. Insight into mycoviruses infecting Fusarium species. Adv. Virus Res. 2013, 86, 273–288. [Google Scholar] [CrossRef]
- Li, P.; Bhattacharjee, P.; Wang, S.; Zhang, L.; Ahmed, I.; Guo, L. Mycoviruses in Fusarium species: An update. Front. Cell Infect. Microbiol. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q. Fusarium culmorum: Causal agent of foot and root rot and head blight on wheat. Mol. Plant Pathol. 2013, 14, 323–341. [Google Scholar] [CrossRef]
- Hellin, P.; Dedeurwaerder, G.; Duvivier, M.; Scauflaire, J.; Huybrechts, B.; Callebaut, A.; Munaut, F.; Legrève, A. Relationship between Fusarium spp. diversity and mycotoxin contents of mature grains in southern Belgium. Food Addit. Contam. Part A 2016, 33, 1228–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Mello, J.P.F.; Placinta, C.M.; Macdonald, A.M.C. Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Anim. Feed Sci. Technol. 1999, 80, 183–205. [Google Scholar] [CrossRef] [Green Version]
- Hellin, P.; King, R.; Urban, M.; Hammond-Kosack, K.E.; Legrève, A. The adaptation of Fusarium culmorum to DMI fungicides is mediated by major transcriptome modifications in response to azole fungicide, including the overexpression of a PDR transporter (FcABC1). Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Jiang, D. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tóth, B.; Fónad, P.; Mesterházy, Á.; Varga, J. Double-stranded RNA mycoviruses in Fusarium culmorum and Fusarium graminearum isolates. Cereal Res. Commun. 2005, 33, 733–740. [Google Scholar] [CrossRef]
- Herrero, N.; Sánchez Márquez, S.; Zabalgogeazcoa, I. Mycoviruses are common among different species of endophytic fungi of grasses. Arch. Virol. 2009, 154, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Mahillon, M.; Decroës, A.; Liénard, C.; Bragard, C.; Legrève, A. Full genome sequence of a new polymycovirus infecting Fusarium Redolens. Arch. Virol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Khankhum, S.; Escalante, C.; de Souto and, E.R.; Valverde, R.A. Extraction and electrophoretic analysis of large dsRNAs from desiccated plant tissues infected with plant viruses and biotrophic fungi. Eur. J. Plant Pathol. 2017, 147, 431–441. [Google Scholar] [CrossRef]
- Wang, S.; Kondo, H.; Liu, L.; Guo, L.; Qiu, D. A novel virus in the family Hypoviridae from the plant pathogenic fungus Fusarium graminearum. Virus Res. 2013, 174, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Ghabrial, S.A. Molecular characterizations of two mitoviruses co-infecting a hyovirulent isolate of the plant pathogenic fungus Sclerotinia sclerotiorum. Virology 2012, 428, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Okonechnikov, K.; Golosova, O.; Fursov, M. UGENE team Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, 108277. [Google Scholar] [CrossRef] [PubMed]
- Ludwiczak, J.; Winski, A.; Szczepaniak, K.; Alva, V.; Dunin-Horkawicz, S. DeepCoil—A fast and accurate prediction of coiled-coil domains in protein sequences. Bioinformatics 2019. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, 200–204. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Depierreux, D.; Vong, M.; Nibert, M.L. Nucleotide sequence of Zygosaccharomyces bailii virus Z: Evidence for +1 programmed ribosomal frameshifting and for assignment to family Amalgaviridae. Virus Res. 2016, 217, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Campo, S.; Gilbert, K.B.; Carrington, J.C. Small RNA-based antiviral defense in the phytopathogenic fungus Colletotrichum higginsianum. PLoS Pathog. 2016, 12, 1005640. [Google Scholar] [CrossRef] [Green Version]
- Pyle, J.D.; Keeling, P.J.; Nibert, M.L. Amalga-like virus infecting Antonospora locustae, a microsporidian pathogen of grasshoppers, plus related viruses associated with other arthropods. Virus Res. 2017, 233, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Nerva, L.; Ciuffo, M.; Vallino, M.; Margaria, P.; Varese, G.C.; Gnavi, G.; Turina, M. Multiple approaches for the detection and characterization of viral and plasmid symbionts from a collection of marine fungi. Virus Res. 2016, 219, 22–38. [Google Scholar] [CrossRef]
- Kotta-Loizou, I.; Sipkova, J.; Coutts, R.H.A. Identification and sequence determination of a novel double-stranded RNA mycovirus from the entomopathogenic fungus Beauveria bassiana. Arch. Virol. 2015, 160, 873–875. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, H.; Zhao, C.; Liu, S.; Guo, L. The complete genome sequence of a novel mycovirus from Alternaria longipes strain HN28. Arch. Virol. 2015, 160, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.J.; Chen, D.; Zhong, J.; Zhang, S.Y.; Gao, B.D. A novel mycovirus identified from the rice false smut fungus Ustilaginoidea virens. Virus Genes 2015, 51, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhang, T.; Luo, C.; Jiang, D.; Li, G.; Li, Q.; Hsiang, T.; Huang, J. Prevalence and diversity of mycoviruses infecting the plant pathogen Ustilaginoidea virens. Virus Res. 2015, 195, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, M.; Redda, E.T.; Mei, J.; Zhang, J.; Wu, B.; Jiang, X. A novel double-stranded RNA mycovirus isolated from Trichoderma harzianum. Virol. J. 2019, 16, 113. [Google Scholar] [CrossRef] [PubMed]
- Nerva, L.; Turina, M.; Zanzotto, A.; Gardiman, M.; Gaiotti, F.; Gambino, G.; Chitarra, W. Isolation, molecular characterization and virome analysis of culturable wood fungal endophytes in esca symptomatic and asymptomatic grapevine plants. Environ. Microbiol. 2019, 21, 2886–2904. [Google Scholar] [CrossRef]
- Kotta-Loizou, I.; Coutts, R.H.A. Studies on the virome of the entomopathogenic fungus Beauveria bassiana reveal novel dsRNA elements and mild hypervirulence. PLoS Pathog. 2017, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nibert, M.L.; Pyle, J.D.; Firth, A.E. A +1 ribosomal frameshifting motif prevalent among plant amalgaviruses. Virology 2016, 498, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Krupovic, M.; Dolja, V.V.; Koonin, E.V. Plant viruses of the Amalgaviridae family evolved via recombination between viruses with double-stranded and negative-strand RNA genomes. Biol. Direct. 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Urayama, S.; Takaki, Y.; Nishi, S.; Yoshida-Takashima, Y.; Deguchi, S.; Takai, K.; Nunoura, T. Unveiling the RNA virosphere associated with marine microorganisms. Mol. Ecol. Resour. 2018, 18, 1444–1455. [Google Scholar] [CrossRef]
- Riley, R.; Haridas, S.; Wolfe, K.H.; Lopes, M.R.; Hittinger, C.T.; Göker, M.; Salamov, A.A.; Wisecaver, J.H.; Long, T.M.; Calvey, C.H.; et al. Comparative genomics of biotechnologically important yeasts. Proc. Natl. Acad. Sci. USA 2016, 113, 9882–9887. [Google Scholar] [CrossRef] [Green Version]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014, 42, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Bruenn, J.A. A structural and primary sequence comparison of the viral RNA-dependent RNA polymerases. Nucleic Acids Res. 2003, 31, 1821–1829. [Google Scholar] [CrossRef] [PubMed]
- Herrero, N. A novel monopartite dsRNA virus isolated from the entomopathogenic and nematophagous fungus Purpureocillium lilacinum. Arch. Virol. 2016, 161, 3375–3384. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lin, X.-D.; Tian, J.-H.; Chen, L.-J.; Chen, X.; Li, C.-X.; Qin, X.-C.; Li, J.; Cao, J.-P.; Eden, J.-S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Neville, P.; Nicholson, J.; Eden, J.-S.; Imrie, A.; Holmes, E.C. High-resolution metatranscriptomics reveals the ecological dynamics of mosquito-associated RNA viruses in Western Australia. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero, N.; Zabalgogeazcoa, I. Mycoviruses infecting the endophytic and entomopathogenic fungus Tolypocladium cylindrosporum. Virus Res. 2011, 160, 409–413. [Google Scholar] [CrossRef] [Green Version]
- Bhatti, M.F.; Jamal, A.; Petrou, M.A.; Cairns, T.C.; Bignell, E.M.; Coutts, R.H.A. The effects of dsRNA mycoviruses on growth and murine virulence of Aspergillus fumigatus. Fungal Genet. Biol. 2011, 48, 1071–1075. [Google Scholar] [CrossRef]
- Balmas, V.; Scherm, B.; Marcello, A.; Beyer, M.; Hoffmann, L.; Migheli, Q.; Pasquali, M. Fusarium species and chemotypes associated with fusarium head blight and fusarium root rot on wheat in Sardinia. Plant Pathol. 2015, 64, 972–979. [Google Scholar] [CrossRef] [Green Version]
- Mishra, P.K.; Fox, R.T.V.; Culham, A. Inter-simple sequence repeat and aggressiveness analyses revealed high genetic diversity, recombination and long-range dispersal in Fusarium culmorum. Ann. Appl. Biol. 2003, 143, 291–301. [Google Scholar] [CrossRef]
- Zhang, T.; Jiang, Y.; Dong, W. A novel monopartite dsRNA virus isolated from the phytopathogenic fungus Ustilaginoidea virens and ancestrally related to a mitochondria-associated dsRNA in the green alga Bryopsis. Virology 2014, 462–463, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Frank, A.C.; Wolfe, K.H. Evolutionary capture of viral and plasmid DNA by yeast nuclear chromosomes. Eukaryot Cell 2009, 8, 1521–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, D.J.; Bruenn, J. The evolution of novel fungal genes from non-retroviral RNA viruses. BMC Biol. 2009, 7, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Fu, Y.; Jiang, D.; Li, G.; Xie, J.; Cheng, J.; Peng, Y.; Ghabrial, S.A.; Yi, X. Widespread horizontal gene transfer from double-stranded RNA viruses to eukaryotic nuclear genomes. J. Virol. 2010, 84, 11876–11887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warner, B.E.; Ballinger, M.J.; Yerramsetty, P.; Reed, J.; Taylor, D.J.; Smith, T.J.; Bruenn, J.A. Cellular production of a counterfeit viral protein confers immunity to infection by a related virus. PeerJ 2018, 6, 5679. [Google Scholar] [CrossRef]
- Sabanadzovic, S.; Valverde, R.A.; Brown, J.K.; Martin, R.R.; Tzanetakis, I.E. Southern tomato virus: The link between the families Totiviridae and Partitiviridae. Virus Res. 2009, 140, 130–137. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Name | Abbreviation | Accession Number | Genome Size (bp) | 5′UTR (bp) | 3′UTR (bp) | ORF1p (aa) | ORF1+2p (aa) | Reference |
|---|---|---|---|---|---|---|---|---|
| Penicillium janczewskii Beauveria bassiana-like virus 1 | PjBlV1 | KT601106 | 2890 1 | 10 | 110 | 314 | -1 | [29] |
| Beauveria bassiana non-segmented virus 1 | BbNV1 | LN610699 | 3218 | 320 | 79 | 315 | 926 | [30] |
| Alternaria longipes dsRNA virus 1 | AlRV1 | KJ817371 | 3415 | 318 | 107 | 394 | 997 | [31] |
| Ustilaginoidea virens unassigned RNA virus HNND 1 | UvURVHNND1 | KR106133 | 2903 | 33 | 72 | 314 | 932 | [32] |
| Ustilaginoidea virens RNA virus M | UvRVM | KJ101567 | 2714 1 | -1 | -1 | -1 | -1 | [33] |
| Colletotrichum higginsianum non-segmented dsRNA virus 1 | ChNRV1 | KM923925 | 2923 | 16 | 106 | 324 | 933 | [27] |
| Trichoderma harzianum mycovirus 1 | ThV1 | MH155602 | 3160 | 113 | 111 | 379 | 979 | [34] |
| Combu-like dsRNA virus 1 | ClV1 | MH990637 | 2992 | 12 | 188 | 326 | 930 | Unpublished |
| Penicillium miczynskii RNA virus 1 | PmRV1 | MK584820 | 2891 | 226 | 134 | 238 | -2 | [35] |
| Fusarium culmorum virus 1 | FcV1 | MN187541 | 2898 | 14 | 106 | 314 | 926 | This study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahillon, M.; Romay, G.; Liénard, C.; Legrève, A.; Bragard, C. Description of a Novel Mycovirus in the Phytopathogen Fusarium culmorum and a Related EVE in the Yeast Lipomyces starkeyi. Viruses 2020, 12, 523. https://0-doi-org.brum.beds.ac.uk/10.3390/v12050523
Mahillon M, Romay G, Liénard C, Legrève A, Bragard C. Description of a Novel Mycovirus in the Phytopathogen Fusarium culmorum and a Related EVE in the Yeast Lipomyces starkeyi. Viruses. 2020; 12(5):523. https://0-doi-org.brum.beds.ac.uk/10.3390/v12050523
Chicago/Turabian StyleMahillon, Mathieu, Gustavo Romay, Charlotte Liénard, Anne Legrève, and Claude Bragard. 2020. "Description of a Novel Mycovirus in the Phytopathogen Fusarium culmorum and a Related EVE in the Yeast Lipomyces starkeyi" Viruses 12, no. 5: 523. https://0-doi-org.brum.beds.ac.uk/10.3390/v12050523