West Nile Virus Lineage 2 Vector Competence of Indigenous Culex and Aedes Mosquitoes from Germany at Temperate Climate Conditions

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Origin and Rearing
2.2. Mosquito Infection
2.3. Mosquito Processing
2.4. Vector Competence Indices
2.5. Data Analysis
2.6. Ethics Statement
3. Results
3.1. Feeding and Survival Rates
3.2. Vector Competence of Cx. pipiens and Ae. albopictus at 25 °C
3.3. Vector Competence of Cx. pipiens Biotype pipiens at Three Different Temperature Regimes (18 °C, 25 °C, and 28 °C)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The global ecology and epidemiology of West Nile virus. Biomed. Res. Int. 2015, 2015, 376230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komar, N.; Langevin, S.; Hinten, S.; Nemeth, N.; Edwards, E.; Hettler, D.; Davis, B.; Bowen, R.; Bunning, M. Experimental infection of North American birds with the New York 1999 strain of West Nile virus. Emerg. Infect. Dis. 2003, 9, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ramírez, E.; Llorente, F.; Jiménez-Clavero, M.A. Experimental infections of wild birds with West Nile virus. Viruses 2014, 6, 752–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smithburn, K.C.; Hughes, T.P.; Burke, A.W.; Paul, J.H. A neurotropic virus isolated from the blood of a native of Uganda. Am. J. Trop. Med. Hyg. 1940, 20, 471–492. [Google Scholar] [CrossRef]
- Owen, J.; Moore, F.; Panella, N.; Edwards, E.; Bru, R.; Hughes, M.; Komar, N. Migrating birds as dispersal vehicles for West Nile virus. Ecohealth 2006, 3, 79–85. [Google Scholar] [CrossRef]
- Asnis, D.S.; Conetta, R.; Teixeira, A.A.; Waldman, G.; Sampson, B.A. The West Nile virus outbreak of 1999 in New York: The flushing hospital experience. Clin. Infect. Dis. 2000, 30, 413–418. [Google Scholar] [CrossRef] [Green Version]
- McLean, R.G. West Nile Virus in North American Birds; USDA National Wildlife Research Center-Staff Publications: Fort Collins, CO, USA, 2006. [Google Scholar]
- Danis, K.; Papa, A.; Theocharopoulos, G.; Dougas, G.; Athanasiou, M.; Detsis, M.; Baka, A.; Lytras, T.; Mellou, K.; Bonovas, S.; et al. Outbreak of West Nile virus infection in Greece, 2010. Emerg. Infect. Dis. 2011, 17, 1868–1872. [Google Scholar] [CrossRef]
- Sirbu, A.; Ceianu, C.S.; Panculescu-Gatej, R.I.; Vázquez, A.; Tenorio, A.; Rebreanu, R.; Niedrig, M.; Nicolescu, G.; Pistol, A. Outbreak of West Nile virus infection in humans, Romania, July to October 2010. Eurosurveillance 2011, 16, 19762. [Google Scholar]
- Barzon, L.; Pacenti, M.; Franchin, E.; Pagni, S.; Lavezzo, E.; Squarzon, L.; Martello, T.; Russo, F.; Nicoletti, L.; Rezza, G.; et al. Large human outbreak of West Nile virus infection in north-eastern Italy in 2012. Viruses 2013, 5, 2825–2839. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, U.; Lühken, R.; Keller, M.; Cadar, D.; van der Grinten, E.; Michel, F.; Albrecht, K.; Eiden, M.; Rinder, M.; Lachmann, L.; et al. West Nile virus epizootic in Germany, 2018. Antivir. Res. 2019, 162, 39–43. [Google Scholar] [CrossRef]
- Ziegler, U.; Santos, P.D.; Groschup, M.H.; Hattendorf, C.; Eiden, M.; Höper, D.; Eisermann, P.; Keller, M.; Michel, F.; Klopfleisch, R.; et al. West Nile virus epidemic in Germany triggered by epizootic emergence, 2019. Viruses 2020, 12, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcus, I. West Nile Virus 48: Europe (Germany) ECDC. 2019. Available online: http://www.promedmail.org (accessed on 20 January 2020).
- Kilpatrick, A.M.; Kramer, L.D.; Campbell, S.R.; Alleyne, E.O.; Dobson, A.P.; Daszak, P. West Nile virus risk assessment and the bridge vector paradigm. Emerg. Infect. Dis. 2005, 11, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.J.; Sardelis, M.R.; Dohm, D.J.; O’Guinn, M.L. Potential North American vectors of West Nile virus. Ann. N. Y. Acad. Sci. 2001, 951, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Andreadis, T.G. The contribution of Culex pipiens complex mosquitoes to transmission and persistence of West Nile virus in North America. J. Am. Mosq. Control Assoc. 2012, 28, 137–151. [Google Scholar] [CrossRef]
- Vogels, C.B.; Göertz, G.P.; Pijlman, G.P.; Koenraadt, C.J. Vector competence of European mosquitoes for West Nile virus. Emerg. Microbes Infect. 2017, 6, e96. [Google Scholar] [CrossRef] [Green Version]
- Balenghien, T.; Vazeille, M.; Grandadam, M.; Schaffner, F.; Zeller, H.; Reiter, P.; Sabatier, P.; Fouque, F.; Bicout, D.J. Vector competence of some French Culex and Aedes mosquitoes for West Nile virus. Vector Borne Zoonotic Dis. 2008, 8, 589–595. [Google Scholar] [CrossRef]
- Fortuna, C.; Remoli, M.E.; Di Luca, M.; Severini, F.; Toma, L.; Benedetti, E.; Bucci, P.; Montarsi, F.; Minelli, G.; Boccolini, D.; et al. Experimental studies on comparison of the vector competence of four Italian Culex pipiens populations for West Nile virus. Parasites Vectors 2015, 8, 463. [Google Scholar] [CrossRef] [Green Version]
- Brustolin, M.; Talavera, S.; Santamaría, C.; Rivas, R.; Pujol, N.; Aranda, C.; Marquès, E.; Valle, M.; Verdún, M.; Pagès, N.; et al. Culex pipiens and Stegomyia albopicta (= Aedes albopictus) populations as vectors for lineage 1 and 2 West Nile virus in Europe. Med. Vet. Entomol. 2016, 30, 166–173. [Google Scholar] [CrossRef] [Green Version]
- Fros, J.J.; Miesen, P.; Vogels, C.B.; Gaibani, P.; Sambri, V.; Martina, B.E.; Koenraadt, C.J.; van Rij, R.P.; Vlak, J.M.; Takken, W.; et al. Comparative usutu and West Nile virus transmission potential by local Culex pipiens mosquitoes in north-western Europe. One Health 2015, 1, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Fros, J.J.; Geertsema, C.; Vogels, C.B.; Roosjen, P.P.; Failloux, A.B.; Vlak, J.M.; Koenraadt, C.J.; Takken, W.; Pijlman, G.P. West Nile virus: High transmission rate in north-western European mosquitoes indicates its epidemic potential and warrants increased surveillance. PLoS Negl. Trop. Dis. 2015, 9, e0003956. [Google Scholar] [CrossRef] [Green Version]
- Vogels, C.B.; Fros, J.J.; Göertz, G.P.; Pijlman, G.P.; Koenraadt, C.J. Vector competence of northern European Culex pipiens biotypes and hybrids for West Nile virus is differentially affected by temperature. Parasites Vectors 2016, 9, 393. [Google Scholar] [CrossRef] [Green Version]
- Vogels, C.B.F.; Göertz, G.P.; Pijlman, G.P.; Koenraadt, C.J.M. Vector competence of northern and southern European Culex pipiens pipiens mosquitoes for West Nile virus across a gradient of temperatures. Med. Vet. Entomol. 2017, 31, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Koenraadt, C.J.M.; Möhlmann, T.W.R.; Verhulst, N.O.; Spitzen, J.; Vogels, C.B.F. Effect of overwintering on survival and vector competence of the West Nile virus vector Culex pipiens. Parasites Vectors 2019, 12, 147. [Google Scholar] [CrossRef] [PubMed]
- Leggewie, M.; Badusche, M.; Rudolf, M.; Jansen, S.; Börstler, J.; Krumkamp, R.; Huber, K.; Krüger, A.; Schmidt-Chanasit, J.; Tannich, E.; et al. Culex pipiens and Culex torrentium populations from Central Europe are susceptible to West Nile virus infection. One Health 2016, 2, 88–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, S.; Heitmann, A.; Lühken, R.; Leggewie, M.; Helms, M.; Badusche, M.; Rossini, G.; Schmidt-Chanasit, J.; Tannich, E. Culex torrentium: A potent vector for the transmission of West Nile virus in Central Europe. Viruses 2019, 11, 492. [Google Scholar] [CrossRef] [Green Version]
- Becker, N.; Petrić, D.; Zgomba, M.; Boase, C.; Dahl, C.; Madon, M.; Kaiser, A. Mosquitoes and Their Control, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Harbach, R.E.; Harrison, B.A.; Gad, A.M. Culex (Culex) molestus Forskal (Diptera: Culicidae): Neotype 538 designation, description, variation, and taxonomic status. Proc. Entomol. Soc. Wash 1984, 86, 521–542. [Google Scholar]
- Kampen, H.; Schuhbauer, A.; Walther, D. Emerging mosquito species in Germany-a synopsis after 6 years of mosquito monitoring (2011–2016). Parasitol. Res. 2017, 116, 3253–3263. [Google Scholar] [CrossRef]
- Kuhlisch, C.; Kampen, H.; Walther, D. The Asian tiger mosquito Aedes albopictus (Diptera: Culicidae) in central Germany: Surveillance in its northernmost distribution area. Acta Trop. 2018, 188, 78–85. [Google Scholar] [CrossRef]
- Rudolf, M.; Czajka, C.; Börstler, J.; Melaun, C.; Jöst, H.; von Thien, H.; Badusche, M.; Becker, N.; Schmidt-Chanasit, J.; Krüger, A.; et al. First nationwide surveillance of Culex pipiens complex and Culex torrentium mosquitoes demonstrated the presence of Culex pipiens biotype pipiens/molestus hybrids in Germany. PLoS ONE 2013, 8, e71832. [Google Scholar] [CrossRef]
- Eiden, M.; Viña-Rodríguez, A.; Hoffmann, B.; Ziegler, U.; Groschup, M.H. Two new real-time quantitative reverse transcription polymerase chain reaction assays with unique target sites for the specific and sensitive detection of lineages 1 and 2 West Nile virus strains. J. Vet. Diagn. Investig. 2010, 22, 748–753. [Google Scholar] [CrossRef] [Green Version]
- Viña-Rodríguez, A.; Sachse, K.; Ziegler, U.; Chaintoutis, S.C.; Keller, M.; Groschup, M.H.; Eiden, M. A novel pan-flavivirus detection and identification assay based on RT-qPCR and microarray. Biomed. Res. Int. 2017, 2017, e4248756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayr, A.; Bachmann, P.A.; Bibrack, B.; Wittmann, G. Virologische Arbeitsmethoden, (Zellkulturen-Bebrütete Hühnereier-Versuchstiere); VEB Gustav Fischer Verlag: Jena, Germany, 1974; Volume 1. [Google Scholar]
- Ziegler, U. Personal Communication; Friedrich-Loeffler-Institut: Greifswald-Insel Riems, Germany, 2019. [Google Scholar]
- Heitmann, A.; Jansen, S.; Lühken, R.; Leggewie, M.; Schmidt-Chanasit, J.; Tannich, E. Forced salivation as a method to analyze vector competence of mosquitoes. J. Vis. Exp. 2018, 138, e57980. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Development Core Team: Vienna, Austria, 2019. [Google Scholar]
- Lenth, R.V. Least-squares means: The R package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Harvey, W. Least-Squares Analysis of Data with Unequal Subclass Numbers; USDA National Agricultural Library: Washington, DC, USA, 1960; pp. 20–28. [Google Scholar]
- Tukey, W.J. The philosophy of multiple comparisons. Stat. Sci. 1991, 6, 100–116. [Google Scholar] [CrossRef]
- Deutscher Wetterdienst. Annual report 2018. In Yearbook 2018 of the Deutscher Wetterdienst; Deutscher Wetterdienst: Offenbach am Main, Germany, 2018; p. 16. [Google Scholar]
- Kampen, H.; Holicki, C.M.; Ziegler, U.; Groschup, M.H.; Tews, B.A.; Werner, D. West Nile virus mosquito vectors (Diptera: Culicidae) in Germany. Viruses 2020, 12, 493. [Google Scholar] [CrossRef]
- Tiawsirisup, S.; Platt, K.B.; Evans, R.B.; Rowley, W.A. A comparision of West Nile virus transmission by Ochlerotatus trivittatus (COQ.), Culex pipiens (L.), and Aedes albopictus (Skuse). Vector Borne Zoonotic Dis. 2005, 5, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Fortuna, C.; Remoli, M.E.; Severini, F.; Di Luca, M.; Toma, L.; Fois, F.; Bucci, P.; Boccolini, D.; Romi, R.; Ciufolini, M.G. Evaluation of vector competence for West Nile virus in Italian Stegomyia albopicta (= Aedes albopictus) mosquitoes. Med. Vet. Entomol. 2015, 29, 430–433. [Google Scholar] [CrossRef]
- Turell, M.J.; O’Guinn, M.L.; Dohm, D.J.; Jones, J.W. Vector competence of North American mosquitoes (Diptera: Culicidae) for West Nile virus. J. Med. Entomol. 2001, 38, 130–134. [Google Scholar] [CrossRef]
- Sardelis, M.R.; Turell, M.J.; O’Guinn, M.L.; Andre, R.G.; Roberts, D.R. Vector competence of three North American strains of Aedes albopictus for West Nile virus. J. Am. Mosq. Control Assoc. 2002, 18, 284–289. [Google Scholar]
- Magurano, F.; Remoli, M.E.; Baggieri, M.; Fortuna, C.; Marchi, A.; Fiorentini, C.; Bucci, P.; Benedetti, E.; Ciufolini, M.G.; Rizzo, C.; et al. Circulation of West Nile virus lineage 1 and 2 during an outbreak in Italy. Clin. Microbiol. Infect. 2012, 18, E545–E547. [Google Scholar] [CrossRef] [Green Version]
- Delatte, H.; Desvars, A.; Bouétard, A.; Bord, S.; Gimonneau, G.; Vourc’h, G.; Fontenille, D. Blood-feeding behavior of Aedes albopictus, a vector of chikungunya on la reunion. Vector Borne Zoonotic Dis. 2010, 10, 249–258. [Google Scholar] [CrossRef] [Green Version]
- Thomas, S.M.; Tjaden, N.B.; Frank, C.; Jaeschke, A.; Zipfel, L.; Wagner-Wiening, C.; Faber, M.; Beierkuhnlein, C.; Stark, K. Areas with high hazard potential for autochthonous transmission of Aedes albopictus-associated arboviruses in Germany. Int. J. Environ. Res. Public Health 2018, 15, 1270. [Google Scholar] [CrossRef] [Green Version]
- Deutscher Wetterdienst. The Weather in Germany in Summer 2019; Deutscher Wetterdienst: Offenbach am Main, Germany, 2019. [Google Scholar]
- Ewing, D.A.; Cobbold, C.A.; Purse, B.V.; Nunn, M.A.; White, S.M. Modelling the effect of temperature on the seasonal population dynamics of temperate mosquitoes. J. Biol. 2016, 400, 65–79. [Google Scholar] [CrossRef] [Green Version]
- Murdock, C.C.; Paaijmans, K.P.; Cox-Foster, D.; Read, A.F.; Thomas, M.B. Rethinking vector immunology: The role of environmental temperature in shaping resistance. Nat. Rev. Microbiol. 2012, 10, 869–876. [Google Scholar] [CrossRef] [Green Version]
- Vasseur, D.A.; DeLong, J.P.; Gilbert, B.; Greig, H.S.; Harley, C.D.G.; McCann, K.S.; Savage, V.; Tunney, T.D.; O’Connor, M.I. Increased temperature variation poses a greater risk to species than climate warming. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132612. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Mosquito Taxon | Collection Site | Year of Collection | Laboratory Colony | Field-Collected | Blood Source |
|---|---|---|---|---|---|
| Culex pipiens biotype pipiens | “Schöneiche“ and “Rehfelde” (S), Brandenburg, Germany | 2018 | X | Chicken | |
| “Groß Kreutz” (G), Brandenburg, Germany | 2019 | X | Chicken | ||
| Cx. pipiens biotype molestus | “Wendland” (W), Lower Saxony, Germany | 2012 | X | Bovine | |
| Novi Sad (N), Republic of Serbia | 2012 | X | Bovine | ||
| Aedes albopictus | Jena (J), Thuringia, Germany | 2016 | X | Bovine |
| Mosquito Species | Incubation Temperature (°C) | Feeding Rate (%) (95% CI) | Survival Rate 14/15 dpi (Excluding Day 0 Samples) (%) (95% CI) | Survival Rate 14/15–20/21 dpi (Excluding Day 0 Samples and Mosquitoes Tested 14/15 dpi) (%) (95% CI) |
|---|---|---|---|---|
| Culex pipiens biotype pipiens (S) | 25 | 48/58 (82.8) (70.6–91.4) | 41/46 (89.1) (76.4–96.4) | 17/21 (81.0) (58.1–94.6) |
| Cx. pipiens biotype pipiens (G) | 18 | 147/443 (33.2) (28.8–37.8) | 47/52 (90.4) (77.8–95.9) | 23/23 (100.0) (72.6–97.8) |
| 25 | 49/55 (89.1) (79.0–96.8) | 26/29 (89.7) (85.2–100.0) | ||
| 28 | 21/30 (70.0) (50.6–85.3) | 9/12 (75.0) (42.8–94.5) | ||
| Cx. pipiens biotype molestus (W) | 25 | 34/64 (53.1) (40.2–65.7) | 23/30 (76.7) (57.7–90.1) | 15/16 (93.8) (69.8–99.8) |
| Cx. pipiens biotype molestus (N) | 25 | 159/299 (53.2) (47.3–58.9) | 92/151 (60.9) (52.7–68.8) | 40/61 (65.6) (52.3–77.3) |
| Aedes albopictus (J) | 25 | 200/353 (56.7) (51.3–61.9) | 84/196 (42.9) (35.8–50.1) | 25/43 (58.1) (42.1–73.0) |
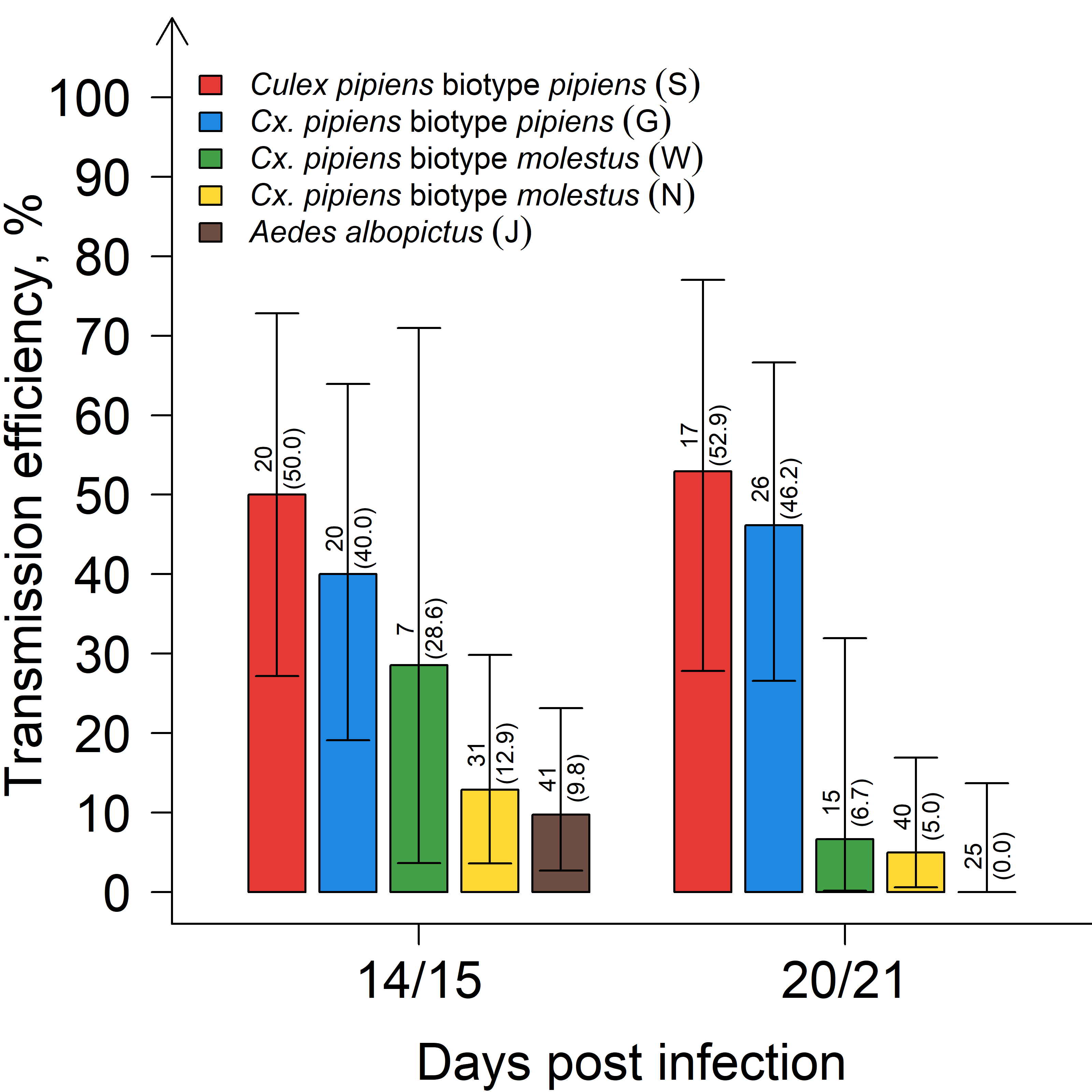
| Mosquito Species | Days Post Infection | Infection Rate (%) (95% CI) | Dissemination Rate (%) (95% CI) | Transmission Rate (%) (95% CI) | Transmission Efficiency (%) (95% CI) |
|---|---|---|---|---|---|
| Culex pipiens biotype pipiens (S) | 14/15 | 20/20 (100.0) (83.2–100.0) | 19/20 (95.0) (75.1–99.9) | 10/19 (52.6) (28.9–75.6) | 10/20 (50.0) (27.2–72.8) |
| 20/21 | 16/17 (94.1) (71.3–99.9) | 16/16 (100.0) (79.4–100.0) | 9/16 (60.0) (29.9–80.2) | 9/17 (52.9) (27.8–77.0) | |
| Cx. pipiens biotype pipiens (G) | 14/15 | 13/20 (65.0) (40.8–84.6) | 10/13 (76.9) (46.2–95.0) | 8/10 (80.0) (44.4–97.5) | 8/20 (40.0) (19.1–63.9) |
| 20/21 | 20/26 (76.9) (56.4–91.0) | 16/20 (80.0) (56.3–94.3) | 12/16 (75.0) (47.6–92.7) | 12/26 (46.2) (26.6–66.6) | |
| Cx. pipiens biotype molestus (W) | 14/15 | 7/7 (100.0) (59.0–100.0) | 6/7 (85.7) (42.1–99.6) | 2/6 (33.3) (4.3–77.7) | 2/7 (28.6) (3.7–71.0) |
| 20/21 | 1/15 (6.7) (0.2–31.9) | 1/1 (100.0) (2.5–100.0) | 1/1 (100.0) (2.5–100.0) | 1/15 (6.7) (0.2–31.9) | |
| Cx. pipiens biotype molestus (N) | 14/15 | 20/31 (64.5) (45.4–80.8) | 16/20 (80.0) (56.3–94.3) | 4/16 (25.0) (7.3–52.4) | 4/31 (12.9) (3.6–29.8) |
| 20/21 | 10/40 (15.0) (12.7–41.2) | 4/10 (40.0) (12.2–73.8) | 2/4 (50.0) (6.8–93.2) | 2/40 (5.0) (0.6–16.9) | |
| Aedes albopictus (J) | 14/15 | 4/41 (9.8) (2.7–23.1) | 4/4 (100.0) (39.8–100) | 4/4 (100.0) (39.8–100) | 4/41 (9.8) (2.7–23.1) |
| 20/21 | 5/25 (20.0) (6.8–40.7) | 4/5 (80.0) (28.4–99.5) | 0/4 (0.0) (0.0–60.2) | 0/25 (0.0) (0.0–13.7) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holicki, C.M.; Ziegler, U.; Răileanu, C.; Kampen, H.; Werner, D.; Schulz, J.; Silaghi, C.; Groschup, M.H.; Vasić, A. West Nile Virus Lineage 2 Vector Competence of Indigenous Culex and Aedes Mosquitoes from Germany at Temperate Climate Conditions. Viruses 2020, 12, 561. https://0-doi-org.brum.beds.ac.uk/10.3390/v12050561
Holicki CM, Ziegler U, Răileanu C, Kampen H, Werner D, Schulz J, Silaghi C, Groschup MH, Vasić A. West Nile Virus Lineage 2 Vector Competence of Indigenous Culex and Aedes Mosquitoes from Germany at Temperate Climate Conditions. Viruses. 2020; 12(5):561. https://0-doi-org.brum.beds.ac.uk/10.3390/v12050561
Chicago/Turabian StyleHolicki, Cora M., Ute Ziegler, Cristian Răileanu, Helge Kampen, Doreen Werner, Jana Schulz, Cornelia Silaghi, Martin H. Groschup, and Ana Vasić. 2020. "West Nile Virus Lineage 2 Vector Competence of Indigenous Culex and Aedes Mosquitoes from Germany at Temperate Climate Conditions" Viruses 12, no. 5: 561. https://0-doi-org.brum.beds.ac.uk/10.3390/v12050561