The Mutation of the Genes Related to Neurovirulence in HSV-2 Produces an Attenuated Phenotype in Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Mice and Ethics
2.3. Plasmid Construction and Transfection
2.4. Construction of Recombinant Mutant Viruses
2.5. Preliminary Analysis of the LAT-HSV-2, RL1-LAT-HSV-2 and HSV-2 Viruses
2.6. Experimental Infection of Mice
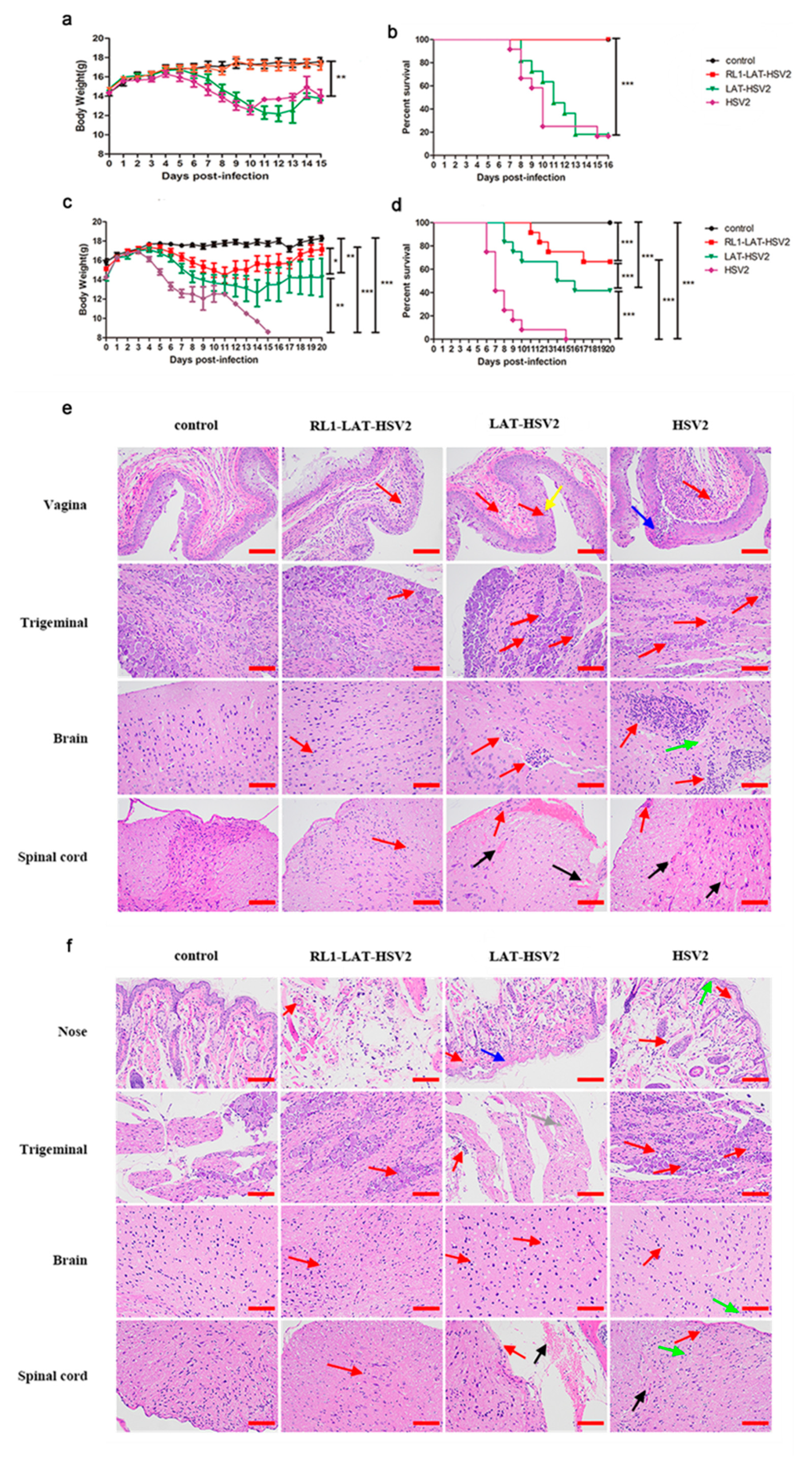
2.7. Histopathology
2.8. qRT-PCR
2.9. Reactivation Analysis of Dorsal Root Ganglia In Vitro
2.10. Flow Cytometry Analysis and ELISPOT (Enzyme-Linked ImmunoSpot)
2.11. Statistical Analysis
3. Results
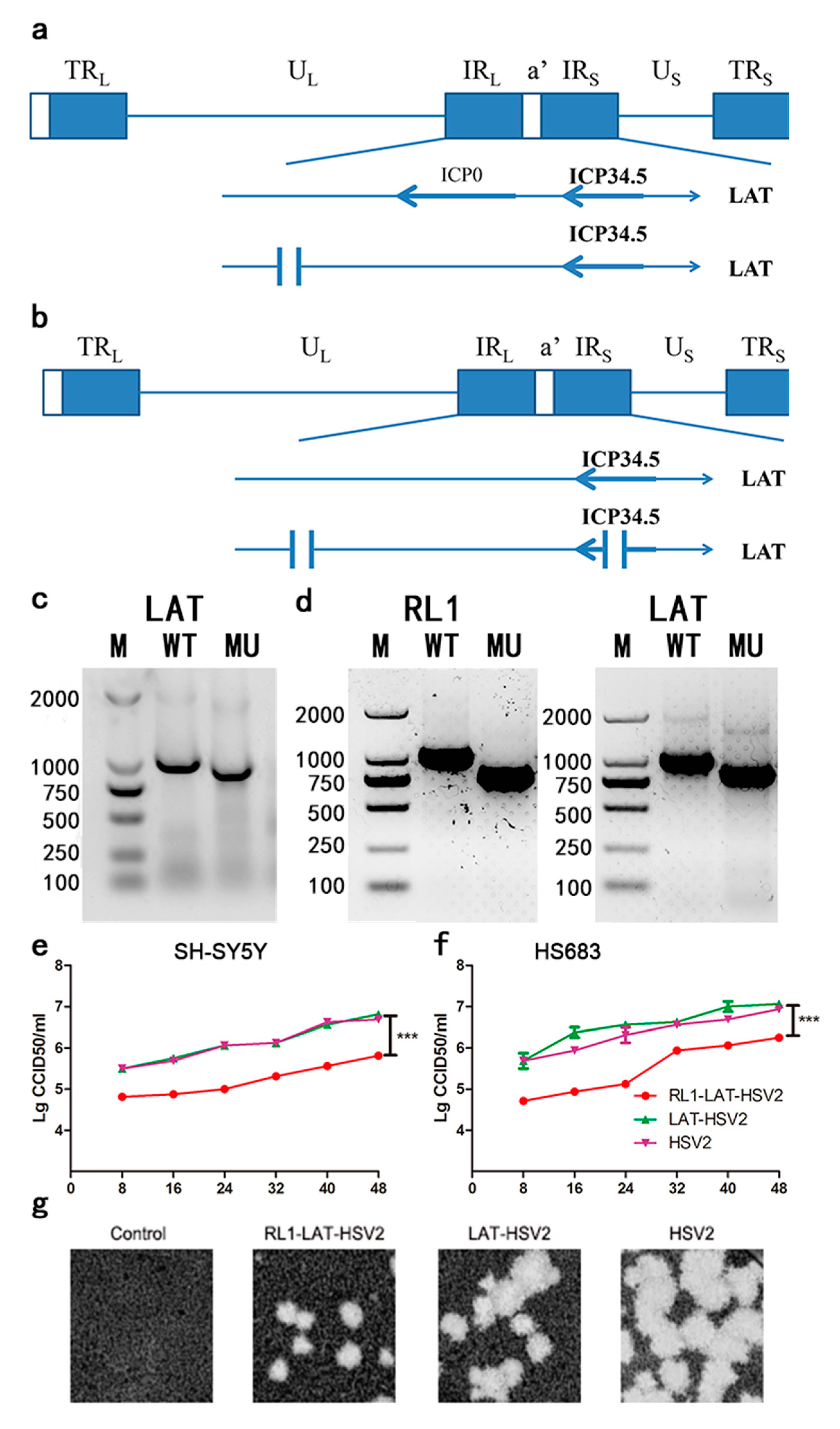
3.1. Modification and Preliminary Analysis of LAT-HSV-2 and RL1-LAT-HSV-2
3.2. Reduced Clinical Symptom Severity in RL1-LAT-HSV-2-Infected Mice
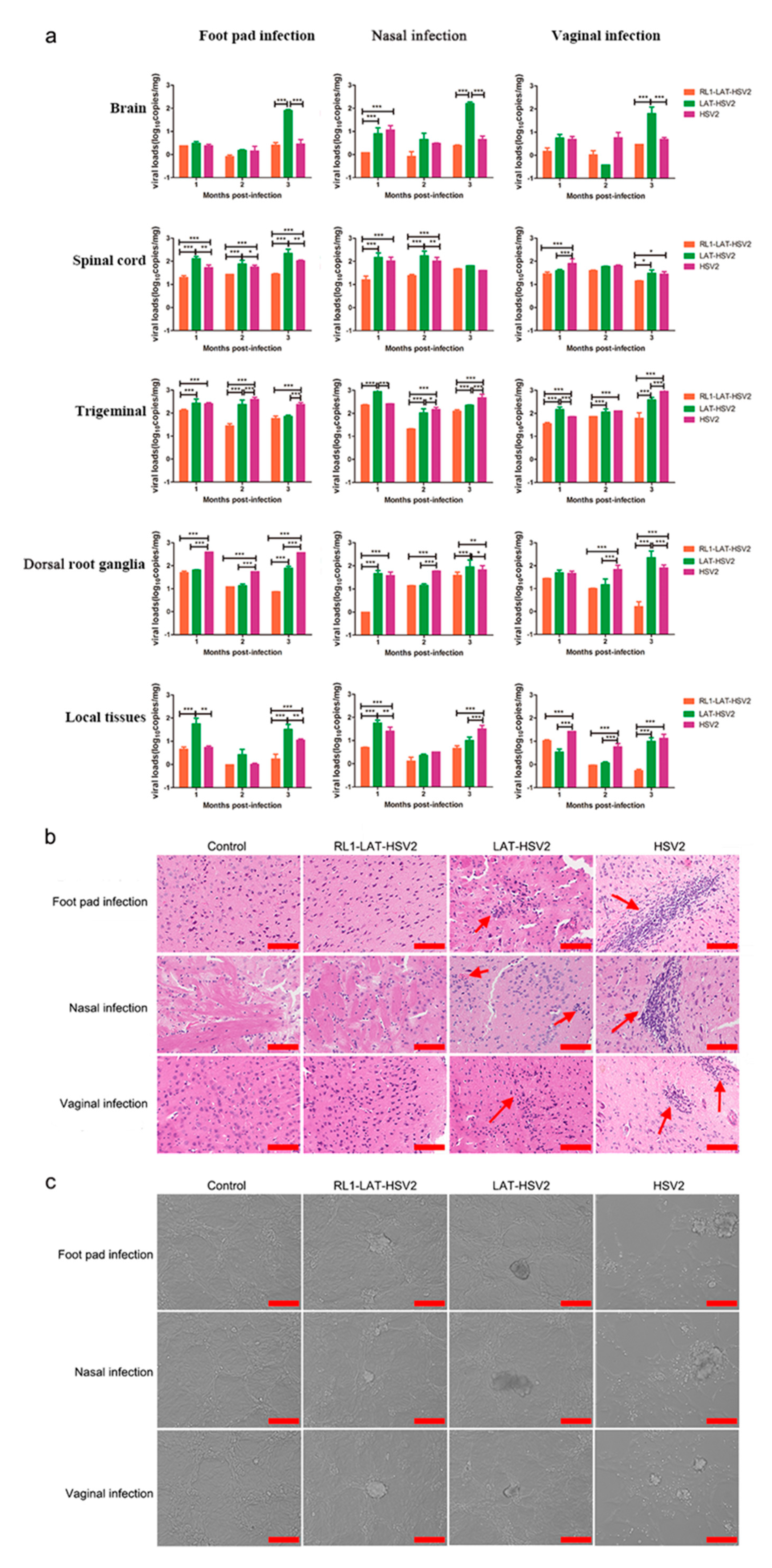
3.3. RL1-LAT-HSV-2 Displays Reduced Viral Proliferation in Mouse Tissues
3.4. The Latent Infection Capacity of RL1-LAT-HSV-2 Is Decreased
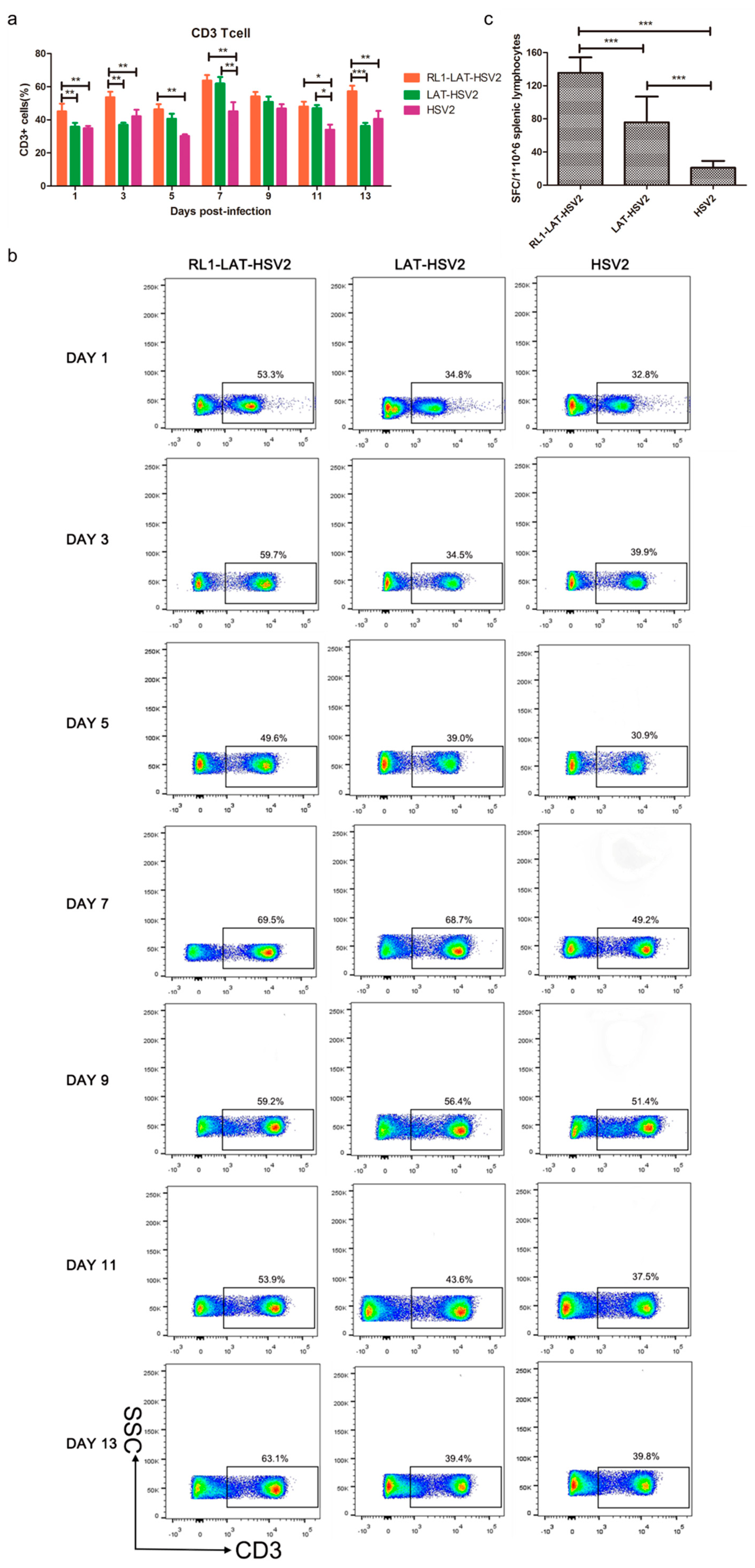
3.5. Increased Specific Immune Response Induced by RL1-LAT-HSV-2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kollias, C.M.; Huneke, R.B.; Wigdahl, B.; Jennings, S.R. Animal models of herpes simplex virus immunity and pathogenesis. J. Neurovirol. 2015, 21, 8–23. [Google Scholar] [CrossRef]
- Kawamura, Y.; Bosch-Marce, M.; Tang, S.; Patel, A.; Krause, P.R. Herpes Simplex Virus 2 Latency-Associated Transcript (LAT) Region Mutations Do Not Identify a Role for LAT-Associated MicroRNAs in Viral Reactivation in Guinea Pig Genital Models. J. Virol. 2018, 92, e00642-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafferty, W.E.; Coombs, R.W.; Benedetti, J.; Critchlow, C.; Corey, L. Recurrences after oral and genital herpes simplex virus infection. Influence of site of infection and viral type. N. Engl. J. Med. 1987, 316, 1444–1449. [Google Scholar] [CrossRef]
- Corey, L.; Wald, A.; Celum, C.L.; Quinn, T.C. The effects of herpes simplex virus-2 on HIV-1 acquisition and transmission: A review of two overlapping epidemics. J. Acquir. Immune Defic. Syndr. 2004, 35, 435–445. [Google Scholar] [CrossRef] [PubMed]
- James, S.H.; Kimberlin, D.W. Neonatal herpes simplex virus infection: Epidemiology and treatment. Clin. Perinatol. 2015, 42, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Yakoub, A.M.; Shukla, D. Basal Autophagy Is Required for Herpes simplex Virus-2 Infection. Sci. Rep. 2015, 5, 12985. [Google Scholar] [CrossRef] [PubMed]
- Brown, Z.A.; Selke, S.; Zeh, J.; Kopelman, J.; Maslow, A.; Ashley, R.L.; Watts, D.H.; Berry, S.; Herd, M.; Corey, L. The acquisition of herpes simplex virus during pregnancy. N. Engl. J. Med. 1997, 337, 509–515. [Google Scholar] [CrossRef]
- Brown, Z.A.; Wald, A.; Morrow, R.A.; Selke, S.; Zeh, J.; Corey, L. Effect of serologic status and cesarean delivery on transmission rates of herpes simplex virus from mother to infant. JAMA 2003, 289, 203–209. [Google Scholar] [CrossRef]
- Wald, A.; Link, K. Risk of human immunodeficiency virus infection in herpes simplex virus type-2-seropositive persons: A meta-analysis. J. Infect. Dis. 2002, 185, 45–52. [Google Scholar] [CrossRef]
- Barnabas, R.V.; Celum, C. Infectious co-factors in HIV-1 transmission herpes simplex virus type-2 and HIV-1: New insights and interventions. Curr. Hiv. Res. 2012, 10, 228–237. [Google Scholar] [CrossRef]
- Looker, K.J.; Elmes, J.A.R.; Gottlieb, S.L.; Schiffer, J.T.; Vickerman, P.; Turner, K.M.E.; Boily, M.C. Effect of HSV-2 infection on subsequent HIV acquisition: An updated systematic review and meta-analysis. Lancet Infect. Dis. 2017, 17, 1303–1316. [Google Scholar] [CrossRef] [Green Version]
- Byrne, C.M.; Gantt, S. Effects of spatiotemporal HSV-2 lesion dynamics and antiviral treatment on the risk of HIV-1 acquisition. PLoS Comput. Biol. 2018, 14, e1006129. [Google Scholar] [CrossRef] [PubMed]
- Corey, L.; Wald, A.; Patel, R.; Sacks, S.L.; Tyring, S.K.; Warren, T.; Douglas, J.M., Jr.; Paavonen, J.; Morrow, R.A.; Beutner, K.R.; et al. Once-daily valacyclovir to reduce the risk of transmission of genital herpes. N. Engl. J. Med. 2004, 350, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Fan, S.; Wang, X.; Hu, Y.; Feng, M.; Wang, L.; Zhang, Y.; Liao, Y.; Zhang, X.; Li, Q.; et al. Analysis of the Protective Immunity Induced by Herpes Simplex Virus 1 Strain M3 with an Attenuated Phenotype Due to Mutations in the Viral ul7, ul41, and LAT Genes. Front. Microbiol. 2017, 8, 1958. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Guo, Y.; Fan, S.; Cui, P.; Feng, M.; Wang, L.; Zhang, Y.; Liao, Y.; Zhang, X.; Li, Q.; et al. Attenuated phenotypes and analysis of a herpes simplex virus 1 strain with partial deletion of the UL7, UL41 and LAT genes. Virol. Sin. 2017, 32, 404–414. [Google Scholar] [CrossRef]
- Wilcox, D.R.; Longnecker, R. The Herpes Simplex Virus Neurovirulence Factor gamma34.5: Revealing Virus-Host Interactions. PLoS Pathog. 2016, 12, e1005449. [Google Scholar] [CrossRef] [Green Version]
- Chou, J.; Roizman, B. Herpes simplex virus 1 gamma(1)34.5 gene function, which blocks the host response to infection, maps in the homologous domain of the genes expressed during growth arrest and DNA damage. Proc. Natl. Acad. Sci. USA 1994, 91, 5247–5251. [Google Scholar] [CrossRef] [Green Version]
- Chou, J.; Kern, E.R.; Whitley, R.J.; Roizman, B. Mapping of herpes simplex virus-1 neurovirulence to gamma 134.5, a gene nonessential for growth in culture. Science 1990, 250, 1262–1266. [Google Scholar] [CrossRef]
- Stevens, J.G.; Wagner, E.K.; Devi-Rao, G.B.; Cook, M.L.; Feldman, L.T. RNA complementary to a herpesvirus alpha gene mRNA is prominent in latently infected neurons. Science 1987, 235, 1056–1059. [Google Scholar] [CrossRef]
- Gale, M., Jr.; Katze, M.G. Molecular mechanisms of interferon resistance mediated by viral-directed inhibition of PKR, the interferon-induced protein kinase. Pharmacol. Ther. 1998, 78, 29–46. [Google Scholar] [CrossRef]
- Cheng, G.; Gross, M.; Brett, M.E.; He, B. AlaArg motif in the carboxyl terminus of the gamma(1)34.5 protein of herpes simplex virus type 1 is required for the formation of a high-molecular-weight complex that dephosphorylates eIF-2alpha. J. Virol. 2001, 75, 3666–3674. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.; Bertke, A.S.; Patel, A.; Margolis, T.P.; Krause, P.R. Herpes simplex virus 2 microRNA miR-H6 is a novel latency-associated transcript-associated microRNA, but reduction of its expression does not influence the establishment of viral latency or the recurrence phenotype. J. Virol. 2011, 85, 4501–4509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, S.; Patel, A.; Krause, P.R. Novel less-abundant viral microRNAs encoded by herpes simplex virus 2 latency-associated transcript and their roles in regulating ICP34.5 and ICP0 mRNAs. J. Virol. 2009, 83, 1433–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, H.H.; Muhlbauer, G.; Rinner, B.; Stelzl, E.; Berger, A.; Dorr, H.W.; Santner, B.; Marth, E.; Rabenau, H. Detection of Herpes simplex virus DNA by real-time PCR. J. Clin. Microbiol. 2000, 38, 2638–2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, K.L.; Korom, M.; Morrison, L.A. Herpes simplex virus 2 ICP34.5 confers neurovirulence by regulating the type I interferon response. Virology 2014, 468, 330–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, S.M.; Harland, J.; MacLean, A.R.; Podlech, J.; Clements, J.B. Cell type and cell state determine differential in vitro growth of nonneurovirulent ICP34.5-negative herpes simplex virus types 1 and 2. J. Gen. Virol. 1994, 75, 2367–2377. [Google Scholar] [CrossRef]
- Zhang, X.; Jiang, Q.; Xu, X.; Wang, Y.; Liu, L.; Lian, Y.; Li, H.; Wang, L.; Zhang, Y.; Jiang, G.; et al. Immune mechanisms induced by an HSV-1 mutant strain: Discrepancy analysis of the immune system gene profile in comparison with a wild-type strain. Vaccine 2018, 36, 2394–2402. [Google Scholar] [CrossRef] [PubMed]
- Mott, K.R.; Bresee, C.J.; Allen, S.J.; BenMohamed, L.; Wechsler, S.L.; Ghiasi, H. Level of herpes simplex virus type 1 latency correlates with severity of corneal scarring and exhaustion of CD8+ T cells in trigeminal ganglia of latently infected mice. J. Virol. 2009, 83, 2246–2254. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Khanna, K.M.; Chen, X.; Fink, D.J.; Hendricks, R.L. CD8(+) T cells can block herpes simplex virus type 1 (HSV-1) reactivation from latency in sensory neurons. J. Exp. Med. 2000, 191, 1459–1466. [Google Scholar] [CrossRef] [Green Version]
- Rechenchoski, D.Z.; Faccin-Galhardi, L.C.; Linhares, R.E.C.; Nozawa, C. Herpesvirus: An underestimated virus. Folia Microbiol. 2017, 62, 151–156. [Google Scholar] [CrossRef]
- Minaya, M.A.; Jensen, T.L.; Goll, J.B.; Korom, M.; Datla, S.H.; Belshe, R.B.; Morrison, L.A. Molecular Evolution of Herpes Simplex Virus 2 Complete Genomes: Comparison between Primary and Recurrent Infections. J. Virol. 2017, 91, e00942-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.; Lu, G.; Qi, J.; Wu, L.; Tian, K.; Luo, T.; Shi, Y.; Yan, J. Structural basis of nectin-1 recognition by pseudorabies virus glycoprotein D. PLoS Pathog. 2017, 13, e1006314. [Google Scholar] [CrossRef] [Green Version]
- Cardone, G.; Heymann, J.B.; Cheng, N.; Trus, B.L.; Steven, A.C. Procapsid assembly, maturation, nuclear exit: Dynamic steps in the production of infectious herpesvirions. Adv. Exp. Med. Biol. 2012, 726, 423–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, C.; Gottlieb, S.L.; Wald, A. Status of vaccine research and development of vaccines for herpes simplex virus. Vaccine 2016, 34, 2948–2952. [Google Scholar] [CrossRef] [Green Version]
- Stanfield, B.; Kousoulas, K.G. Herpes Simplex Vaccines: Prospects of Live-attenuated HSV Vaccines to Combat Genital and Ocular infections. Curr. Clin. Microbiol. Rep. 2015, 2, 125–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kent, J.R.; Kang, W.; Miller, C.G.; Fraser, N.W. Herpes simplex virus latency-associated transcript gene function. J. Neurovirology 2003, 9, 285–290. [Google Scholar] [CrossRef]
- Bloom, D.C. HSV LAT and neuronal survival. Int. Rev. Immunol. 2004, 23, 187–198. [Google Scholar] [CrossRef]
- Koyanagi, N.; Imai, T.; Arii, J.; Kato, A.; Kawaguchi, Y. Role of herpes simplex virus 1 Us3 in viral neuroinvasiveness. Microbiol. Immunol. 2014, 58, 31–37. [Google Scholar] [CrossRef]
- Aubert, M.; Chen, Z.; Lang, R.; Dang, C.H.; Fowler, C.; Sloan, D.D.; Jerome, K.R. The antiapoptotic herpes simplex virus glycoprotein J localizes to multiple cellular organelles and induces reactive oxygen species formation. J. Virol. 2008, 82, 617–629. [Google Scholar] [CrossRef] [Green Version]
- Aubert, M.; Krantz, E.M.; Jerome, K.R. Herpes simplex virus genes Us3, Us5, and Us12 differentially regulate cytotoxic T lymphocyte-induced cytotoxicity. Viral Immunol. 2006, 19, 391–408. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Cheng, J.; Mou, T.; Zhang, Y.; Xu, X.; Zhang, J.; Li, X.; Feng, X.; Xu, X.; Liao, Y.; et al. The Mutation of the Genes Related to Neurovirulence in HSV-2 Produces an Attenuated Phenotype in Mice. Viruses 2020, 12, 770. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070770
Liu L, Cheng J, Mou T, Zhang Y, Xu X, Zhang J, Li X, Feng X, Xu X, Liao Y, et al. The Mutation of the Genes Related to Neurovirulence in HSV-2 Produces an Attenuated Phenotype in Mice. Viruses. 2020; 12(7):770. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070770
Chicago/Turabian StyleLiu, Lei, Jishuai Cheng, Tangwei Mou, Ying Zhang, Xingli Xu, Jingjing Zhang, Xueqi Li, Xiao Feng, Xiangxiong Xu, Yun Liao, and et al. 2020. "The Mutation of the Genes Related to Neurovirulence in HSV-2 Produces an Attenuated Phenotype in Mice" Viruses 12, no. 7: 770. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070770