Human Metapneumovirus Escapes NK Cell Recognition through the Downregulation of Stress-Induced Ligands for NKG2D


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Growth and Cell Lines
2.2. Flow Cytometry and Antibodies
2.3. Primers and Lentiviral Constructs Used for Cloning the HMPV Proteins
2.4. CD107a Degranulation Assay
2.5. RNA Extraction and cDNA Preparation
2.6. Quantitative Real-Time PCR
2.7. Western Blot Analysis
2.8. Proteasomal and Lysosomal Inhibitors
2.9. Statistical Analysis
3. Results
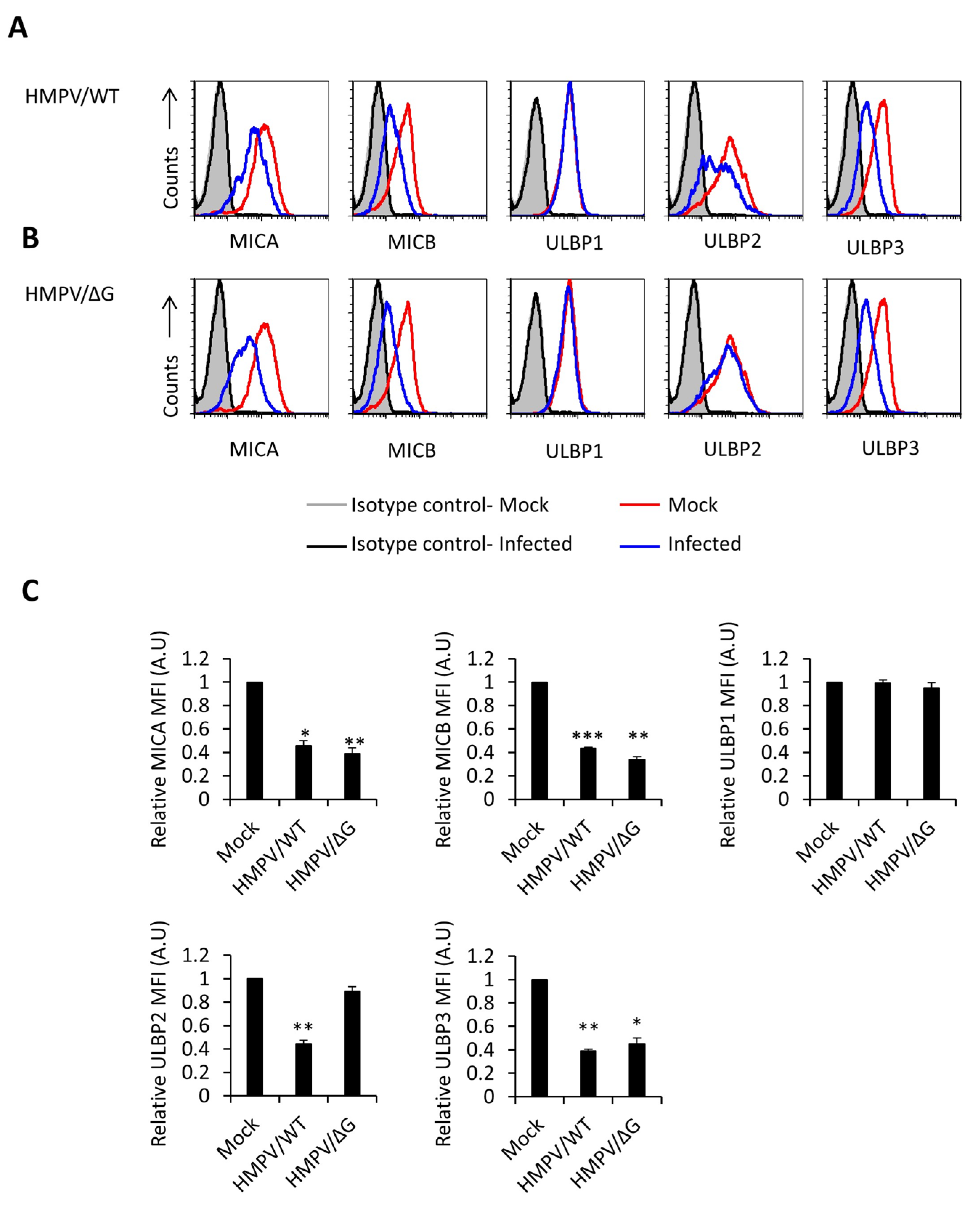
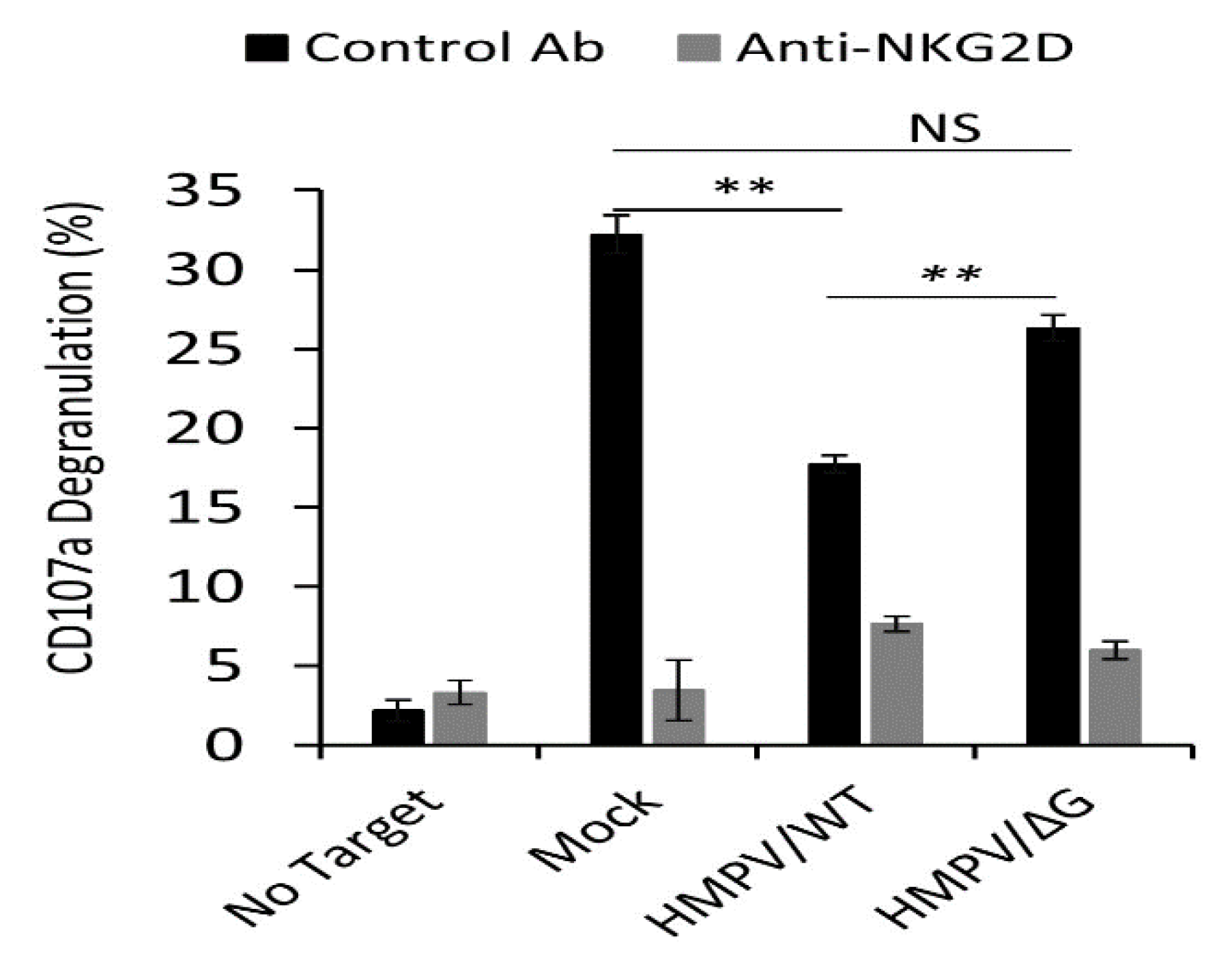
3.1. Ligands of NKG2D Receptor are Downregulated Following HMPV Infection Influencing NKG2D-Mediated Killing
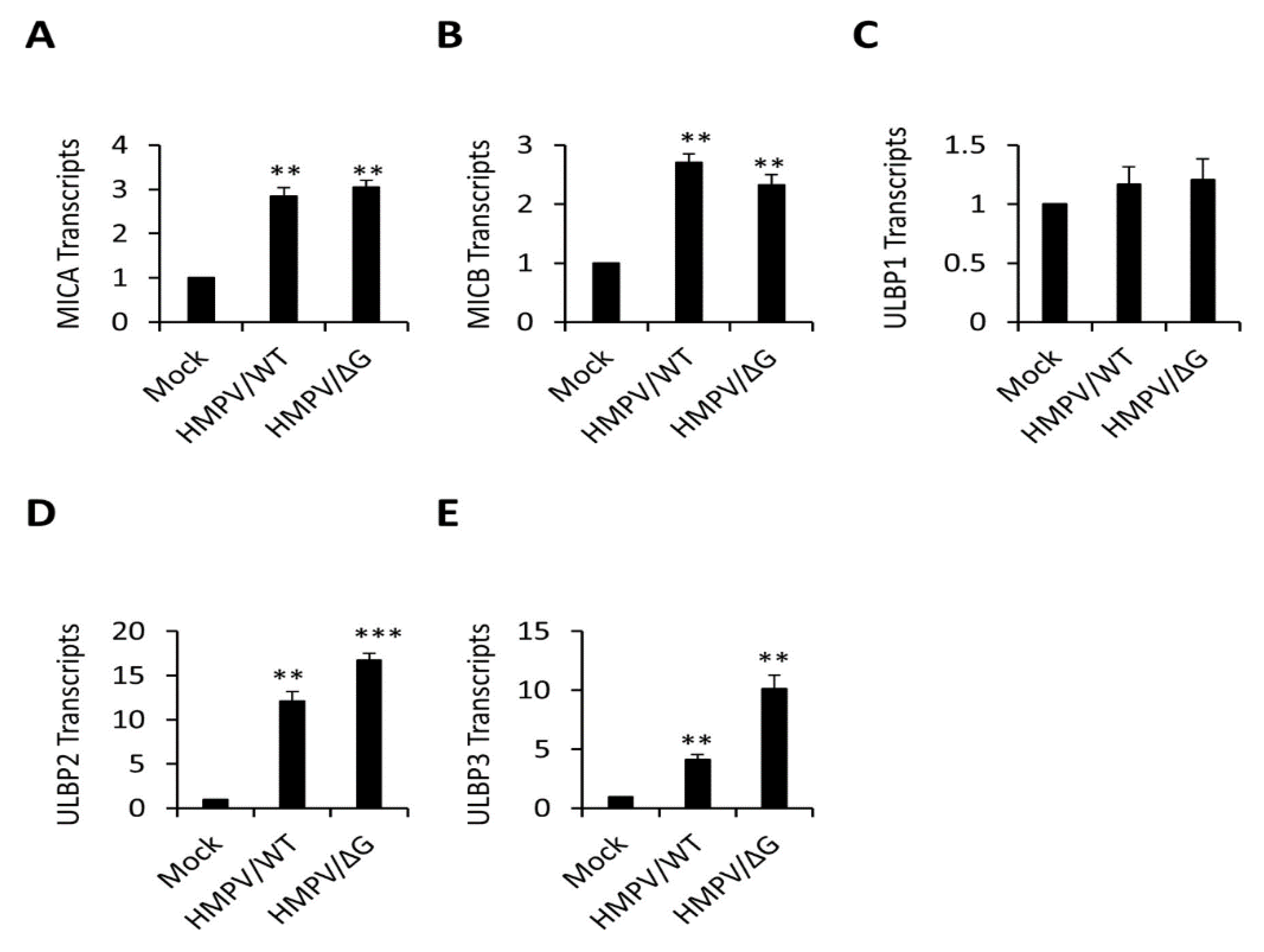
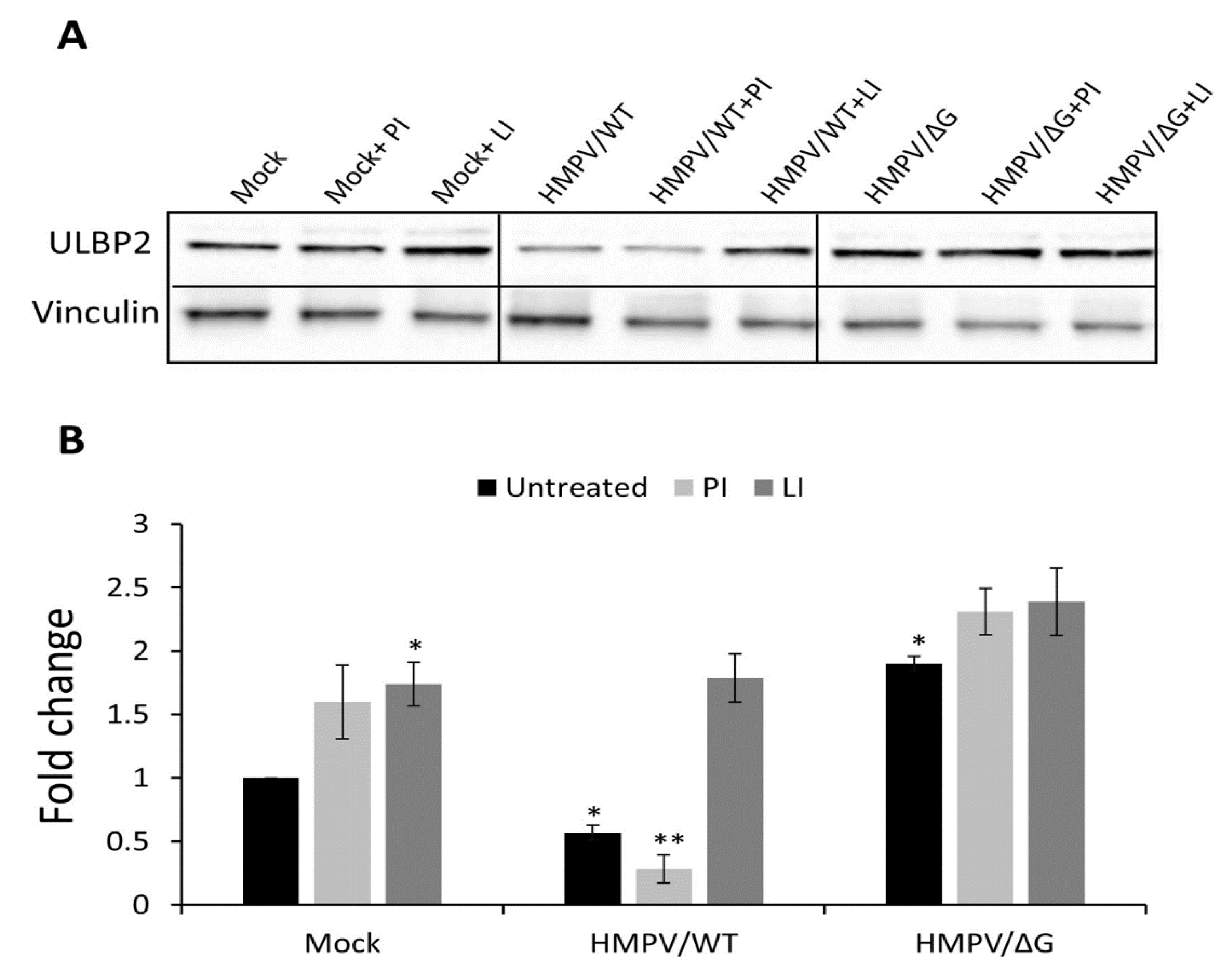
3.2. Mechanism of ULBP2 Downregulation
3.3. The HMPV Protein M2.2 Targets Major Histocompatibility Complex (MHC) Class I Polypeptide-Related Sequences A and B (MICA) and (MICB)
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arnon, T.I.; Markel, G.; Mandelboim, O. Tumor and viral recognition by natural killer cells receptors. Semin. Cancer Boil. 2006, 16, 348–358. [Google Scholar] [CrossRef]
- Cheent, K.; Khakoo, S.I. Natural killer cells: Integrating diversity with function. Immunology 2009, 126, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Lanier, L.L. Natural Killer Cells and Cancer; Academic Press: Cambridge, MA, USA, 2003; Volume 90, pp. 127–156. ISBN 0065-230X. [Google Scholar]
- Lodoen, M.B.; Lanier, L.L. Natural killer cells as an initial defense against pathogens. Curr. Opin. Immunol. 2006, 18, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Lanier, L.L. Up on the tightrope: Natural killer cell activation and inhibition. Nat. Immunol. 2008, 9, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Seidel, E.; Glasner, A.; Mandelboim, O. Virus-mediated inhibition of natural cytotoxicity receptor recognition. Cell. Mol. Life Sci. 2012, 69, 3911–3920. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Zimmermann, S.-Y.; Tramsen, L.; Koehl, U.; Lehrnbecher, T. Natural Killer Cells and Antifungal Host Response. Clin. Vaccine Immunol. 2013, 20, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Koch, J.; Steinle, A.; Watzl, C.; Mandelboim, O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol. 2013, 34, 182–191. [Google Scholar] [CrossRef]
- Pegram, H.J.; Andrews, D.M.; Smyth, M.J.; Darcy, P.K.; Kershaw, M. Activating, and inhibitory receptors of natural killer cells. Immunol. Cell Boil. 2010, 89, 216–224. [Google Scholar] [CrossRef]
- Raulet, D.H. Roles of the NKG2D immunoreceptor and its ligands. Nat. Rev. Immunol. 2003, 3, 781–790. [Google Scholar] [CrossRef]
- Pardoll, D.M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef] [Green Version]
- Obeidy, P.; Sharland, A.F. NKG2D and its ligands. Int. J. Biochem. Cell Boil. 2009, 41, 2364–2367. [Google Scholar] [CrossRef] [PubMed]
- Schmiedel, D.; Mandelboim, O. Disarming Cellular Alarm Systems—Manipulation of Stress-Induced NKG2D Ligands by Human Herpesviruses. Front. Immunol. 2017, 8, 2356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerwenka, A.; Lanier, L. NKG2D ligands: Unconventional MHC class I-like molecules exploited by viruses and cancer. Tiss. Antigens 2003, 61, 335–343. [Google Scholar] [CrossRef]
- Nachmani, D.; Stern-Ginossar, N.; Sarid, R.; Mandelboim, O. Diverse Herpesvirus MicroRNAs Target the Stress-Induced Immune Ligand MICB to Escape Recognition by Natural Killer Cells. Cell Host Microbe 2009, 5, 376–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonjić, S. Manipulation of NKG2D ligands by cytomegaloviruses: Impact on innate and adaptive immune response. Front. Immunol. 2011, 2, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauman, Y.; Drayman, N.; Ben-Nun-Shaul, O.; Vitenstein, A.; Yamin, R.; Ophir, Y.; Oppenheim, A.; Mandelboim, O. Downregulation of the stress-induced ligand ULBP1 following SV40 infection confers viral evasion from NK cell cytotoxicity. Oncotarget 2016, 7, 15369–15381. [Google Scholar] [CrossRef] [Green Version]
- Seidel, E.; Le-Trilling, V.T.K.; Bar-On, Y.; Tsukerman, P.; Enk, J.; Yamin, R.; Stein, N.; Schmiedel, D.; Djian, E.O.; Weisblum, Y.; et al. Dynamic Co-evolution of Host and Pathogen: HCMV Downregulates the Prevalent Allele MICA∗008 to Escape Elimination by NK Cells. Cell Rep. 2015, 10, 968–982. [Google Scholar] [CrossRef] [Green Version]
- Bar-On, Y.; Seidel, E.; Tsukerman, P.; Mandelboim, M.; Mandelboim, O. Influenza virus uses its neuraminidase protein to evade the recognition of two activating NK cell receptors. J. Infect. Dis. 2014, 210, 410–418. [Google Scholar] [CrossRef] [Green Version]
- Schmiedel, D.; Tai, J.; Levi-Schaffer, F.; Dovrat, S.; Mandelboim, O. Human Herpesvirus 6B Downregulates Expression of Activating Ligands during Lytic Infection to Escape Elimination by Natural Killer Cells. J. Virol. 2016, 90, 9608–9617. [Google Scholar] [CrossRef] [Green Version]
- Yamin, R.; Kaynan, N.S.; Glasner, A.; Vitenshtein, A.; Tsukerman, P.; Bauman, Y.; Ophir, Y.; Elias, S.; Bar-On, Y.; Gur, C.; et al. The Viral KSHV Chemokine vMIP-II Inhibits the Migration of Naive and Activated Human NK Cells by Antagonizing Two Distinct Chemokine Receptors. PLoS Pathog. 2013, 9, e1003568. [Google Scholar] [CrossRef]
- Bauman, Y.; Nachmani, D.; Vitenshtein, A.; Tsukerman, P.; Drayman, N.; Stern-Ginossar, N.; Lankry, D.; Gruda, R.; Mandelboim, O. An Identical miRNA of the Human JC and BK Polyoma Viruses Targets the Stress-Induced Ligand ULBP3 to Escape Immune Elimination. Cell Host Microbe 2011, 9, 93–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoogen, B.G.V.D.; de Jong, J.C.; Groen, J.; Kuiken, T.; de Groot, R.; Fouchier, R.A.; Osterhaus, A. A newly discovered human pneumovirus isolated from young children with respiratory tract disease. Nat. Med. 2001, 7, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Xepapadaki, P.; Psarras, S.; Bossios, A.; Tsolia, M.; Gourgiotis, D.; Liapi-Adamidou, G.; Constantopoulos, A.G.; Kafetzis, D.; Papadopoulos, N.G. Human metapneumovirus as a causative agent of acute bronchiolitis in infants. J. Clin. Virol. 2004, 30, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Falsey, A.R.; Erdman, D.; Anderson, L.J.; Walsh, E.E. Human Metapneumovirus Infections in Young and Elderly Adults. J. Infect. Dis. 2003, 187, 785–790. [Google Scholar] [CrossRef]
- Peiris, J.M.; Tang, W.-H.; Chan, K.-H.; Khong, P.-L.; Guan, Y.; Lau, Y.-L.; Chiu, S.S. Children with Respiratory Disease Associated with Metapneumovirus in Hong Kong. Emerg. Infect. Dis. 2003, 9, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Bastien, N.; Ward, D.; van Caeseele, P.; Brandt, K.; Lee, S.H.S.; McNabb, G.; Klisko, B.; Chan, E.; Li, Y. Human Metapneumovirus Infection in the Canadian Population. J. Clin. Microbiol. 2003, 41, 4642–4646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamelin, M.E.; Côtù, S.; la Forge, J.; Lampron, N.; Bourbeau, J.; Weiss, K.; Gilca, R.; de Serres, G.; Boivin, G. Human Metapneumovirus Infection in Adults with Community-Acquired Pneumonia and Exacerbation of Chronic Obstructive Pulmonary Disease. Clin. Infect. Dis. 2005, 41, 498–502. [Google Scholar] [CrossRef]
- García-García, M.L.; Calvo, C.; Casas, I.; Bracamonte, T.; Rellán, A.; Gozalo, F.; Tenorio, T.; Pérez-Breña, P. Human metapneumovirus bronchiolitis in infancy is an important risk factor for asthma at age 5. Pediatr. Pulmonol. 2007, 42, 458–464. [Google Scholar] [CrossRef]
- Kahn, J.S. Epidemiology of Human Metapneumovirus. Clin. Microbiol. Rev. 2006, 19, 546–557. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.; Mohakud, N.K.; Pena, L.; Kumar, S. Human metapneumovirus: Review of an important respiratory pathogen. Int. J. Infect. Dis. 2014, 25, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Diab, M.; Glasner, A.; Isaacson, B.; Bar-On, Y.; Drori, Y.; Yamin, R.; Duev-Cohen, A.; Danziger, O.; Zamostiano, R.; Mandelboim, M.; et al. NK-cell receptors NKp46 and NCR1 control human metapneumovirus infection. Eur. J. Immunol. 2017, 47, 692–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero-Plata, A.; Kolli, D.; Hong, C.; Casola, A.; Garofalo, R.P. Subversion of pulmonary dendritic cell function by paramyxovirus infections. J. Immunol. 2009, 182, 3072–3083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Qin, T.; Zhao, X.; Dong, S.; Zhu, J.; Peng, D.; Zhong, J.; Li, T.; Chen, X. Skewed balance of regulatory T cell and inflammatory T cell in IL-17 defect with human metapneumovirus infection. Cell. Immunol. 2018, 331, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Plata, A.; Casola, A.; Suárez, G.; Yu, X.; Spetch, L.; Peeples, M.E.; Garofalo, R.P. Differential Response of Dendritic Cells to Human Metapneumovirus and Respiratory Syncytial Virus. Am. J. Respir. Cell Mol. Boil. 2006, 34, 320–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolli, D.; Gupta, M.R.; Sbrana, E.; Velayutham, T.S.; Chao, H.; Casola, A.; Garofalo, R.P. Alveolar Macrophages Contribute to the Pathogenesis of Human Metapneumovirus Infection while Protecting against Respiratory Syncytial Virus Infection. Am. J. Respir. Cell Mol. Boil. 2014, 51, 502–515. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, R.; Harrod, K.; Shieh, W.-J.; Zaki, S.; Tripp, R.A. Human Metapneumovirus Persists in BALB/c Mice despite the Presence of Neutralizing Antibodies. J. Virol. 2004, 78, 14003–14011. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, R.; Tripp, R.A. The Immune Response to Human Metapneumovirus Is Associated with Aberrant Immunity and Impaired Virus Clearance in BALB/c Mice. J. Virol. 2005, 79, 5971–5978. [Google Scholar] [CrossRef] [Green Version]
- Herd, K.A.; Mahalingam, S.; Mackay, I.M.; Nissen, M.; Sloots, T.P.; Tindle, R.W. Cytotoxic T-Lymphocyte Epitope Vaccination Protects against Human Metapneumovirus Infection and Disease in Mice. J. Virol. 2006, 80, 2034–2044. [Google Scholar] [CrossRef] [Green Version]
- Kolli, D.; Bataki, E.L.; Spetch, L.; Guerrero-Plata, A.; Jewell, A.M.; Piedra, P.A.; Milligan, G.N.; Garofalo, R.P.; Casola, A. T Lymphocytes Contribute to Antiviral Immunity and Pathogenesis in Experimental Human Metapneumovirus Infection. J. Virol. 2008, 82, 8560–8569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herd, K.A.; Nelson, M.; Mahalingam, S.; Tindle, R.W. Pulmonary infection of mice with human metapneumovirus induces local cytotoxic T-cell and immunoregulatory cytokine responses like those seen with human respiratory syncytial virus. J. Gen. Virol. 2010, 91, 1302–1310. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.C.; Tollefson, S.J.; Johnson, M.; Gilchuk, P.; Boyd, K.L.; Shepherd, B.; Joyce, S.; Williams, J.V. Acute Clearance of Human Metapneumovirus Occurs Independently of Natural Killer Cells. J. Virol. 2014, 88, 10963–10969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diab, M.; Vitenshtein, A.; Drori, Y.; Yamin, R.; Danziger, O.; Zamostiano, R.; Mandelboim, M.; Bacharach, E.; Mandelboim, O. Suppression of human metapneumovirus (HMPV) infection by the innate sensing gene CEACAM1. Oncotarget 2016, 7, 66468–66479. [Google Scholar] [CrossRef] [Green Version]
- Sabo, Y.; Ehrlich, M.; Bacharach, E. The Conserved YAGL Motif in Human Metapneumovirus Is Required for Higher-Order Cellular Assemblies of the Matrix Protein and for Virion Production. J. Virol. 2011, 85, 6594–6609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berhani, O.; Nachmani, D.; Yamin, R.; Schmiedel, D.; Bar-On, Y.; Mandelboim, O. Vigilin Regulates the Expression of the Stress-Induced Ligand MICB by Interacting with Its 5? Untranslated Region. J. Immunol. 2017, 198, 3662–3670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glasner, A.; Oiknine-Djian, E.; Weisblum, Y.; Diab, M.; Panet, A.; Wolf, D.G.; Mandelboim, O. Zika Virus Escapes NK Cell Detection by Upregulating Major Histocompatibility Complex Class I Molecules. J. Virol. 2017, 91, e00785-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhrberg, M. The CD107 mobilization assay: Viable isolation and immunotherapeutic potential of tumor-cytolytic NK cells. Leukemia 2005, 19, 707–709. [Google Scholar] [CrossRef]
- Freymuth, F.; Vabret, A.; Legrand, L.; Eterradossi, N.; Lafay-Delaire, F.; Brouard, J.; Guillois, B. Presence of the new human metapneumovirus in French children with bronchiolitis. Pediatr. Infect. Dis. J. 2003, 22, 92–94. [Google Scholar] [CrossRef]
- Wolf, D.G.; Zakay-Rones, Z.; Fadeela, A.; Greenberg, D.; Dagan, R. High Seroprevalence of Human Metapneumovirus among Young Children in Israel. J. Infect. Dis. 2003, 188, 1865–1867. [Google Scholar] [CrossRef]
- Weekes, M.P.; Tomasec, P.; Huttlin, E.L.; Fielding, C.A.; Nusinow, D.; Stanton, R.J.; Wang, I.C.Y.; Aicheler, R.; Murrell, I.; Wilkinson, G.W.; et al. Quantitative Temporal Viromics: An Approach to Investigate Host-Pathogen Interaction. Cell 2014, 157, 1460–1472. [Google Scholar] [CrossRef] [Green Version]
- Stern-Ginossar, N.; Mandelboim, O. An integrated view of the regulation of NKG2D ligands. Immunology 2009, 128, 1–6. [Google Scholar] [CrossRef]
- Fielding, C.A.; Aicheler, R.; Stanton, R.J.; Wang, E.C.; Han, S.; Seirafian, S.; Davies, J.; McSharry, B.; Weekes, M.P.; Antrobus, P.R.; et al. Two Novel Human Cytomegalovirus NK Cell Evasion Functions Target MICA for Lysosomal Degradation. PLoS Pathog. 2014, 10, e1004058. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diab, M.; Schmiedel, D.; Seidel, E.; Bacharach, E.; Mandelboim, O. Human Metapneumovirus Escapes NK Cell Recognition through the Downregulation of Stress-Induced Ligands for NKG2D. Viruses 2020, 12, 781. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070781
Diab M, Schmiedel D, Seidel E, Bacharach E, Mandelboim O. Human Metapneumovirus Escapes NK Cell Recognition through the Downregulation of Stress-Induced Ligands for NKG2D. Viruses. 2020; 12(7):781. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070781
Chicago/Turabian StyleDiab, Mohammad, Dominik Schmiedel, Einat Seidel, Eran Bacharach, and Ofer Mandelboim. 2020. "Human Metapneumovirus Escapes NK Cell Recognition through the Downregulation of Stress-Induced Ligands for NKG2D" Viruses 12, no. 7: 781. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070781