Characterization of PlyB221 and PlyP32, Two Novel Endolysins Encoded by Phages Preying on the Bacillus cereus Group


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Bioinformatic Analysis
2.3. Cloning Experiments
2.4. Protein Expression and Purification
2.5. Spot-on-Plate Assay
2.6. Endolysin Activity Spectrum
2.7. Endolysin Activity on Purified Cell Wall
2.8. Endolysin Biochemical Characterization
2.9. Cell Wall Decoration Assay
3. Results
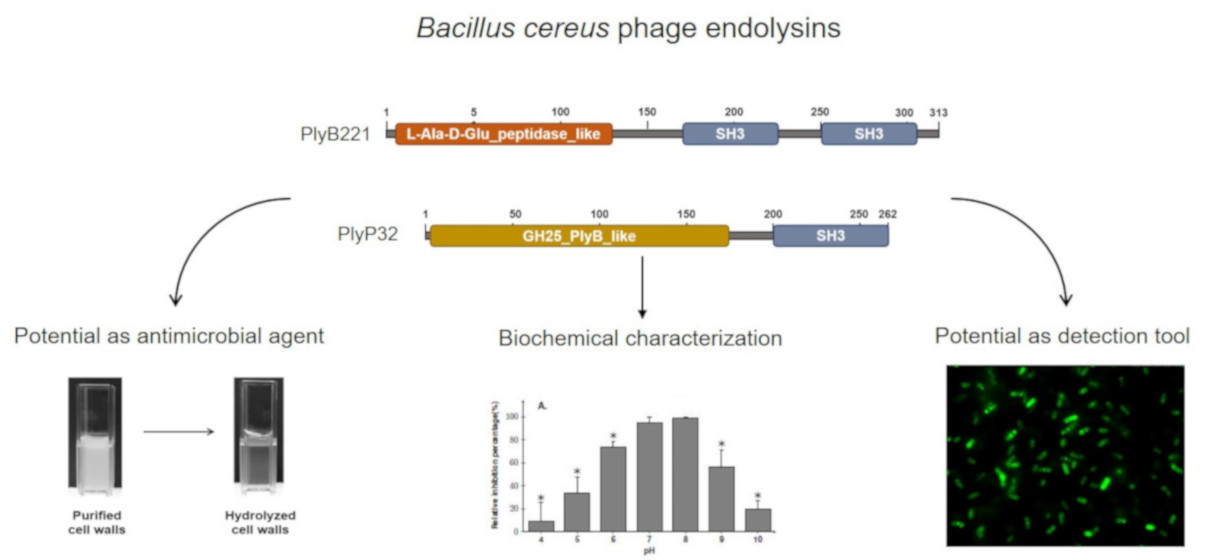
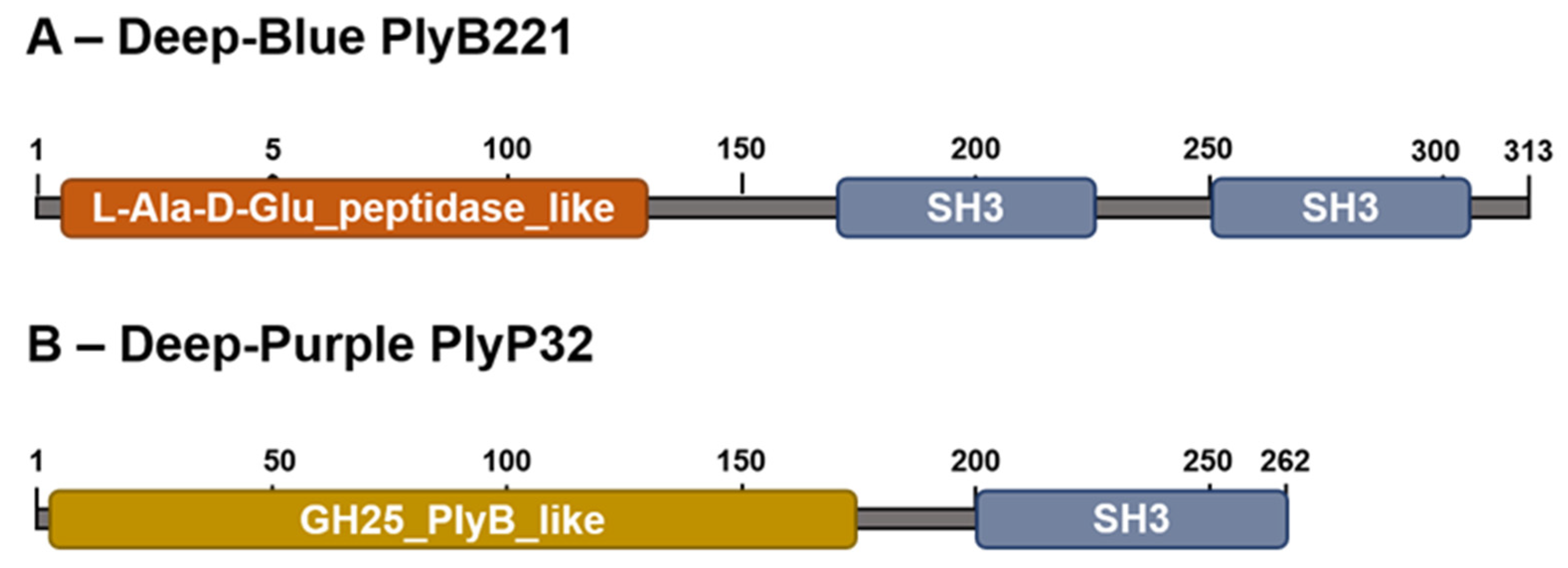
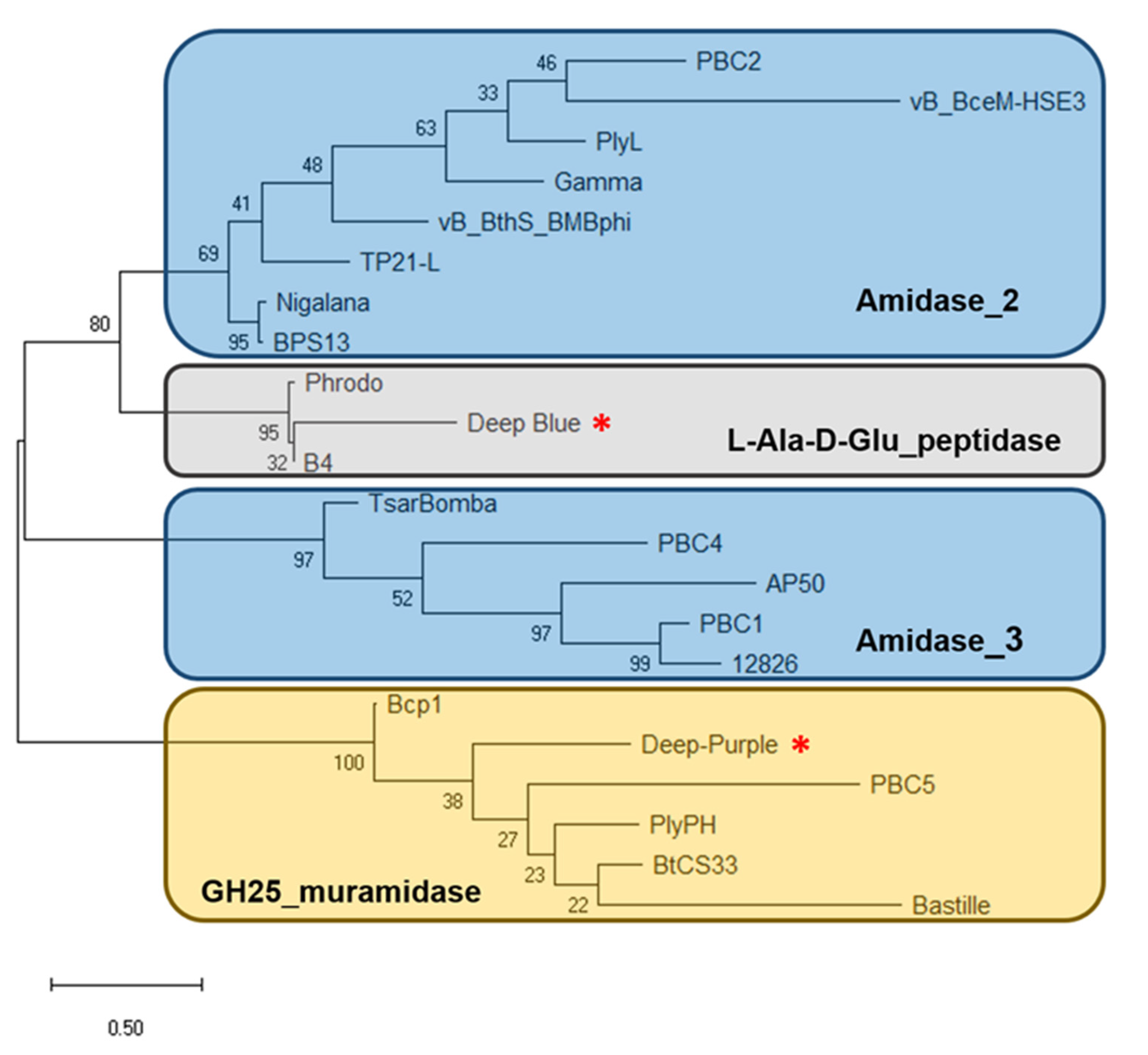
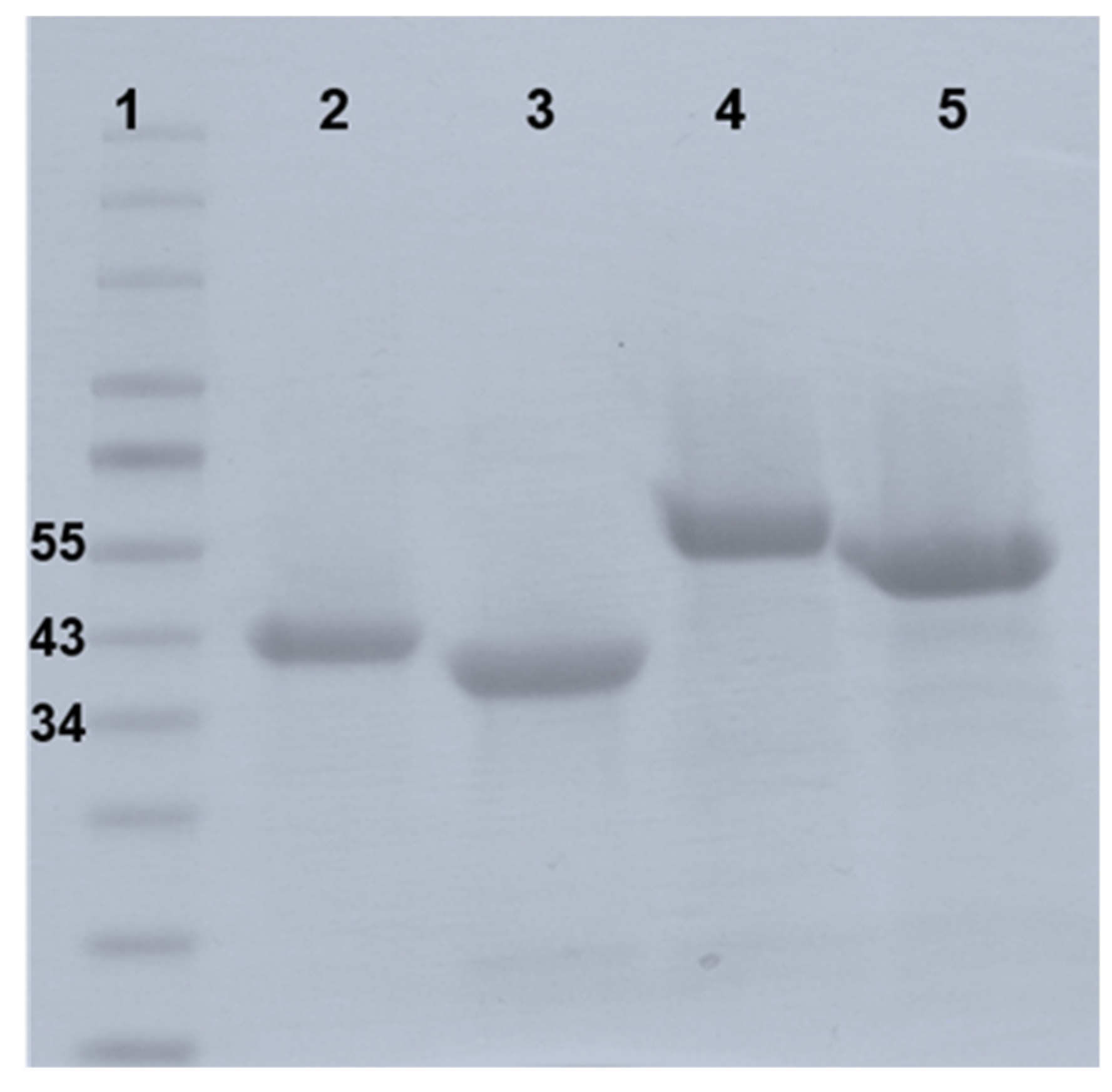
3.1. PlyB221 and PlyP32 Display the Canonical Features Of Endolysins
3.2. PlyB221 and PlyP32 Show a Bacteriolytic Spectrum Broader than That of Their Parental Phages
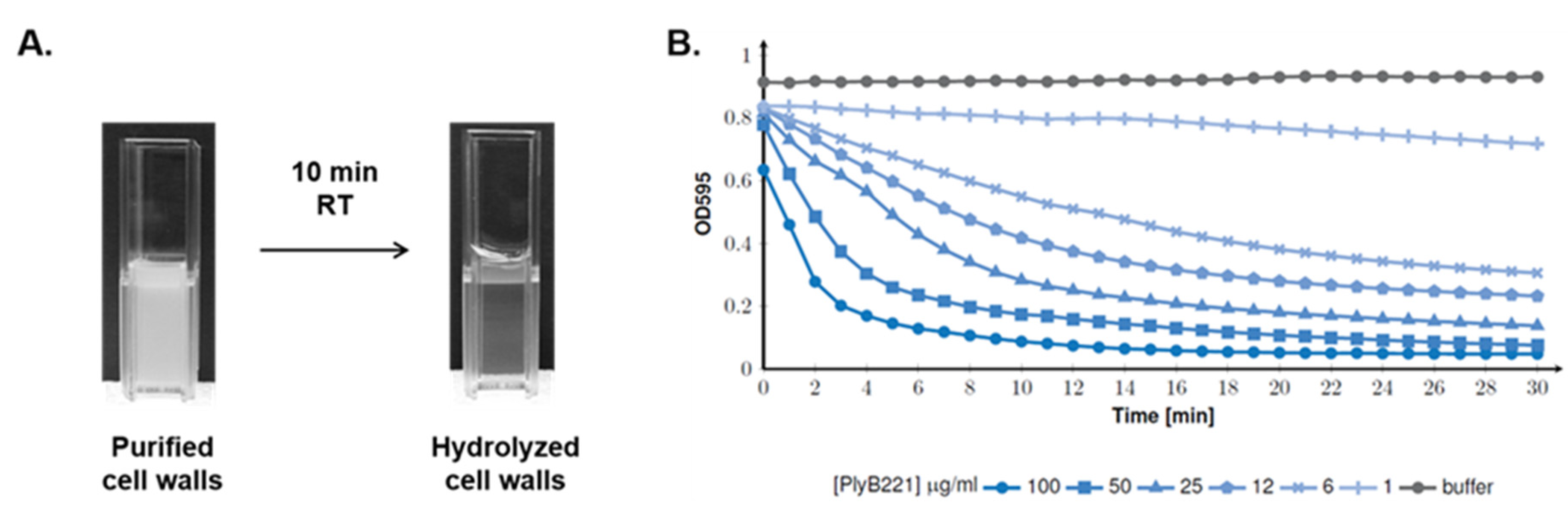
3.3. PlyB221 and PlyP32 Display a Peptidoglycan Hydrolyzing Activity
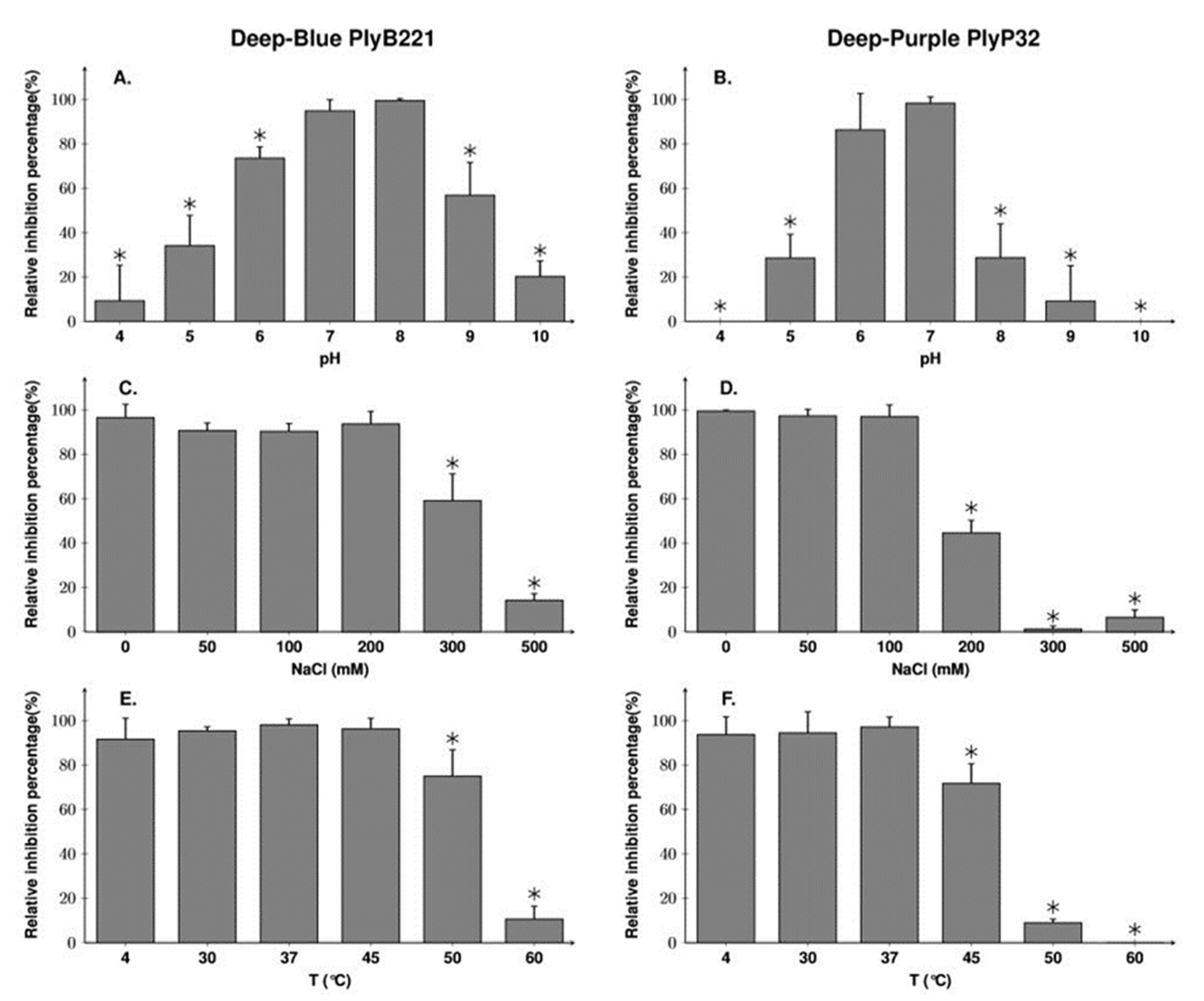
3.4. PlyB221 Displays a Somewhat more Stable Activity than PlyP32
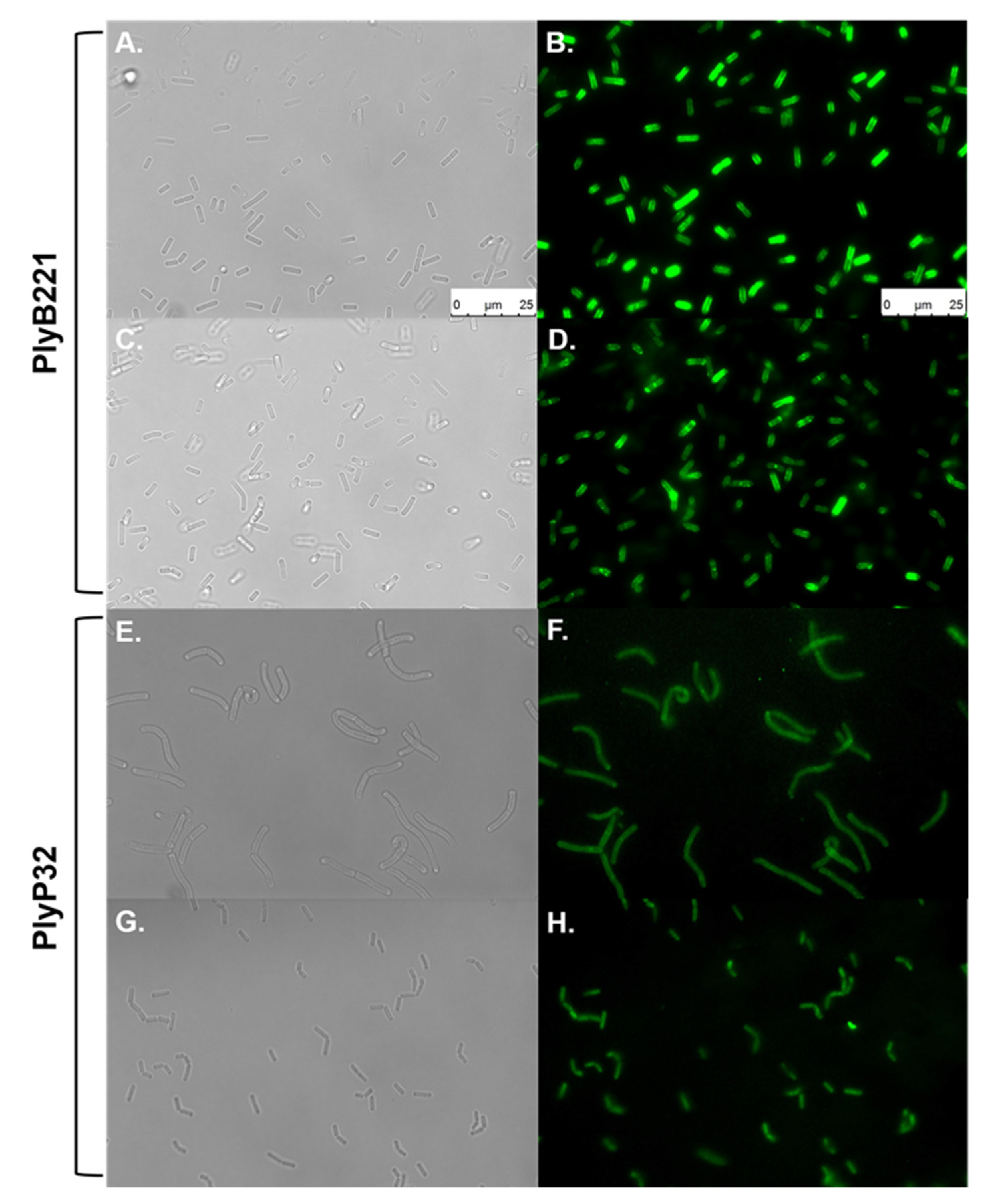
3.5. PlyB221 and PlyP32 Cell Wall Binding Domain (CBD) Can Decorate Bacillus Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Borysowski, J.; Miedzybrodzki, R.; Gorski, A. Phage Therapy: Current Research and Applications; Caister Academic Press: Poole, UK, 2014; ISBN 978-1-908230-40-9. [Google Scholar]
- Díaz-Muñoz, S.L.; Koskella, B. Bacteria-Phage interactions in natural environments. In Advances in Applied Microbiology; Academic Press: Cambridge, MA, USA, 2014; Volume 89, pp. 135–183. ISBN 9780128002599. [Google Scholar]
- Gillis, A.; Mahillon, J. Phages preying on Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis: Past, present and future. Viruses 2014, 6, 2623–2672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, S.; São-José, C. Enzymes and mechanisms employed by tailed bacteriophages to breach the bacterial cell barriers. Viruses 2018, 10, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broendum, S.S.; Buckle, A.M.; McGowan, S. Catalytic diversity and cell wall binding repeats in the phage-encoded endolysins. Mol. Microbiol. 2018, 110, 879–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahill, J.; Young, R. Phage Lysis: Multiple Genes for Multiple Barriers. In Advances in Virus Research; Academic Press Inc.: Cambridge, MA, USA, 2019; Volume 103, pp. 33–70. ISBN 9780128177228. [Google Scholar]
- Oliveira, H.; Melo, L.D.R.; Santos, S.B.; Nóbrega, F.L.; Ferreira, E.C.; Cerca, N.; Azeredo, J.; Kluskens, L.D. Molecular aspects and comparative genomics of bacteriophage endolysins. J. Virol. 2013, 87, 4558–4570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamilla, S.; Jain, V. Mycobacteriophage D29 holin C-terminal region functionally assists in holin aggregation and bacterial cell death. FEBS J. 2016, 283, 173–190. [Google Scholar] [CrossRef]
- Nelson, D.C.; Schmelcher, M.; Rodriguez-Rubio, L.; Klumpp, J.; Pritchard, D.G.; Dong, S.; Donovan, D.M. Endolysins as antimicrobials. Adv. Virus Res. 2012, 83, 299–365. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Kim, Y.; Ryu, S.; Lee, J.; Lee, J. Biocontrol and rapid detection of food-borne pathogens using bacteriophages and endolysins. Front. Microbiol. 2016, 7, 1–15. [Google Scholar] [CrossRef]
- Kong, M.; Na, H.; Ha, N.-C.; Ryu, S. Molecular characterization of LysPBC2, a novel endolysin harboring a Bacillus cereus spore binding domain. Appl. Environ. Microbiol. 2018, 85, e02462-18. [Google Scholar] [CrossRef] [Green Version]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [Green Version]
- Kretzer, J.W.; Schmelcher, M.; Loessner, M.J. Ultrasensitive and fast diagnostics of viable Listeria cells by CBD magnetic separation combined with A511::luxAB detection. Viruses 2018, 10, 626. [Google Scholar] [CrossRef] [Green Version]
- Gerstmans, H.; Criel, B.; Briers, Y. Synthetic biology of modular endolysins. Biotechnol. Adv. 2018, 36, 624–640. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.Y.; Kim, J.S.; Paek, W.K.; Lim, J.; Lee, H.; Kim, P.I.; Ma, J.Y.; Kim, W.; Chang, Y.H. Bacillus manliponensis sp. nov., a new member of the Bacillus cereus group isolated from foreshore tidal flat sediment. J. Microbiol. 2011, 49, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.A.; Beno, S.M.; Kent, D.J.; Carroll, L.M.; Martin, N.H.; Boor, K.J.; Kovac, J. Bacillus wiedmannii sp. nov., a psychrotolerant and cytotoxic Bacillus cereus group species isolated from dairy foods and dairy environments. Int. J. Syst. Evol. Microbiol. 2016, 66, 4744–4753. [Google Scholar] [CrossRef]
- Park, J.; Yun, J.; Lim, J.A.; Kang, D.H.; Ryu, S. Characterization of an endolysin, LysBPS13, from a Bacillus cereus bacteriophage. FEMS Microbiol. Lett. 2012, 332, 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, B.; Yun, J.; Lim, J.-A.; Shin, H.; Heu, S.; Ryu, S. Characterization of LysB4, an endolysin from the Bacillus cereus-infecting bacteriophage B4. BMC Microbiol. 2012, 12, 33. [Google Scholar] [CrossRef] [Green Version]
- Kong, M.; Ryu, S. Bacteriophage PBC1 and its endolysin as an antimicrobial agent against Bacillus cereus. Appl. Environ. Microbiol. 2015, 81, 2274–2283. [Google Scholar] [CrossRef] [Green Version]
- Etobayeva, I.; Linden, S.B.; Alem, F.; Harb, L.; Rizkalla, L.; Mosier, P.D.; Johnson, A.A.; Temple, L.; Hakami, R.M.; Nelson, D.C. Discovery and biochemical characterization of PlyP56, PlyN74, and PlyTB40—Bacillus specific endolysins. Viruses 2018, 10, 276. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Peng, Q.; Yang, S.; Zhang, S.; Fu, Y.; Wu, Y.; Gao, M. Isolation of a novel Bacillus thuringiensis phage representing a new phage lineage and characterization of its endolysin. Viruses 2018, 10, 611. [Google Scholar] [CrossRef] [Green Version]
- Hock, L.; Gillis, A.; Mahillon, J. Complete genome sequence of bacteriophage Deep-Blue infecting emetic Bacillus cereus. Genome Announc. 2016, 4, e00115-16. [Google Scholar] [CrossRef] [Green Version]
- Hock, L.; Leprince, A.; Tournay, M.; Gillis, A.; Mahillon, J. Biocontrol potential of phage Deep-Blue against psychrotolerant Bacillus weihenstephanensis. Food Control 2019, 102, 94–103. [Google Scholar] [CrossRef]
- Hock, L.; Gillis, A.; Mahillon, J. Complete genome sequence of bacteriophage Deep-Purple, a novel member of the family Siphoviridae infecting Bacillus cereus. Arch. Virol. 2018, 163, 2555–2559. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hunter, S.; Apweiler, R.; Attwood, T.K.; Bairoch, A.; Bateman, A.; Binns, D.; Bork, P.; Das, U.; Daugherty, L.; Duquenne, L.; et al. InterPro: The integrative protein signature database. Nucleic Acids Res. 2009, 37, 211–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Söding, J.; Biegert, A.; Lupas, A.N. The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acids Res. 2005, 33, 244–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 37, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.M.; Kruger, N.J. The Bradford method for protein quantitation. Basic Protein Pept. Protoc. 2003, 32, 9–16. [Google Scholar] [CrossRef]
- DeWitt, T.; Grossman, A.D. The bifunctional cell wall hydrolase CwlT is needed for conjugation of the integrative and conjugative element ICEBs1 in Bacillus subtilis and B. anthracis. J. Bacteriol. 2014, 196, 1588–1596. [Google Scholar] [CrossRef] [Green Version]
- Bielmann, R.; Habann, M.; Eugster, M.R.; Lurz, R.; Calendar, R.; Klumpp, J.; Loessner, M.J. Receptor binding proteins of Listeria monocytogenes bacteriophages A118 and P35 recognize serovar-specific teichoic acids. Virology 2015, 477, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.O.; Kong, M.; Kim, I.; Bai, J.; Cha, S.; Kim, B.; Ryu, K.-S.; Ryu, S.; Suh, J.-Y. Structural basis for cell-wall recognition by bacteriophage PBC5 endolysin. Structure 2019, 27, 1355–1365. [Google Scholar] [CrossRef]
- Benešík, M.; Nováček, J.; Janda, L.; Dopitová, R.; Pernisová, M.; Melková, K.; Tišáková, L.; Doškař, J.; Žídek, L.; Hejátko, J.; et al. Role of SH3b binding domain in a natural deletion mutant of Kayvirus endolysin LysF1 with a broad range of lytic activity. Virus Genes 2018, 54, 130–139. [Google Scholar] [CrossRef]
- Yuan, Y.; Peng, Q.; Gao, M. Characteristics of a broad lytic spectrum endolysin from phage BtCS33 of Bacillus thuringiensis. BMC Microbiol. 2012, 12, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischetti, V.A. Development of phage lysins as novel therapeutics: A historical perspective. Viruses 2018, 10, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Auwera, G.A.; Feldgarden, M.; Kolter, R.; Mahillon, J.; Diseases, I.; Services, H. Whole-Genome Sequences of 94 Environmental Isolates of Bacillus cereus sensu lato. Genome Announc. 2013, 1, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castiaux, V.; Liu, X.; Delbrassinne, L.; Mahillon, J. Is Cytotoxin K from Bacillus cereus a bona fide enterotoxin? Int. J. Food Microbiol. 2015, 211, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, P.C.B.; Kramer, J.M.; Jörgensen, K.; Gilbert, R.J.; Melling, J. Properties and production characteristics of vomiting, diarrheal, and necrotizing toxins of Bacillus cereus. Am. J. Clin. Nutr. 1979, 32, 219–228. [Google Scholar] [CrossRef]
- Hoffmaster, A.R.; Novak, R.T.; Marston, C.K.; Gee, J.E.; Helsel, L.; Pruckler, J.M.; Wilkins, P.P. Genetic diversity of clinical isolates of Bacillus cereus using multilocus sequence typing. BMC Microbiol. 2008, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lechner, S.; Mayr, R.; Francis, K.P.; Prüß, B.M.; Kaplan, T.; Wießner-Gunkel, E.; Stewart, G.S.A.B.; Scherer, S. Bacillus weihenstephanensis sp. nov. is a new psychrotolerant species of the Bacillus cereus group. Int. J. Syst. Evol. Microbiol. 1998, 48, 1373–1382. [Google Scholar] [CrossRef]
- Jensen, G.B.; Wilcks, A.; Petersen, S.S.; Damgaard, J.; Baum, J.A.; Andrup, L. The genetic basis of the aggregation system in Bacillus thuringiensis subsp. israelensis is located on the large conjugative plasmid pXO16. J. Bacteriol. 1995, 177, 2914–2917. [Google Scholar] [CrossRef] [Green Version]
- Guinebretière, M.H.; Auger, S.; Galleron, N.; Contzen, M.; de Sarrau, B.; de Buyser, M.L.; Lamberet, G.; Fagerlund, A.; Granum, P.E.; Lereclus, D.; et al. Bacillus cytotoxicus sp. nov. is a novel thermotolerant species of the Bacillus cereus group occasionally associated with food poisoning. Int. J. Syst. Evol. Microbiol. 2013, 63, 31–40. [Google Scholar] [CrossRef]
- Kunst, F.; Lenoble, P.; Meurice, G.; Sekowska, A.; Vallenet, D.; Danchin, A.; Wang, T.; Moszer, I.; Me, C. From a consortium sequence to a unified sequence: The Bacillus subtilis 168 reference genome a decade later. Microbiology 2009, 155, 1758–1775. [Google Scholar] [CrossRef] [Green Version]
- Caulier, S.; Gillis, A.; Colau, G.; Licciardi, F.; Liépin, M.; Desoignies, N.; Modrie, P.; Legrève, A.; Mahillon, J.; Bragard, C. Versatile antagonistic activities of soil-borne Bacillus spp. and Pseudomonas spp. against Phytophthora infestans and other potato pathogens. Front. Microbiol. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Love, M.J.; Bhandari, D.; Dobson, R.C.J.; Billington, C. Potential for bacteriophage endolysins to supplement or replace antibiotics in food production and clinical care. Antibiotics 2018, 7, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuch, R.; Pelzek, A.J.; Nelson, D.C.; Fischetti, V.A. The PlyB endolysin of bacteriophage vB_BanS_Bcp1 exhibits broad-spectrum bactericidal activity against Bacillus cereus sensu lato isolates. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Gao, M.; Wu, D.; Liu, P.; Wu, Y. Genome characteristics of a novel phage from Bacillus thuringiensis showing high similarity with phage from Bacillus cereus. PLoS ONE 2012, 7, e37557. [Google Scholar] [CrossRef] [Green Version]
- Yoong, P.; Schuch, R.; Nelson, D.; Fischetti, V.A. PlyPH, a bacteriolytic enzyme with a broad pH range of activity and lytic action against Bacillus anthracis. J. Bacteriol. 2006, 188, 2711–2714. [Google Scholar] [CrossRef] [Green Version]
- Schmelcher, M.; Waldherr, F.; Loessner, M.J. Listeria bacteriophage peptidoglycan hydrolases feature high thermoresistance and reveal increased activity after divalent metal cation substitution. Appl. Microbiol. Biotechnol. 2012, 93, 633–643. [Google Scholar] [CrossRef] [Green Version]
- Peng, Q.; Yuan, Y. Characterization of a novel phage infecting the pathogenic multidrug-resistant Bacillus cereus and functional analysis of its endolysin. Appl. Microbiol. Biotechnol. 2018, 102, 7901–7912. [Google Scholar] [CrossRef]
- Park, S.; Jun, S.Y.; Kim, C.-H.; Jung, G.M.; Son, J.S.; Jeong, S.T.; Yoon, S.J.; Lee, S.Y.; Kang, S.H. Characterisation of the antibacterial properties of the recombinant phage endolysins AP50-31 and LysB4 as potent bactericidal agents against Bacillus anthracis. Sci. Rep. 2018, 8, 18. [Google Scholar] [CrossRef] [Green Version]
- Gründling, A.; Schneewind, O. Cross-linked peptidoglycan mediates lysostaphin binding to the cell wall envelope of Staphylococcus aureus. J. Bacteriol. 2006, 188, 2463–2472. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains/Plasmids | Species/Purpose | Reference |
|---|---|---|
| Strains | ||
| 10-beta | E. coli/cloning strain | NEB |
| BL21(DE3) | E. coli/expression strain | Novagen |
| Plasmids | ||
| pET30a | Expression plasmid | Novagen |
| pUC18::gfp | GFP amplification | Clontech/Takara |
| pET30::gp221 | Test PlyB221 lytic activity | This study |
| pET30::gfp(linker)::gp221_cbd | Test PlyB221 CBD binding activity | This study |
| pET30::gp32 | Test PlyP32 lytic activity | This study |
| pET30::gfp(linker)::gp32_cdb | Test PlyP32 CBD binding activity | This study |
| Target | Primer Name | Sequence (5′ → 3′) |
|---|---|---|
| gfp | GFP_EcoRI_F | TTCCGAATTCAAAGGAGAAGAACTTTTCACTGGAG |
| GFP_EagI_linker_R | TTCGGCCGTCCACTACCTGATCCACTACCTTTGTAGAGCTCATCCATGCC | |
| gp221 | gp221_NcoI_F | TATCCATGGTGGCAATGTCTTTAGATACTTTAATCA |
| gp221_XhoI_R | TATTCTCGAGCTACTCCTCAATGAAGTTGATGTATG | |
| gp221_cbd | gp221_CBD_EagI_F | AGCGGCCGGGTGGAGCAGTAGACACT |
| gp221_XhoI_R | TATTCTCGAGCTACTCCTCAATGAAGTTGATGTATG | |
| gp32 | gp32_NcoI_F | TATCCATGGACAAAAAGATTTTAGATATTTCACATCAC |
| gp32_XhoI_R | ATTACTCGAGTTAAATGATTTTTACGTTATCGCCA | |
| gp32_cbd | gp32[181]_EagI_F | AACGGCCGGTTAGTTTCTATATTGGTAACTCTT |
| gp32_XhoI_R | ATTACTCGAGTTAAATGATTTTTACGTTATCGCCA |
| Deep-Blue | Deep-Purple | |||||||
|---|---|---|---|---|---|---|---|---|
| Strain | Species | Reference | Φ a | PlyB221 b | CBD c | Φ | PlyP32 | CBD |
| Strains of the B. cereus group | ||||||||
| VD021 | B. cereus | [37] | S | 74.9 ± 3.9 | + | S | 77.1 ± 2.4 | + |
| ATCC 10987 | B. cereus | [38] | I | 72.6 ± 2.3 | + | I | 77.8 ± 9.9 | + |
| F4810/72 | B. cereus * | [39] | L | 67.8 ± 10.7 | + | L | 70.3 ± 1.7 | + |
| H3081.97 | B. cereus * | [40] | L | 57.3 ± 5.9 | + | I | 32.7 ± 0.7 | + |
| WSBC10202 | B. weihenstephanensis | [41] | S | 61.1 ± 3.4 | + | L | 71.2 ± 8.4 | + |
| WSBC10204 | B. weihenstephanensis | [41] | L | 76.4 ± 9.1 | + | L | 68 ± 6.8 | + |
| Si0239 | B. weihenstephanensis * | [23] | S | 66.6 ± 4 | + | L | 79.3 ± 2.8 | + |
| LH002 | B. weihenstephanensis * | [24] | I | 69.3 ± 10.5 | + | S | 75.7 ± 5.7 | + |
| GBJ001 | B. thuringiensis | [42] | L | 76.2 ± 3 | + | L | 85 ± 0.4 | + |
| HD4 | B. thuringiensis | BGSC 1 | L | 38.1 ± 0.9 | + | S | 73.4 ± 7 | + |
| NVH 391-98 | B. cytotoxicus | [43] | L | 74.2 ± 6 | + | L | 80.4 ± 4.7 | + |
| BcA20 086 | B. cytotoxicus | MIAE 2 | L | 46.6 ± 11 | + | L | 34.4 ± 14.2 | + |
| KBS 2–12 | B. mycoides | MIAE | L | 62.1 ± 2.7 | + | S | 62 ± 4.2 | + |
| KNC2-18 | B. mycoides | MIAE | I | 66 ± 23.3 | + | I | 15.4 ± 8.6 | + |
| Other Gram-positive bacteria | ||||||||
| Bs168 | B. subtilis | [44] | I | 0.2 ± 0.4 | - | I | 0 ± 0 | - |
| ATCC 10716 | B. licheniformis | BBCM 3 | I | 23.2 ± 1 | - | I | 40.1 ± 4.5 | - |
| SI0279 | B. pumilus | MIAE | I | 6.8 ± 0.9 | - | I | 21.8 ± 7.7 | - |
| 10A-B5 | B. pumilus | [45] | I | 28.3 ± 1.5 | - | I | 26.3 ± 10.2 | - |
| SI0003 | B. megaterium | MIAE | L | 73.9 ± 2.7 | + | I | 34.2 ± 6.6 | + |
| 10A-B3 | B. megaterium | [45] | I | 79.9 ± 0.5 | + | I | 35.3 ± 14.3 | + |
| Liv001 | L. ivanovii | MBLG 4 | I | 0.5 ± 0.9 | - | I | 0 ± 0 | - |
| ATCC 12228 | S. epidermidis | BBCM | I | 0.1 ± 0.2 | - | I | 0 ± 0 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leprince, A.; Nuytten, M.; Gillis, A.; Mahillon, J. Characterization of PlyB221 and PlyP32, Two Novel Endolysins Encoded by Phages Preying on the Bacillus cereus Group. Viruses 2020, 12, 1052. https://0-doi-org.brum.beds.ac.uk/10.3390/v12091052
Leprince A, Nuytten M, Gillis A, Mahillon J. Characterization of PlyB221 and PlyP32, Two Novel Endolysins Encoded by Phages Preying on the Bacillus cereus Group. Viruses. 2020; 12(9):1052. https://0-doi-org.brum.beds.ac.uk/10.3390/v12091052
Chicago/Turabian StyleLeprince, Audrey, Manon Nuytten, Annika Gillis, and Jacques Mahillon. 2020. "Characterization of PlyB221 and PlyP32, Two Novel Endolysins Encoded by Phages Preying on the Bacillus cereus Group" Viruses 12, no. 9: 1052. https://0-doi-org.brum.beds.ac.uk/10.3390/v12091052