Experimental Infection and Genetic Characterization of Two Different Capripox Virus Isolates in Small Ruminants

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Infection and Collection of Samples
2.3. DNA Extraction of Different Samples and Pan Capripox Real-Time qPCR Analysis
2.4. Serological Analyses
2.5. Sequencing of SPPV-“V/104” Strain and GTPV-“V/103” Strain
3. Results
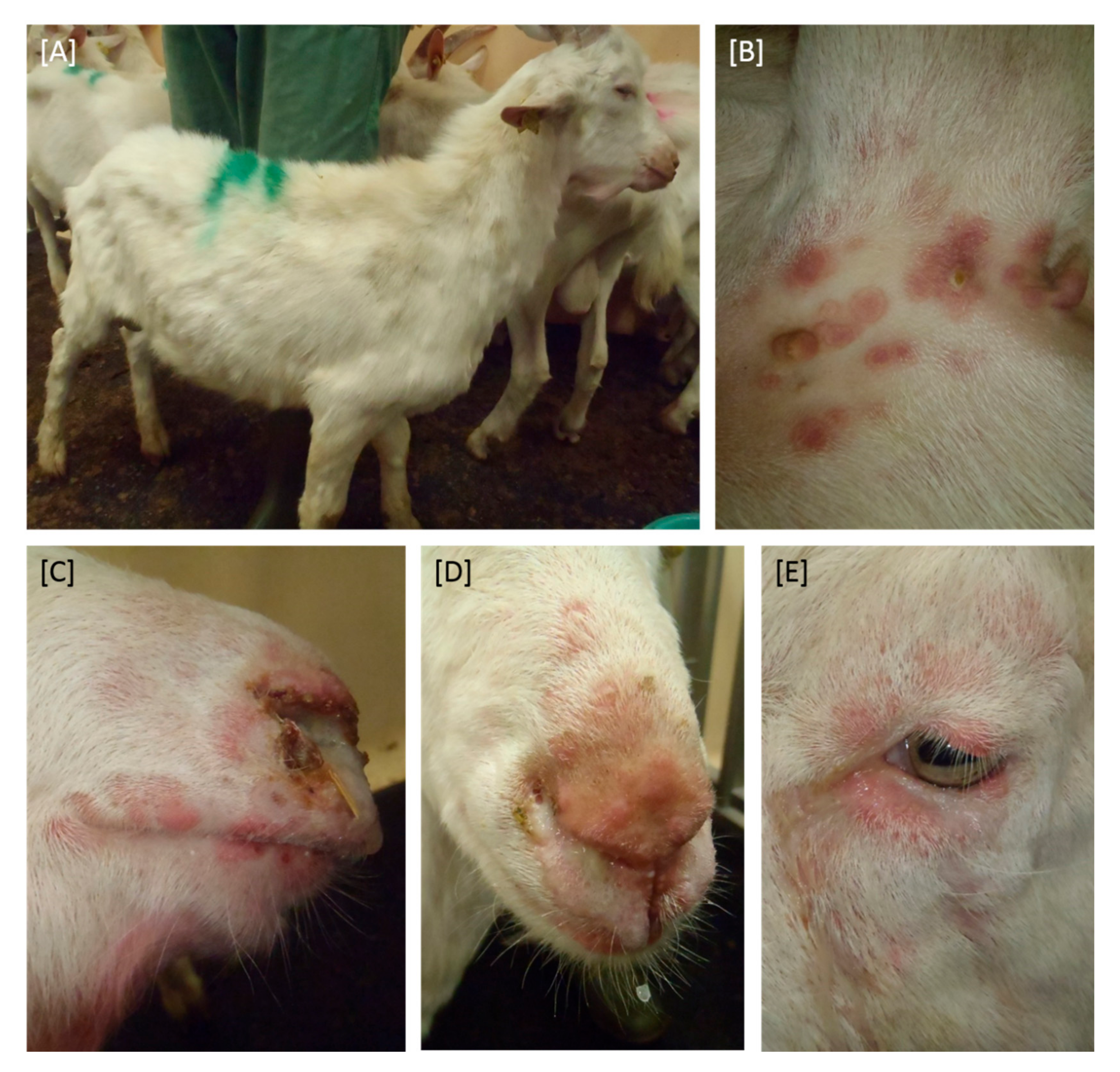
3.1. Clinical Signs after Experimental Infection
3.2. Replication and Virus Shedding
3.3. Viral Genome Loads in Organ Samples
3.4. Serological Evaluation
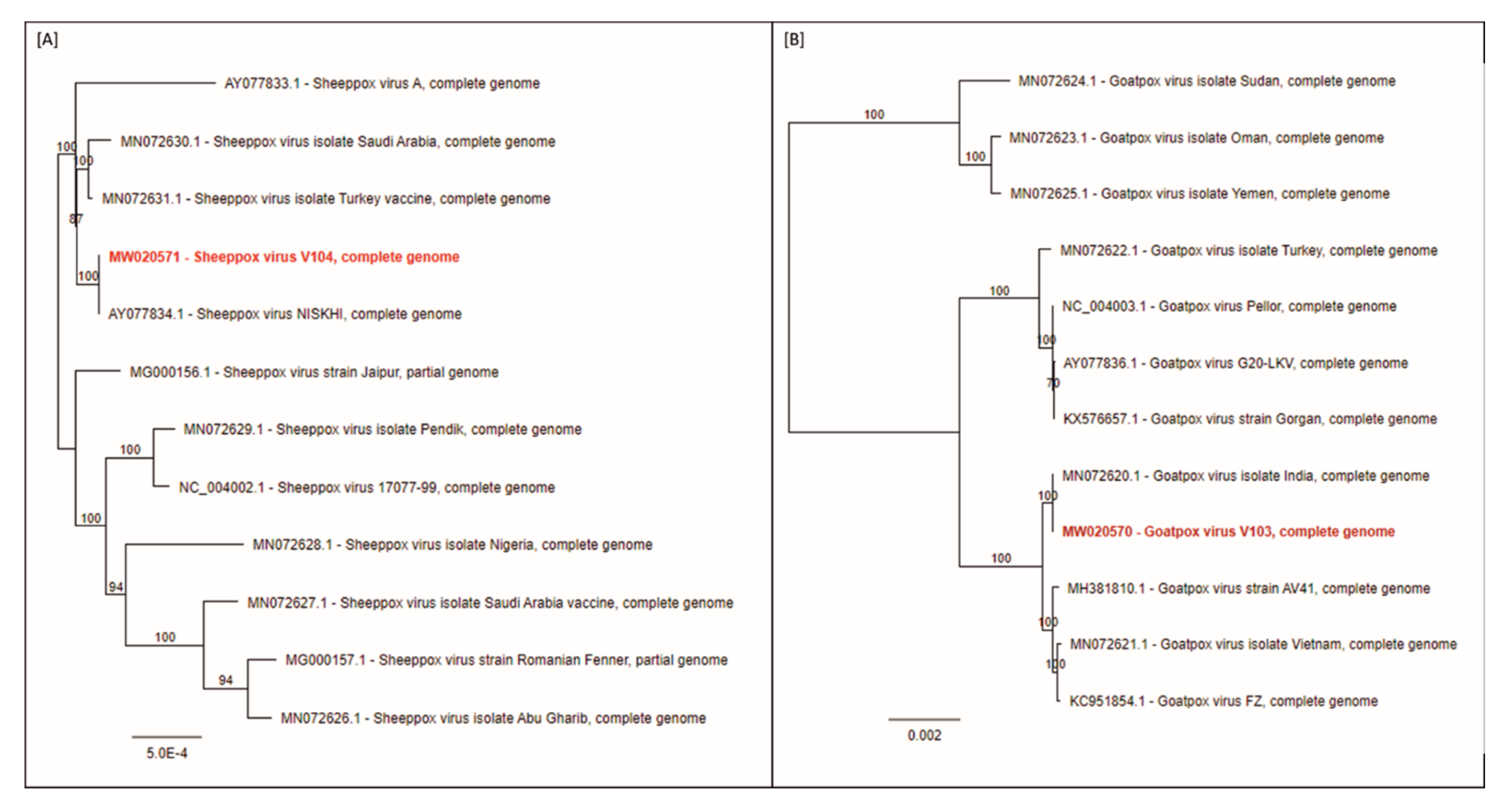
3.5. Full Genome Sequencing of Strains SPPV-“V/104” and GTPV-“V/103”
4. Discussion
4.1. Pathogenesis in Sheep after Inoculation with the SPPV-“V/104” Strain
4.2. Pathogenesis in Goats after Inoculation with the GTPV-“V/103” Strain
4.3. Comparison of the Different Inoculation Methods
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E. Virus Taxonomy-Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier Academic Press: Amsterdam, The Netherlands, 2012; pp. 291–309. [Google Scholar]
- OIE. OIE-Listed Diseases, Infections and Infestations in Force in 2020. 2020. Available online: https://www.oie.int/animal-health-in-the-world/oie-listed-diseases-2019/ (accessed on 15 July 2020).
- Kitching, P. Progress towards sheep and goat pox vaccines. Vaccine 1983, 1, 4–9. [Google Scholar] [CrossRef]
- Bhanuprakash, V.; Indrani, B.K.; Hosamani, M.; Singh, R.K. The current status of sheep pox disease. Comp. Immunol. Microbiol. Infect. Dis. 2006, 29, 27–60. [Google Scholar] [CrossRef]
- Rao, T.V.; Bandyopadhyay, S.K. A comprehensive review of goat pox and sheep pox and their diagnosis. Anim. Health Res. Rev. 2000, 1, 127–136. [Google Scholar] [CrossRef] [PubMed]
- OIE. Technical Disease Card: Sheep pox and Goat Pox. 2013. Available online: https://www.oie.int/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/Disease_cards/SHEEP_GOAT_POX.pdf (accessed on 28 September 2020).
- OIE. Sheep Pox and Goat Pox. OIE Terrestrial Manual 2017, Chapter 2.7.13. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/2.07.13_S_POX_G_POX.pdf (accessed on 28 September 2020).
- Babiuk, S.; Bowden, T.R.; Parkyn, G.; Dalman, B.; Hoa, D.M.; Long, N.T.; Vu, P.P.; Bieu do, X.; Copps, J.; Boyle, D.B. Yemen and Vietnam capripoxviruses demonstrate a distinct host preference for goats compared with sheep. J. Gen. Virol. 2009, 90, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Tuppurainen, E.S.M.; Venter, E.H.; Shisler, J.L.; Gari, G.; Mekonnen, G.A.; Juleff, N.; Lyons, N.A.; De Clercq, K.; Upton, C.; Bowden, T.R.; et al. Review: Capripoxvirus Diseases: Current Status and Opportunities for Control. Transbound. Emerg. Dis. 2017, 64, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Kitching, R.P.; Carn, V.M. Sheep Pox and goat pox. In Office International des Epizooties Manual of Diagnostic Tests and Vaccines for Terrestrial Animals (Mammals, Birds and Bees); OIE: Paris, France, 2004. [Google Scholar]
- Babiuk, S.; Bowden, T.R.; Boyle, D.B.; Wallace, D.B.; Kitching, R.P. Capripoxviruses: An emerging worldwide threat to sheep, goats and cattle. Transbound. Emerg. Dis. 2008, 55, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitching, R.P.; Taylor, W.P. Transmission of capripoxvirus. Res. Vet. Sci. 1985, 39, 196–199. [Google Scholar] [CrossRef]
- Carn, V.M. Control of capripoxvirus infections. Vaccine 1993, 11, 1275–1279. [Google Scholar] [CrossRef]
- Carn, V.M.; Kitching, R.P. An investigation of possible routes of transmission of lumpy skin disease virus (Neethling). Epidemiol. Infect. 1995, 114, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Mechanical transmission of lumpy skin disease virus by Aedes aegypti (Diptera: Culicidae). Epidemiol. Infect. 2001, 126, 317–321. [Google Scholar] [CrossRef] [Green Version]
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Attempted mechanical transmission of lumpy skin disease virus by biting insects. Med. Vet. Entomol. 2003, 17, 294–300. [Google Scholar] [CrossRef]
- Mellor, P.S.; Kitching, R.P.; Wilkinson, P.J. Mechanical transmission of capripox virus and African swine fever virus by Stomoxys calcitrans. Res. Vet. Sci. 1987, 43, 109–112. [Google Scholar] [CrossRef]
- Bowden, T.R.; Babiuk, S.L.; Parkyn, G.R.; Copps, J.S.; Boyle, D.B. Capripoxvirus tissue tropism and shedding: A quantitative study in experimentally infected sheep and goats. Virology 2008, 371, 380–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, F.G. Characteristics of a virus causing a pox disease in sheep and goats in Kenya, with observation on the epidemiology and control. J. Hyg. (London) 1976, 76, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.M.; Chu, Y.F.; Wu, G.H.; Zhao, Z.X.; Li, J.; Zhu, H.X.; Zhang, Q. An outbreak of sheep pox associated with goat poxvirus in Gansu province of China. Vet. Microbiol. 2012, 156, 425–428. [Google Scholar] [CrossRef] [PubMed]
- Kitching, R.P. Passive protection of sheep against capripoxvirus. Res. Vet. Sci. 1986, 41, 247–250. [Google Scholar] [CrossRef]
- Lamien, C.E.; Le Goff, C.; Silber, R.; Wallace, D.B.; Gulyaz, V.; Tuppurainen, E.; Madani, H.; Caufour, P.; Adam, T.; El Harrak, M.; et al. Use of the Capripoxvirus homologue of Vaccinia virus 30 kDa RNA polymerase subunit (RPO30) gene as a novel diagnostic and genotyping target: Development of a classical PCR method to differentiate Goat poxvirus from Sheep poxvirus. Vet. Microbiol. 2011, 149, 30–39. [Google Scholar] [CrossRef]
- Zhao, Z.; Wu, G.; Yan, X.; Zhu, X.; Li, J.; Zhu, H.; Zhang, Z.; Zhang, Q. Development of duplex PCR for differential detection of goatpox and sheeppox viruses. BMC Vet. Res. 2017, 13, 278. [Google Scholar] [CrossRef] [Green Version]
- Gelaye, E.; Lamien, C.E.; Silber, R.; Tuppurainen, E.S.; Grabherr, R.; Diallo, A. Development of a cost-effective method for capripoxvirus genotyping using snapback primer and dsDNA intercalating dye. PLoS ONE 2013, 8, e75971. [Google Scholar] [CrossRef] [Green Version]
- Pestova, Y.; Byadovskaya, O.; Kononov, A.; Sprygin, A. A real time high-resolution melting PCR assay for detection and differentiation among sheep pox virus, goat pox virus, field and vaccine strains of lumpy skin disease virus. Mol. Cell Probes. 2018, 41, 57–60. [Google Scholar] [CrossRef]
- Tulman, E.R.; Afonso, C.L.; Lu, Z.; Zsak, L.; Sur, J.H.; Sandybaev, N.T.; Kerembekova, U.Z.; Zaitsev, V.L.; Kutish, G.F.; Rock, D.L. The genomes of sheeppox and goatpox viruses. J. Virol. 2002, 76, 6054–6061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forth, J.H.; Forth, L.F.; King, J.; Groza, O.; Hubner, A.; Olesen, A.S.; Hoper, D.; Dixon, L.K.; Netherton, C.L.; Rasmussen, T.B.; et al. A Deep-Sequencing Workflow for the Fast and Efficient Generation of High-Quality African Swine Fever Virus Whole-Genome Sequences. Viruses 2019, 11, 846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolff, J.; Krstevski, K.; Beer, M.; Hoffmann, B. Minimum Infective Dose of a Lumpy Skin Disease Virus Field Strain from North Macedonia. Viruses 2020, 12, 768. [Google Scholar] [CrossRef] [PubMed]
- Carn, V.M.; Kitching, R.P. The clinical response of cattle experimentally infected with lumpy skin disease (Neethling) virus. Arch. Virol. 1995, 140, 503–513. [Google Scholar] [CrossRef]
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods 2006, 136, 200–209. [Google Scholar] [CrossRef]
- Dietze, K.; Moritz, T.; Alexandrov, T.; Krstevski, K.; Schlottau, K.; Milovanovic, M.; Hoffmann, D.; Hoffmann, B. Suitability of group-level oral fluid sampling in ruminant populations for lumpy skin disease virus detection. Vet. Microbiol. 2018, 221, 44–48. [Google Scholar] [CrossRef]
- Spearman, C. The Method of ‘Right and Wrong Cases’ (‘Constant Stimuli’) without Gauss’s Formulae. Br. J. Psychol. 1904–1920 1908, 2, 227–242. [Google Scholar] [CrossRef]
- Kärber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Naunyn-Schmiedebergs Arch. Exp. Pathol. Pharmakol. 1931, 162, 480–483. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Biswas, S.; Noyce, R.S.; Babiuk, L.A.; Lung, O.; Bulach, D.M.; Bowden, T.R.; Boyle, D.B.; Babiuk, S.; Evans, D.H. Extended sequencing of vaccine and wild-type capripoxvirus isolates provides insights into genes modulating virulence and host range. Transbound. Emerg. Dis. 2020, 67, 80–97. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, J.; Bamouh, Z.; Jazouli, M.; Boumart, Z.; Tadlaoui, K.O.; Fihri, O.F.; El Harrak, M. Experimental evaluation of the cross-protection between Sheeppox and bovine Lumpy skin vaccines. Sci. Rep. 2020, 10, 8888. [Google Scholar] [CrossRef] [PubMed]
- Boumart, Z.; Daouam, S.; Belkourati, I.; Rafi, L.; Tuppurainen, E.; Tadlaoui, K.O.; El Harrak, M. Comparative innocuity and efficacy of live and inactivated sheeppox vaccines. BMC Vet. Res. 2016, 12, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Möller, J.; Moritz, T.; Schlottau, K.; Krstevski, K.; Hoffmann, D.; Beer, M.; Hoffmann, B. Experimental lumpy skin disease virus infection of cattle: Comparison of a field strain and a vaccine strain. Arch. Virol. 2019, 164, 2931–2941. [Google Scholar] [CrossRef] [PubMed]
- Abutarbush, S.M.; Hananeh, W.M.; Ramadan, W.; Al Sheyab, O.M.; Alnajjar, A.R.; Al Zoubi, I.G.; Knowles, N.J.; Bachanek-Bankowska, K.; Tuppurainen, E.S. Adverse Reactions to Field Vaccination Against Lumpy Skin Disease in Jordan. Transbound. Emerg. Dis. 2016, 63, e213–e219. [Google Scholar] [CrossRef]
- Katsoulos, P.D.; Chaintoutis, S.C.; Dovas, C.I.; Polizopoulou, Z.S.; Brellou, G.D.; Agianniotaki, E.I.; Tasioudi, K.E.; Chondrokouki, E.; Papadopoulos, O.; Karatzias, H.; et al. Investigation on the incidence of adverse reactions, viraemia and haematological changes following field immunization of cattle using a live attenuated vaccine against lumpy skin disease. Transbound. Emerg. Dis. 2018, 65, 174–185. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| SPPV-“V/104“ | Capri-p32-Mix 1-Taq-FAM | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| dpi | ||||||||||||
| 0 | 3 | 5 | 7 | 10 | 13 | 15 | 18 | 21 | 23/24 | 28 | ||
| Animal | matrix | |||||||||||
| S/542 i.v. + s.c. | EDTA-blood | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq |
| serum | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| nasal swab | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| oral swab | no Cq | no Cq | no Cq | no Cq | n.a. | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| S/573 i.v. + s.c. | EDTA-blood | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq |
| serum | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| nasal swab | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| oral swab | no Cq | no Cq | no Cq | no Cq | n.a. | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| S/643 i.v. + s.c. | EDTA-blood | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq |
| serum | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| nasal swab | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| oral swab | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| S/652 i.n. | EDTA-blood | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq |
| serum | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| nasal swab | no Cq | 36.4 | no Cq | no Cq | n.a. | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| oral swab | no Cq | no Cq | 36.3 | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| S/673 i.n. | EDTA-blood | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq |
| serum | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| nasal swab | no Cq | 38.0 | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| oral swab | no Cq | no Cq | no Cq | no Cq | n.a. | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| S/718 i.n. | EDTA-blood | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq |
| serum | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| nasal swab | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| oral swab | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| S/724 i-c | EDTA-blood | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq |
| serum | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| nasal swab | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| oral swab | no Cq | no Cq | no Cq | no Cq | n.a. | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| S/725 i-c | EDTA-blood | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq |
| serum | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| nasal swab | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| oral swab | no Cq | no Cq | no Cq | no Cq | n.a. | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | |
| GTPV-“V/103“ | Capri-p32-Mix 1-Taq-FAM | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| dpi | ||||||||||||
| 0 | 3 | 5 | 7 | 10 | 13 | 15 | 18 | 21 | 23/24 | 28 | ||
| Animal | matrix | |||||||||||
| Z/253 i.v. + s.c. | EDTA-blood | no Cq | no Cq | 36.4 | 34.3 | 32.1 | - | - | - | - | - | - |
| serum | no Cq | no Cq | no Cq | no Cq | 37.5 | - | - | - | - | - | - | |
| nasal swab | no Cq | no Cq | 39.5 | no Cq | 27.9 | - | - | - | - | - | - | |
| oral swab | no Cq | no Cq | no Cq | no Cq | 34.8 | - | - | - | - | - | - | |
| Z/254 i.v. + s.c. | EDTA-blood | no Cq | no Cq | 36.6 | 26.8 | 27.2 | - | - | - | - | - | - |
| serum | no Cq | no Cq | no Cq | 37.7 | 34.9 | - | - | - | - | - | - | |
| nasal swab | no Cq | no Cq | no Cq | 37.0 | 26.6 | - | - | - | - | - | - | |
| oral swab | no Cq | no Cq | no Cq | no Cq | 32.2 | - | - | - | - | - | - | |
| Z/255 i.v. + s.c. | EDTA-blood | no Cq | no Cq | 34.4 | 30.1 | 27.5 | - | - | - | - | - | - |
| serum | no Cq | no Cq | no Cq | no Cq | 36.3 | - | - | - | - | - | - | |
| nasal swab | no Cq | no Cq | 36.1 | no Cq | 28.4 | - | - | - | - | - | - | |
| oral swab | no Cq | no Cq | no Cq | no Cq | 34.4 | - | - | - | - | - | - | |
| Z/256 i.n. | EDTA-blood | no Cq | no Cq | no Cq | no Cq | 30.3 | 29.2 | 31.2 | - | - | - | - |
| serum | no Cq | no Cq | no Cq | no Cq | no Cq | 35.6 | 37.9 | - | - | - | - | |
| nasal swab | no Cq | no Cq | 27.02 | 23.6 | 18.9 | 19.0 | 19.6 | - | - | - | - | |
| oral swab | no Cq | no Cq | no Cq | 28.6 | 27.4 | 26.0 | 25.7 | - | - | - | - | |
| Z/257 i.n. | EDTA-blood | no Cq | no Cq | no Cq | 34.4 | 33.6 | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq |
| serum | no Cq | no Cq | no Cq | n.a. | no Cq | no Cq | n.a. | no Cq | no Cq | no Cq | no Cq | |
| nasal swab | no Cq | 37.7 | 26.3 | 21.4 | 19.9 | 19.1 | 20.4 | 21.5 | 22.4 | 26.6 | 26.1 | |
| oral swab | no Cq | no Cq | 36.9 | 32.0 | 27.8 | 27.7 | 26.4 | 30.2 | 29.6 | 32.1 | 36.1 | |
| Z/258 i.n. | EDTA-blood | no Cq | no Cq | no Cq | no Cq | 33.0 | 35.0 | no Cq | no Cq | no Cq | no Cq | no Cq |
| serum | no Cq | no Cq | no Cq | no Cq | 37.4 | 37.2 | no Cq | no Cq | no Cq | no Cq | no Cq | |
| nasal swab | no Cq | 38.1 | 29.4 | 24.0 | 24.0 | 19.8 | 21.5 | 25.4 | 25.0 | 24.4 | 33.4 | |
| oral swab | no Cq | no Cq | 36.8 | 33.6 | 31.2 | 31.0 | 28.9 | 29.8 | 32.7 | 31.2 | 35.7 | |
| Z/259 i-c | EDTA-blood | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | 29.8 | 27.8 | - |
| serum | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | 37.3 | 31.4 | - | |
| nasal swab | no Cq | no Cq | no Cq | no Cq | no Cq | 34.1 | 33.8 | 35.1 | 21.1 | 20.5 | - | |
| oral swab | no Cq | no Cq | no Cq | no Cq | no Cq | 34.4 | 36.0 | 33.4 | 33.6 | 30.3 | - | |
| Z/260 i-c | EDTA-blood | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | 35.0 | 29.6 |
| serum | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | 35.9 | |
| nasal swab | no Cq | no Cq | no Cq | no Cq | 31.4 | 36.4 | 33.7 | 33.5 | 31.8 | 34.1 | 20.4 | |
| oral swab | no Cq | no Cq | 35.8 | no Cq | 37.3 | 33.3 | 37.8 | 31.7 | 33.9 | 33.9 | 27.9 | |
| GTPV-“V/103“ | Capri p32 Taq Mix FAM | |||||||
|---|---|---|---|---|---|---|---|---|
| Organ Sample | Intravenously + Subcutaneously | Intranasal | In-Contact | |||||
| Z/253 | Z/254 | Z/255 | Z/256 | Z/257 | Z/258 | Z/259 | Z/260 | |
| mandibular lymph node | 30.4 | 33.4 | 34.1 | 31.0 | 35.1 | no Cq | 21.4 | 28.3 |
| cervical lymph node | 29.7 | 30.1 | 27.9 | 27.2 | no Cq | no Cq | 20.2 | 25.5 |
| mediastinal lymph node | 31.7 | 31.8 | 34.2 | 37.0 | 36.7 | no Cq | 23.1 | 32.2 |
| mesenterial lymph node | 36.5 | no Cq | no Cq | no Cq | no Cq | no Cq | 27.4 | 34.2 |
| tonsil | 36.2 | no Cq | no Cq | 32.0 | no Cq | no Cq | 24.8 | 28.3 |
| spleen | 32.9 | 36.2 | 35.3 | 31.5 | no Cq | no Cq | 19.8 | 32.0 |
| liver | 36.8 | 33.9 | no Cq | 35.5 | no Cq | no Cq | 22.8 | 33.7 |
| lung | 21.9 | 26.8 | 21.6 | 28.3 | no Cq | no Cq | 19.2 | 26.2 |
| heart | no Cq | no Cq | no Cq | 34.9 | no Cq | no Cq | 30.7 | 35.6 |
| kidney | 35.2 | 37.0 | 35.6 | 36.2 | no Cq | no Cq | 23.6 | 30.4 |
| salivary gland | no Cq | no Cq | no Cq | no Cq | no Cq | no Cq | 34.5 | 31.4 |
| testicles | 29.4 | 30.2 | 27.3 | 25.3 | 37.5 | 37.5 | 29.2 | 26.1 |
| oral mucosa | 36.9 | 32.3 | 32.1 | 27.0 | 31.3 | 33.4 | 23.5 | 28.4 |
| thigh muscle | no Cq | 37.0 | no Cq | 38.1 | no Cq | no Cq | 34.2 | 32.4 |
| nose/conchae | n. t. | n. t. | n. t. | 17.8 | 27.7 | 37.5 | 22.4 | 21.3 |
| trachea | 19.4 | n. t. | 26.2 | 21.6 | 28.1 | n. t. | 19.9 | n. t. |
| rumen | n. t. | n. t. | 33.0 | n. t. | n. t. | n. t. | 27.5 | n. t. |
| abomasum | n. t. | n. t. | n. t. | n. t. | n. t. | n. t. | 22.4 | n. t. |
| epididymis | n. t. | n. t. | n. t. | 18.5 | n. t. | n. t. | n. t. | n. t. |
| location of skin sample | ||||||||
| mouth/nose | 22.6 | 20.5 | 23.3 | 20.7 | no Cq | 36.6 | 20.7 | 21.4 |
| eye/lid edge | 25.3 | 22.3 | 25.0 | 20.2 | no Cq | 36.8 | 19.8 | 19.0 |
| closely behind the ear | 20.3 | 20.8 | 29.9 | 20.7 | 33.6 | 31.4 | 20.5 | 21.1 |
| thoracic wall | 18.9 | 20.8 | 29.6 | 21.5 | 33.0 | 21.3 | 21.2 | 19.1 |
| skin inguinal | n. t. | 23.5 | 25.1 | 18.8 | n. t. | n. t. | n. t. | n. t. |
| hind leg | n. t. | n. t. | n. t. | n. t. | n. t. | 21.3 | n. t. | n. t. |
| tail base | n. t. | n. t. | n. t. | 16.9 | n. t. | n. t. | n. t. | n. t. |
| prepuce | n. t. | n. t. | n. t. | 19.1 | n. t. | n. t. | n. t. | n. t. |
| scrotum | 22.3 | n. t. | n. t. | n. t. | 32.5 | n. t. | 24.4 | 19.8 |
| SPPV-“V/104“ | Serological Examination | ||
|---|---|---|---|
| ELISA (S/P%) | SNT (Titer) | ||
| S/542 i.v. + s.c. | 0 dpi | −1 | < 1:10 |
| 7 dpi | 14 | 1:13 | |
| 15 dpi | 23 | 1:20 | |
| 21 dpi | 51 | 1:25 | |
| 28 dpi | 129 | 1:50 | |
| S/573 i.v. + s.c. | 0 dpi | 0 | < 1:10 |
| 7 dpi | 3 | < 1:10 | |
| 15 dpi | 0 | < 1:10 | |
| 21 dpi | 0 | 1:13 | |
| 28 dpi | 7 | 1:13 | |
| S/643 i.v. + s.c. | 0 dpi | −1 | < 1:10 |
| 7 dpi | 9 | < 1:10 | |
| 15 dpi | 1 | < 1:10 | |
| 21 dpi | 3 | < 1:10 | |
| 28 dpi | 3 | < 1:10 | |
| S/652 i.n. | 0 dpi | −1 | < 1:10 |
| 7 dpi | 1 | < 1:10 | |
| 15 dpi | 4 | < 1:10 | |
| 21 dpi | 0 | < 1:10 | |
| 28 dpi | 0 | < 1:10 | |
| S/673 i.n. | 0 dpi | −1 | < 1:10 |
| 7 dpi | −1 | < 1:10 | |
| 15 dpi | 0 | < 1:10 | |
| 21 dpi | 0 | < 1:10 | |
| 28 dpi | 1 | < 1:10 | |
| S/718 i.n. | 0 dpi | −1 | < 1:10 |
| 7 dpi | −1 | < 1:10 | |
| 15 dpi | −1 | < 1:10 | |
| 21 dpi | −1 | < 1:10 | |
| 28 dpi | −1 | < 1:10 | |
| S/724 i-c | 0 dpi | −1 | < 1:10 |
| 7 dpi | −1 | < 1:10 | |
| 15 dpi | −1 | < 1:10 | |
| 21 dpi | −1 | < 1:10 | |
| 28 dpi | 0 | < 1:10 | |
| S/725 i-c | 0 dpi | 6 | < 1:10 |
| 7 dpi | −1 | < 1:10 | |
| 15 dpi | −1 | < 1:10 | |
| 21 dpi | 0 | < 1:10 | |
| 28 dpi | 0 | < 1:10 | |
| GTPV-“V/103“ | Serological Examination | ||
|---|---|---|---|
| ELISA (S/P%) | SNT (Titer) | ||
| Z/253 i.v. + s.c. | 0 dpi | −1 | <1:10 |
| 7 dpi | −1 | <1:10 | |
| 10 dpi | −1 | 1:13 | |
| Z/254 i.v. + s.c. | 0 dpi | −1 | <1:10 |
| 7 dpi | 0 | 1:13 | |
| 10 dpi | 16 | 1:80 | |
| Z/255 i.v. + s.c. | 0 dpi | 0 | <1:10 |
| 7 dpi | 0 | <1:10 | |
| 10 dpi | 53 | 1:25 | |
| Z/256 i.n. | 0 dpi | −1 | <1:10 |
| 7 dpi | −1 | <1:10 | |
| 15 dpi | 23 | 1:16 | |
| Z/257 i.n. | 0 dpi | −1 | <1:10 |
| 7 dpi | −1 | <1:10 | |
| 15 dpi | 11 | 1:50 | |
| 21 dpi | 80 | 1:160 | |
| 28 dpi | 83 | 1:80 | |
| Z/258 i.n. | 0 dpi | −1 | <1:10 |
| 7 dpi | −1 | <1:10 | |
| 15 dpi | 1 | 1:80 | |
| 21 dpi | 15 | 1:256 | |
| 28 dpi | 65 | 1:128 | |
| Z/259 i-c | 0 dpi | 6 | <1:10 |
| 7 dpi | 1 | <1:10 | |
| 15 dpi | −1 | <1:10 | |
| 21 dpi | −1 | <1:10 | |
| 23 dpi | 0 | 1:40 | |
| Z/260 i-c | 0 dpi | −1 | <1:10 |
| 7 dpi | −1 | <1:10 | |
| 15 dpi | −1 | <1:10 | |
| 21 dpi | 0 | <1:10 | |
| 28 dpi | 15 | 1:13 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wolff, J.; King, J.; Moritz, T.; Pohlmann, A.; Hoffmann, D.; Beer, M.; Hoffmann, B. Experimental Infection and Genetic Characterization of Two Different Capripox Virus Isolates in Small Ruminants. Viruses 2020, 12, 1098. https://0-doi-org.brum.beds.ac.uk/10.3390/v12101098
Wolff J, King J, Moritz T, Pohlmann A, Hoffmann D, Beer M, Hoffmann B. Experimental Infection and Genetic Characterization of Two Different Capripox Virus Isolates in Small Ruminants. Viruses. 2020; 12(10):1098. https://0-doi-org.brum.beds.ac.uk/10.3390/v12101098
Chicago/Turabian StyleWolff, Janika, Jacqueline King, Tom Moritz, Anne Pohlmann, Donata Hoffmann, Martin Beer, and Bernd Hoffmann. 2020. "Experimental Infection and Genetic Characterization of Two Different Capripox Virus Isolates in Small Ruminants" Viruses 12, no. 10: 1098. https://0-doi-org.brum.beds.ac.uk/10.3390/v12101098