
Inhibition of Enterovirus A71 by a Novel 2-Phenyl-Benzimidazole Derivative

 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
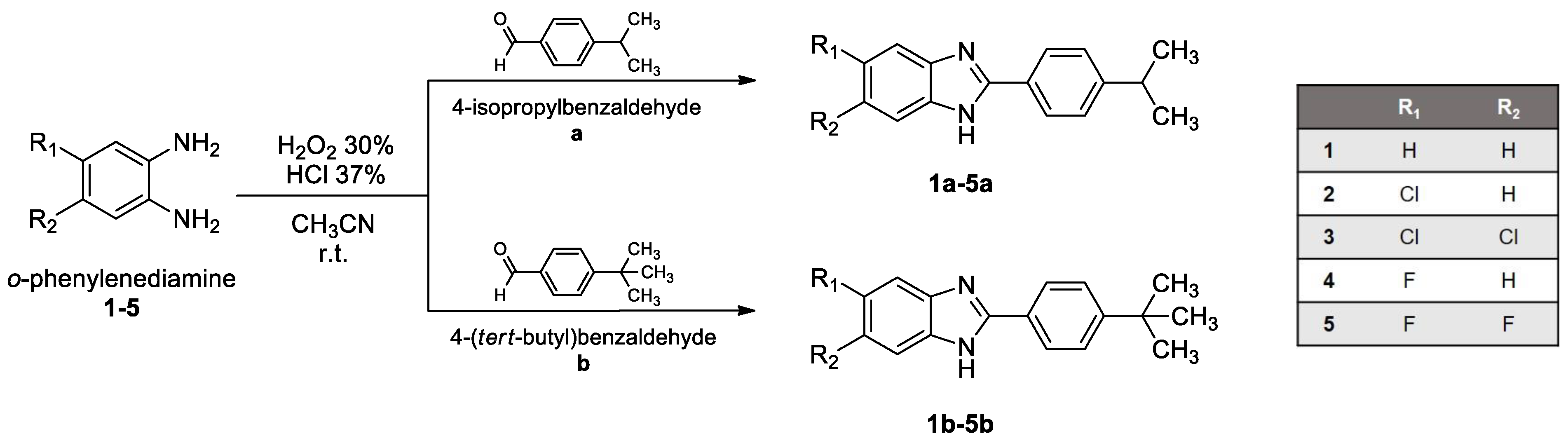
2.1. Chemistry
2.1.1. Synthesis
2.1.2. Chemical Characterization
2.2. Biology
2.2.1. Cells and Viruses
2.2.2. Cytotoxicity Assays
2.2.3. Antiviral Assays
2.2.4. Yield Reduction Assay
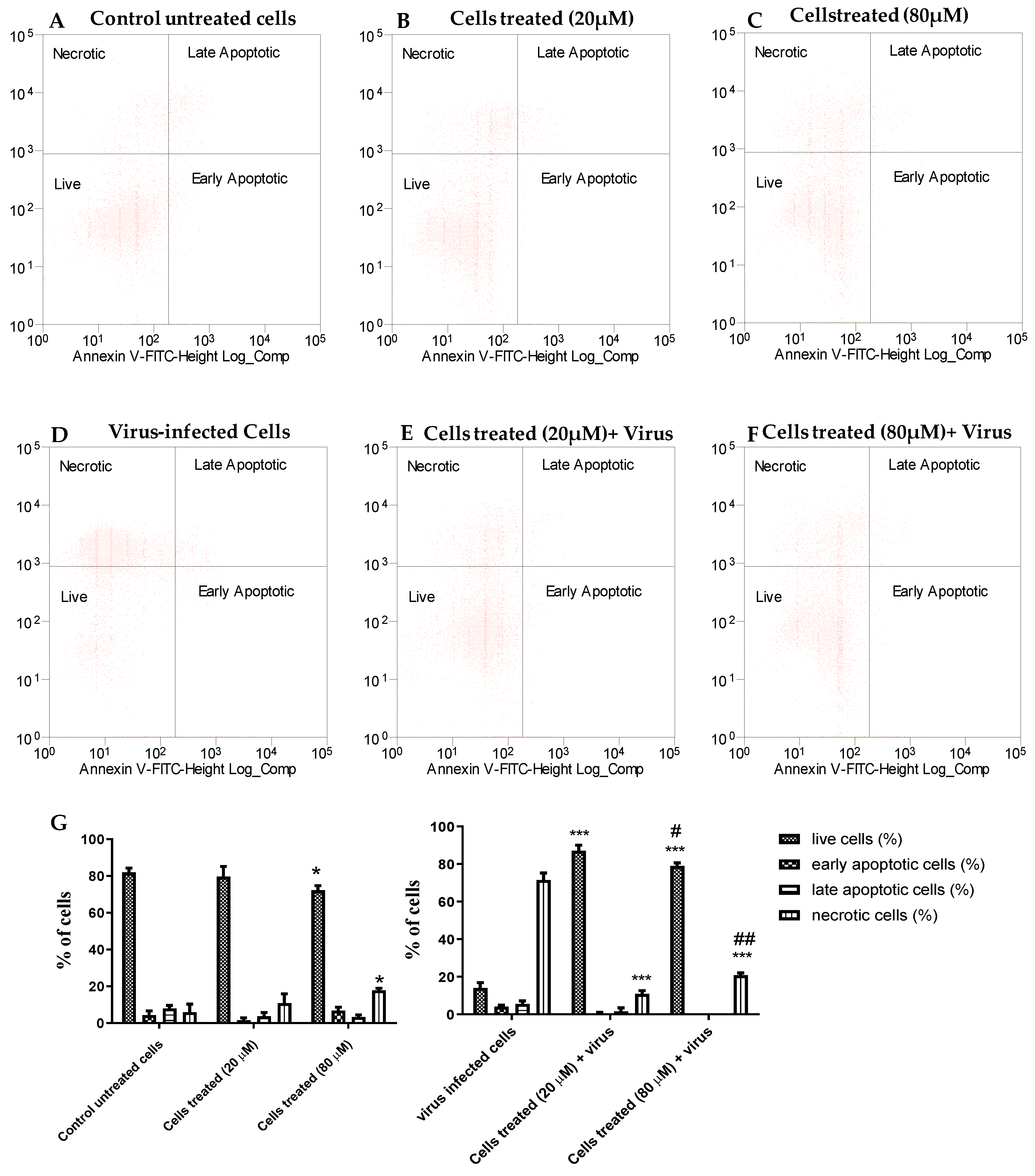
2.2.5. Apoptosis Assay
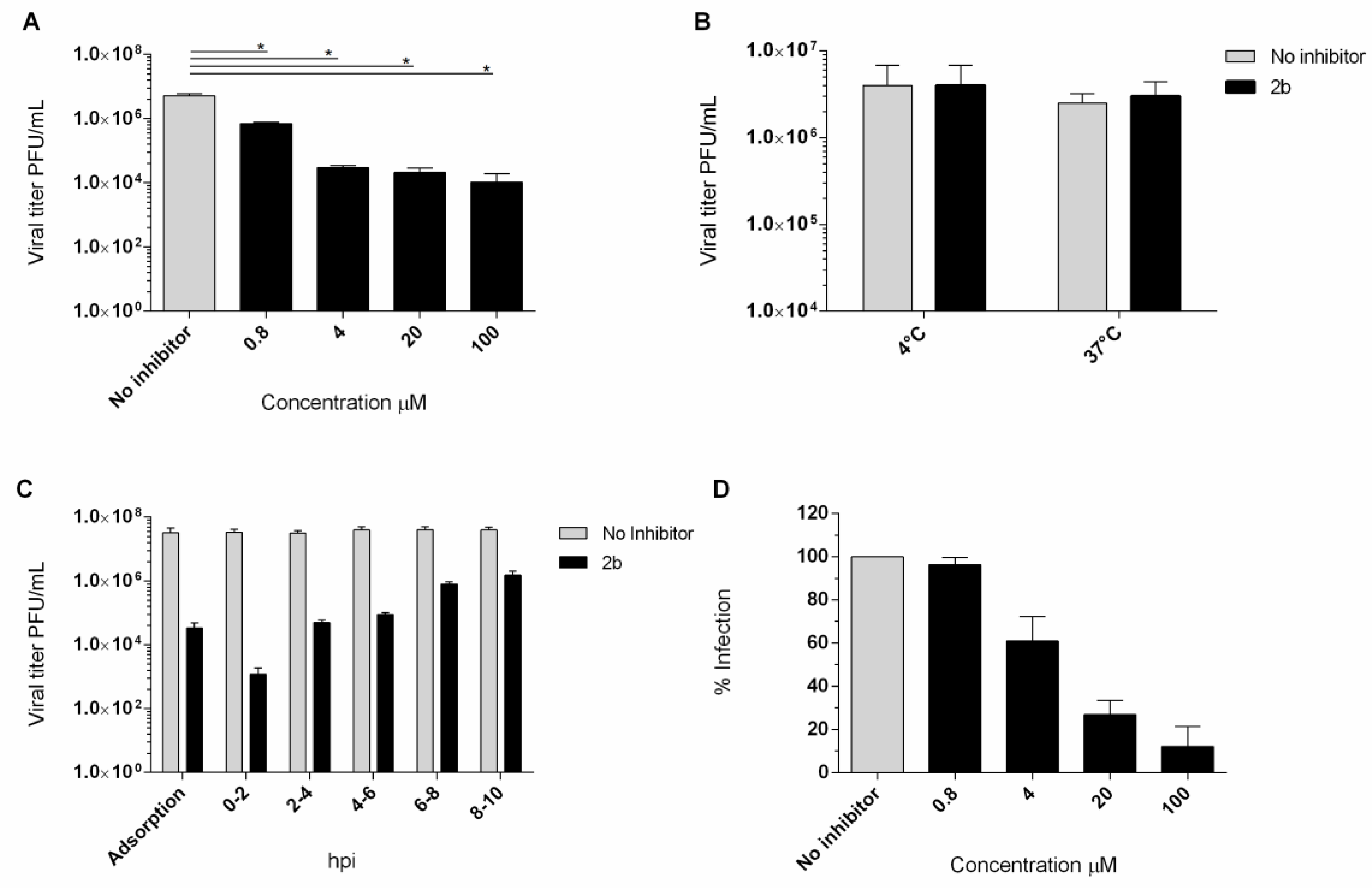
2.2.6. Virucidal Activity Assay
2.2.7. Cell Pretreatment Assay
2.2.8. Adsorption Assays
2.2.9. Time-of-Addition Assay
2.2.10. Penetration Assay
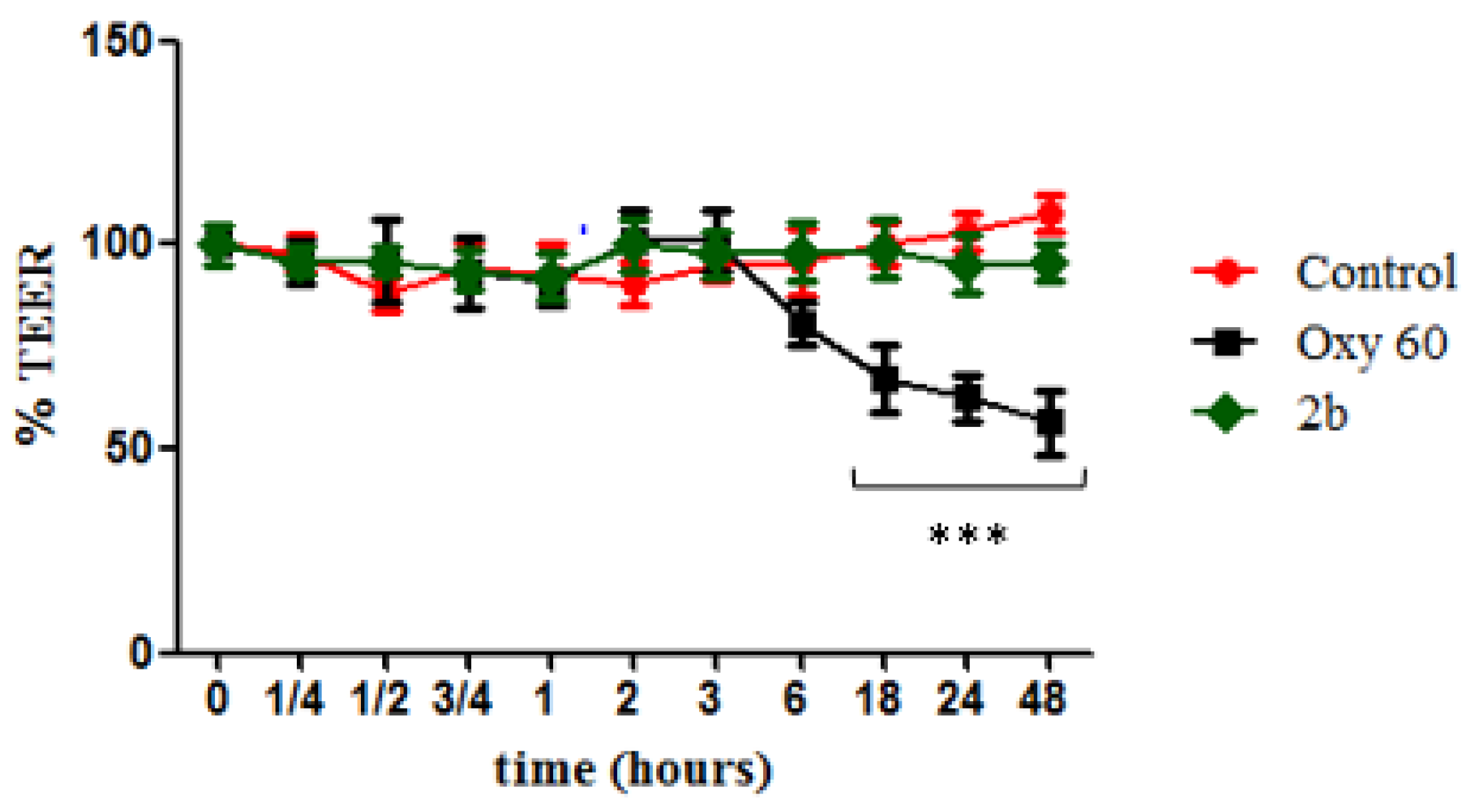
2.2.11. Evaluation of Cell Monolayer Permeabilization (TEER)
2.2.12. Statistical Analysis
2.2.13. Linear Regression Analysis
2.3. In Silico Screening
3. Experimental Section
3.1. 2-(4-Isopropylphenyl)-1H-Benzo[d]imidazole (1a)
3.2. 5-Chloro-2-(4-Isopropylphenyl)-1H-Benzo[d]imidazole (2a)
3.3. 5,6-Dichloro-2-(4-Isopropylphenyl)-1H-Benzo[d]imidazole (3a)
3.4. 5-Fluoro-2-(4-Isopropylphenyl)-1H-Benzo[d]imidazole (4a)
3.5. 5,6-Difluoro-2-(4-Isopropylphenyl)-1H-Benzo[d]imidazole (5a)
3.6. 2-(4-(Tert-Butyl)phenyl)-1H-Benzo[d]imidazole (1b)
3.7. 2-(4-(Tert-Butyl)phenyl)-5-Chloro-1H-Benzo[d]imidazole (2b)
3.8. 2-(4-(Tert-Butyl)phenyl)-5,6-Dichloro-1H-Benzo[d]imidazole (3b)
3.9. 2-(4-(Tert-Butyl)phenyl)-5-Fluoro-1H-Benzo[d]imidazole (4b)
3.10. 2-(4-(Tert-Butyl)phenyl)-5,6-Difluoro-1H-Benzo[d]imidazole (5b)
4. Results
4.1. Chemistry
4.2. Antiviral Assays
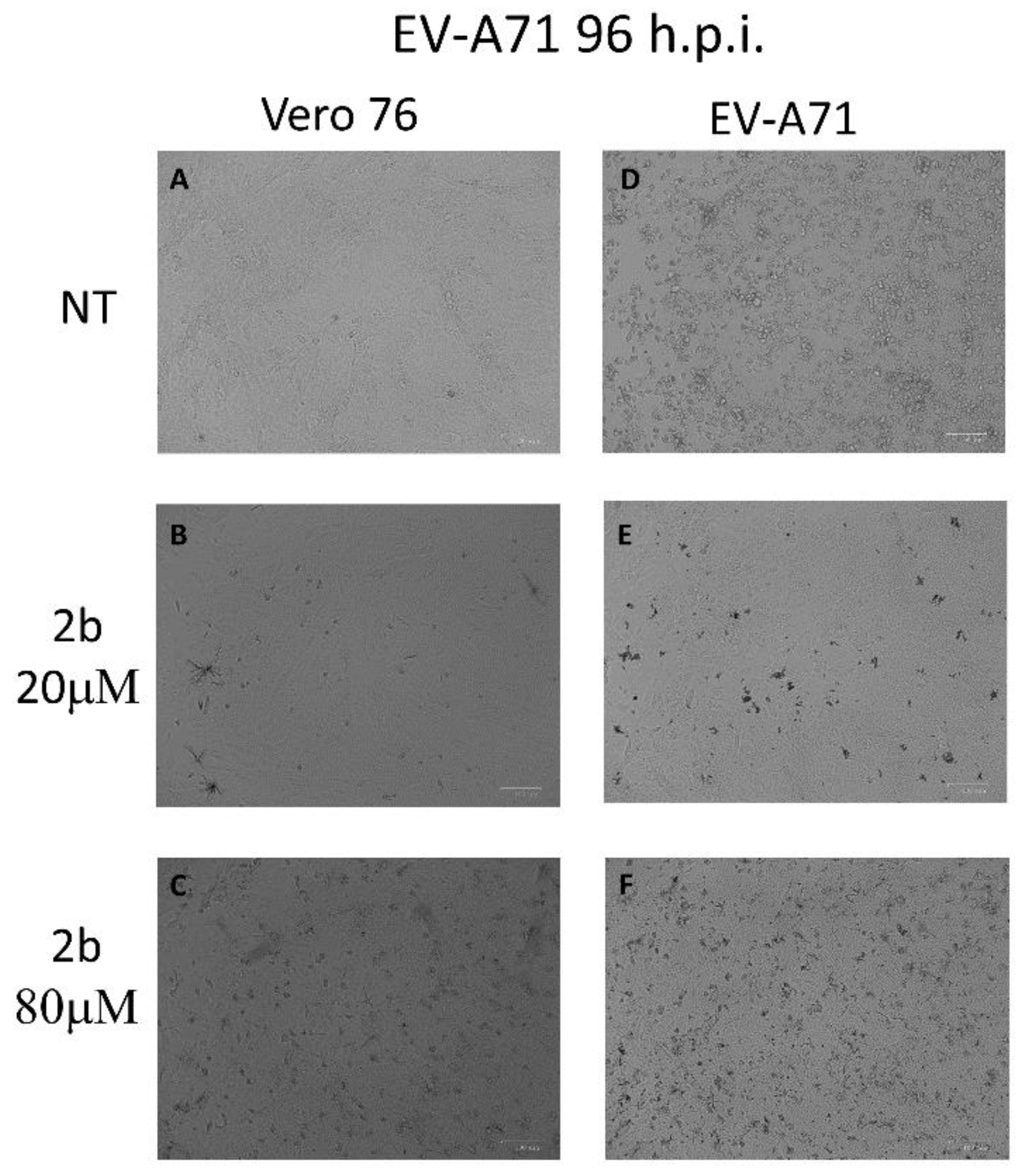
4.3. Protective Effect of 2b on EV-A71 Infected Cells
4.4. TEER Experiment
4.5. Yield Reduction Assay and Study of the Mechanism of Action
4.6. Druglikeness, Pharmacokinetic and Toxicity Predictions
5. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hsing-I, H.; Shin-Ru, S. Neurotropic Enterovirus Infections in the Central Nervous System. Viruses 2015, 7, 6051–6066. [Google Scholar]
- Jane, Q. Enterovirus 71 infection: A new threat to global public health? Lancet Neurol. 2008, 7, 868–869. [Google Scholar]
- Xing, W.; Liao, Q.; Viboud, C.; Zhang, J.; Sun, J.; Wu, J.T.; Chang, Z.; Liu, F.; Fang, V.J.; Zheng, Y.; et al. Hand, foot, and mouth disease in China, 2008–2012: An epidemiological study. Lancet Infect. Dis. 2014, 14, 308–318. [Google Scholar] [CrossRef] [Green Version]
- Puenpa, J.; Wanlapakorn, N.; Vongpunsawad, S.; Poovorawan, Y. The History of Enterovirus A71 Outbreaks and Molecular Epidemiology in the Asia-Pacific Region. J. Biomed. Sci. 2019, 26, 75. [Google Scholar] [CrossRef]
- Wang, S.M.; Liu, C.C. Update of enterovirus 71 infection: Epidemiology, pathogenesis and vaccine. Expert Rev. Anti Infect. Ther. 2014, 12, 447–456. [Google Scholar] [CrossRef]
- Lugo, D.; Krogstad, P. Enteroviruses in the early 21st century: New manifestations and challenges. Curr. Opin. Pediatr. 2016, 28, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.S.; Lee, H.C.; Lee, K.M.; Gong, Y.N.; Shih, S.R. Enterovirus and Encephalitis. Front. Microbiol. 2020. [Google Scholar] [CrossRef]
- Emerging Infectious Diseases. Available online: www.cdc.gov/eid (accessed on 22 September 2016).
- Yi, E.J.; Shin, Y.J.; Kim, J.H.; Kim, T.G.; Chang, S.Y. Enterovirus 71 infection and vaccines. Clin. Exp. Vaccine Res. 2017, 6, 4–14. [Google Scholar] [CrossRef]
- Lin, J.Y.; Kung, Y.A.; Shih, S.R. Antivirals and vaccines for Enterovirus A71. J. Biomed. Sci. 2019, 26, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.N.; Yao, C.T.; Yeh, C.N.; Huang, C.C.; Wang, S.M.; Liu, C.C.; Wu, J.M. Critical management in patients with severe enterovirus 71 infection. Pediatrics Int. 2006, 48, 250–256. [Google Scholar] [CrossRef]
- Ooi, M.H.; Wong, S.C.; Lewthwaite, P.; Cardosa, M.J.; Solomon, T. Clinical features, diagnosis, and management of enterovirus 71. Lancet Neurol. 2010, 9, 1097–1105. [Google Scholar] [CrossRef]
- Carta, A.; Loriga, G.; Piras, S.; Paglietti, G.; Ferrone, M.; Fermeglia, M.; Pricl, S.; La Colla, P.; Secci, B.; Collu, G.; et al. Synthesis and In vitro Evaluation of the Anti-Viral Activity of N-[4-(1H(2H)-benzotriazol-1(2)-yl)phenyl]alkylcarboxamides. Med. Chem. 2006, 2, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Vitale, G.; Corona, P.; Loriga, M.; Carta, A.; Paglietti, G.; Giliberti, G.; Sanna, G.; Farci, P.; Marongiu, M.E.; La Colla, P. 5-Acetyl-2-arylbenzimidazoles asantiviral agents. Part 4. Eur. J. Med. Chem. 2012, 53, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Briguglio, I.; Loddo, R.; Laurini, E.; Fermeglia, M.; Piras, S.; Corona, P.; Giunchedi, P.; Gavini, E.; Sanna, G.; Giliberti, G.; et al. Synthesis, cytotoxicity and antiviral evaluation of new series of imidazo[4,5-g[quinoline and pyrido[2,3-g[quinoxalinone derivatives. Eur. J. Med. Chem. 2015, 105, 63–79. [Google Scholar] [CrossRef] [PubMed]
- Sanna, P.; Carta, A.; Paglietti, G.; Zanetti, S.; Fadda, G. 1,2,3-triazolo[4,5-h]quinolines. III. Preparation and antimicrobial evaluation of 4-ethyl-4,7-dihydro-1(2)-R-1(2)H triazolo[4,5-h]quinolin-7-one-6-carboxylic acids as anti-infectives of the urinary tract. Farmaco 1992, 47, 1001–1019. [Google Scholar]
- Corona, P.; Gibellini, F.; Cavalli, A.; Saxena, P.; Carta, A.; Loriga, M.; Luciani, R.; Paglietti, G.; Guerrieri, D.; Nerini, E.; et al. Structure-Based Selectivity Optimization of Piperidine–Pteridine Derivatives as Potent Leishmania Pteridine Reductase Inhibitors. J. Med. Chem. 2012, 55, 8318–8329. [Google Scholar] [CrossRef]
- Piras, S.; Corona, P.; Ibba, R.; Riu, F.; Murineddu, G.; Sanna, G.; Madeddu, S.; Delogu, I.; Loddo, R.; Carta, A. Preliminary anti-Coxsackie activity of novel 1-[4-5,6-dimethyl(H)-1H(2H)-benzotriazol-1(2)-yl)phenyl]-3-alkyl(aryl)ureas. Med. Chem. 2020, 16, 677–688. [Google Scholar] [CrossRef]
- Piras, S.; Sanna, G.; Carta, A.; Corona, P.; Ibba, R.; Loddo, R.; Madeddu, S.; Caria, P.; Aulic, S.; Laurini, E.; et al. Dichloro-phenyl-benzotriazoles: A new selective class of Human Respiratory Syncytial Virus entry inhibitors. Front. Chem. 2019, 16, 247. [Google Scholar] [CrossRef]
- Bahrami, K.; Khodaei, M.M.; Kavianinia, I. A simple and efficient one-pot synthesis of 2-substituted benzimidazoles. Synthesis 2007, 38, 547–550. [Google Scholar] [CrossRef]
- Serreli, G.; Melis, M.P.; Corona, G.; Deiana, M. Modulation of LPS-induced nitric oxide production in intestinal cells by hydroxytyrosol and tyrosol metabolites: Insight into the mechanism of action. Food Chem. Toxicol. 2019, 125, 520–527. [Google Scholar] [CrossRef] [Green Version]
- Sanna, G.; Madeddu, S.; Murgia, G.; Serreli, G.; Begala, M.; Caboni, P.; Incani, A.; Franci, G.; Galdiero, M.; Giliberti, G. Potent and Selective Activity against Human Immunodeficiency Virus 1 (HIV-1) of Thymelaeahirsuta Extracts. Viruses 2020, 12, 664. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, R.; Balzarini, J.; Baba, M.; Snoeck, R.; Schols, D.; Herdewijn, P.; Desmyter, J.; De Clercq, E. Rapid and automated tetrazolium-based colorimetric assay for the detection of anti-HIV compounds. J. Virol. Methods 1988, 20, 309–321. [Google Scholar] [CrossRef]
- Carta, A.; Sanna, G.; Briguglio, I.; Madeddu, S.; Vitale, G.; Piras, S.; Corona, P.; Peana, A.T.; Laurini, E.; Fermeglia, M.; et al. Quinoxaline derivatives as new inhibitors of coxsackievirus B5. Eur. J. Med. Chem. 2018, 145, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-W.; Leu, Y.-L.; Horng, J.-T. Daphne GenkwaSieb. et Zucc. Water-Soluble Extracts Act on Enterovirus 71 by Inhibiting Viral Entry. Viruses 2012, 4, 539–556. [Google Scholar] [CrossRef] [Green Version]
- Rosenthal, K.S.; Perez, R.; Hodnichak, C. Inhibition of herpes simplex virus type 1 penetration by cytochalasins B and D. J. Gen. Virol. 1985, 66, 1601–1605. [Google Scholar] [CrossRef]
- Serreli, G.; Incani, A.; Atzeri, A.; Angioni, A.; Campus, M.; Cauli, E.; Zurru, R.; Deiana, M. Antioxidant Effect of Natural Table Olives Phenolic Extract Against Oxidative Stress and Membrane Damage in Enterocyte-Like Cells. J. Food Sci. 2017, 82, 380–385. [Google Scholar] [CrossRef]
- Serra, G.; Incani, A.; Serreli, G.; Porru, L.; Melis, M.P.; Tuberoso, C.I.G.; Rossin, D.; Biasi, F.; Deiana, M. Olive oil polyphenols reduce oxysterols -induced redox imbalance and pro-inflammatory response in intestinal cells. Redox. Biol. 2018, 17, 348–354. [Google Scholar] [CrossRef]
- Van Noord, C.; Sturkenboom, M.C.; Straus, S.M.; Witteman, J.C.; Stricker, B.H. Non-cardiovascular drugs that inhibit hERG-encoded potassium channels and risk of sudden cardiac death. Heart 2011, 97, 215–220. [Google Scholar] [CrossRef]
- Schmidt, N.J.; Lennette, E.H.; Ho, H.H. An Apparently New Enterovirus Isolated from Patients with Disease of the Central Nervous System. J. Infect. Dis. 1974, 129, 304–309. [Google Scholar] [CrossRef]
- Barberis, A.; Deiana, M.; Spissu, Y.; Azara, E.; Fadda, A.; Serra, P.A.; D’hallewin, G.; Pisano, M.; Serreli, G.; Orrù, G.; et al. Antioxidant, antimicrobial, and other biological properties of pompia juice. Molecules 2020, 25, 3186. [Google Scholar] [CrossRef]
- Han, B.X.; Cong, H. Enterovirus 71 induces apoptosis by directly modulating the conformational activation of pro-apoptotic protein. J. Gen. Virol. 2017, 98, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Du, N.; Cong, H.; Tian, H.; Zhang, H.; Zhang, W.; Song, L.; Tien, P. Cell Surface Vimentin Is an Attachment Receptor for Enterovirus 71. J. Virol. 2014, 88, 5816–5833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doitsh, G.; Galloway, N.L.K.; Geng, X.; Yang, Z.; Monroe, K.M.; Zepeda, O.; Hunt, P.W.; Hatano, H.; Sowinski, S.; Muñoz-Arias, I.; et al. Cell death by pyroptosis drives CD4 T-cell depletion in HIV-1 infection. Nature 2014, 505, 509–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kofahi, H.M.; Taylor, N.G.A.; Hirasawa, K.; Grant, M.D.; Russell, R.S. Hepatitis C Virus Infection of Cultured Human Hepatoma Cells Causes Apoptosis and Pyroptosis in Both Infected and Bystander Cells. Sci. Rep. 2016, 6, 37433. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.Y.; Chu, J.J.H. Dengue virus-infected human monocytes trigger late activation of caspase-1, which mediates pro-inflammatory IL-1β secretion and pyroptosis. J. Gen. Virol. 2013, 94, 2215–2220. [Google Scholar] [CrossRef]
- Yogarajah, T.; Ong, K.C.; Perera, D.; Wong, K.T. AIM2 Inflammasome-Mediated Pyroptosis in Enterovirus A71-Infected Neuronal Cells Restricts Viral Replication. Sci. Rep. 2017, 7, 1–16. [Google Scholar]
- Wang, H.; Lei, X.; Xiao, X.; Yang, C.; Lu, W.; Huang, Z.; Leng, Q.; Jin, Q.; He, B.; Meng, G.; et al. Reciprocal Regulation between Enterovirus 71 and the NLRP3 Inflammasome. Cell Rep. 2015, 12, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Qin, Y.; Wang, T.; Chen, Y.; Lang, X.; Zheng, J.; Gao, S.; Chen, S.; Zhong, X.; Mu, Y.; et al. Pyroptosis induced by enterovirus 71 and coxsackievirus B3 infection affects viral replication and host response. Sci. Rep. 2018, 8, 2887. [Google Scholar] [CrossRef]
- Zhu, X.; Wu, T.; Chi, Y.; Ge, Y.; Wu, B.; Zhou, M.; Zhu, F.; Ji, M.; Cui, L. Pyroptosis induced by enterovirus A71 infection in cultured human neuroblastoma cells. Virology 2018, 521, 69–76. [Google Scholar] [CrossRef]
- Nagata, N.; Shimizu, H.; Ami, Y.; Tano, Y.; Harashima, A.; Suzaki, Y.; Sato, Y.; Miyamura, T.; Sata, T.; Iwasaki, T. Pyramidal and extrapyramidal involvement in experimental infection of cynomolgus monkeys with enterovirus 71. J. Med. Virol. 2002, 67, 207–216. [Google Scholar] [CrossRef]
- Zhang, Y.; Cui, W.; Liu, L.; Wang, J.; Zhao, H.; Liao, Y.; Na, R.; Dong, C.; Wang, L.; Xie, Z.; et al. Pathogenesis study of enterovirus 71 infection in rhesus monkeys. Lab. Investig. 2011, 91, 1337–1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kataoka, C.; Suzuki, T.; Kotani, O.; Iwata-Yoshikawa, N.; Nagata, N.; Ami, Y.; Wakita, T.; Nishimura, Y.; Shimizu, H. The Role of VP1 Amino Acid Residue 145 of Enterovirus 71 in Viral Fitness and Pathogenesis in a Cynomolgus Monkey Model. PLoS Path. 2015, 11, e1005033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Chou, C.; Lei, H.; Liu, C.; Wang, S.; Yan, J.; Su, I.; Wang, J.; Yeh, T.; Chen, S.; et al. A mouse muscle-adapted enterovirus 71 strain with increased virulence in mice. Microbes Infect. 2011, 13, 862–870. [Google Scholar] [CrossRef]
- Wang, Y.F.; Liu, C.; Wang, S.; Yan, J. A mouse-adapted enterovirus 71 strain causes neurological disease in mice after oral infection. J. Virol. 2004, 78, 7916–7924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.C. A murine oral enterovirus 71 infection model with central nervous system involvement. J. Gen. Virol. 2004, 85, 69–77. [Google Scholar] [CrossRef]
- Chua, B.H.; Phuektes, P.; Sanders, S.A.; Nicholls, P.K.; McMinn, P.C. The molecular basis of mouse adaptation by human enterovirus 71. J. Gen. Virol. 2008, 89, 1622–1632. [Google Scholar] [CrossRef]
- Yao, P.P.; Qian, L.; Xia, Y.; Xu, F.; Yang, Z.N.; Xie, R.H.; Li, X.; Liang, W.F.; Huang, X.X.; Zhu, Z.Y.; et al. Enterovirus 71-induced neurological disorders in young gerbils, Merionesunguiculatus: Development and application of a neurological disease model. PLoS ONE 2012, 7, e51996. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-F.; Yu, C.-K. Animal models of enterovirus 71 infection: Applications and limitations. J. Biomed. Sci. 2014, 21, 31. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cmp | Vero-76 | EV-A71 | CV-B3 | CV-B4 | CV-B5 | PV-1 | HeLa | EV-D68 | LLC-MK2 | Echo9 |
| a CC50 | b EC50 | c EC50 | c EC50 | c EC50 | c EC50 | d CC50 | c EC50 | e CC50 | c EC50 | |
| 1a | 60/24 * | >24 | >60 | >60 | >60 | 11.5 | 9 | >9 | 6 | >6 |
| 1b | 46/40 * | >40 | >46 | >46 | 7 | 9 | 9 | >9 | 6.5 | >6.5 |
| 2a | >100 | 7 | 17 | ≥100 | 10 | 9 | 100 | >100 | >100 | 8 |
| 2b | >100 | 3.5 | 33 | 25.5 | 10 | 6 | 100 | >100 | 12 | 4 |
| 3a | 25.5/8 * | >8 | >25.5 | >25.5 | >25.5 | >25.5 | 8 | >8 | 7 | >7 |
| 3b | 9 | >9 | >9 | >9 | 2 | - | 6.4 | >6.4 | 12 | >12 |
| 4a | >100 | >100 | >100 | >100 | >100 | ≥100 | >100 | >100 | 29.5 | >29.5 |
| 4b | >100 | 70 | >100 | >100 | >100 | 20 | >100 | >100 | >100 | >100 |
| 5a | >100 | 31 | >100 | >100 | >100 | 12 | >100 | >100 | 100 | >100 |
| 5b | >100 | 6 | >100 | >100 | >100 | 10 | >100 | 4 | >100 | >100 |
| Plc | >100 | 0.005 ± 0.001 | 2 ± 0.005 | 2 ± 0.5 | 0.3 | >100 | 0.1 | |||
| NM 107 | >100 | 29 | 6 | |||||||
| Rup | >100 | 0.07 |
| Compounds | BHK-21 | MDBK | Vero-76 | HeLa | LLC-MK2 |
|---|---|---|---|---|---|
| a CC50 | b CC50 | c CC50 | d CC50 | e CC50 | |
| 1a | 36 | 65 | 60/24 * | 9 | 6 |
| 1b | 20 | 9.0 | 46/40 * | 9 | 6.5 |
| 2a | >100 | 80 | >100 | 100 | >100 |
| 2b | >100 | >100 | >100 | 100 | 12 |
| 3a | 8.0 | 2.5 | 25.5/8 * | 8 | 7 |
| 3b | 9 | 7 | 9 | 6.4 | 12 |
| 4a | >100 | >100 | >100 | >100 | 29.5 |
| 4b | >100 | >100 | >100 | >100 | >100 |
| 5a | >100 | >100 | >100 | >100 | 100 |
| 5b | >100 | >100 | >100 | >100 | >100 |
| ADME Profile | Toxicity a | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Samples | PPB (%) a | BBB Permeability | Lipophilicity b | Water sol b | GI abs b | Caco-2 cells a | P-gp sub b | hERG Inhibition | Ames Test | ||||||
| (Cbrain/Cblood) a | Pr. a | Pr. b | Consensus | Solubility | (nm/sec) | Pr. | TA100 | TA1535 | |||||||
| Log Po/w | Class | S9 act | No S9 act | S9 act | No S9 act | ||||||||||
| 1a | 92 | 11.95 | High | Yes | 3.88 | Soluble (m) | High | 36.9 | Yes | Medium | M | Pos | Neg | Neg | Pos |
| 1b | 95 | 12.61 | High | Yes | 4.14 | Soluble (m) | High | 42.3 | No | Medium | M | Pos | Neg | Neg | Neg |
| 2a | 90 | 13.29 | High | Yes | 4.41 | Soluble (m) | High | 53.2 | No | Medium | M | Pos | Neg | Neg | Pos |
| 2b | 93 | 13.74 | High | Yes | 4.68 | Soluble (m) | High | 54.8 | No | Medium | NM | Neg | Neg | Neg | Neg |
| 3a | 92 | 14.56 | High | Yes | 4.93 | Soluble (m) | High | 49.2 | No | Medium | M | Neg | Neg | Neg | Pos |
| 3b | 94 | 14.87 | High | Yes | 5.19 | Soluble (m) | High | 52.9 | No | Medium | NM | Neg | Neg | Neg | Neg |
| 4a | 92 | 12.24 | High | Yes | 4.19 | Soluble (m) | High | 49.9 | Yes | Medium | M | Pos | Neg | Neg | Neg |
| 4b | 95 | 12.74 | High | Yes | 4.45 | Soluble (m) | High | 53.4 | No | Medium | M | Neg | Neg | Pos | Neg |
| 5a | 93 | 12.79 | High | Yes | 4.49 | Soluble (m) | High | 43.6 | Yes | Medium | M | Neg | Neg | Pos | Pos |
| 5b | 95 | 13.24 | High | Yes | 4.75 | Soluble (m) | High | 50.1 | No | Medium | M | Neg | Neg | Pos | Neg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibba, R.; Carta, A.; Madeddu, S.; Caria, P.; Serreli, G.; Piras, S.; Sestito, S.; Loddo, R.; Sanna, G. Inhibition of Enterovirus A71 by a Novel 2-Phenyl-Benzimidazole Derivative. Viruses 2021, 13, 58. https://0-doi-org.brum.beds.ac.uk/10.3390/v13010058
Ibba R, Carta A, Madeddu S, Caria P, Serreli G, Piras S, Sestito S, Loddo R, Sanna G. Inhibition of Enterovirus A71 by a Novel 2-Phenyl-Benzimidazole Derivative. Viruses. 2021; 13(1):58. https://0-doi-org.brum.beds.ac.uk/10.3390/v13010058
Chicago/Turabian StyleIbba, Roberta, Antonio Carta, Silvia Madeddu, Paola Caria, Gabriele Serreli, Sandra Piras, Simona Sestito, Roberta Loddo, and Giuseppina Sanna. 2021. "Inhibition of Enterovirus A71 by a Novel 2-Phenyl-Benzimidazole Derivative" Viruses 13, no. 1: 58. https://0-doi-org.brum.beds.ac.uk/10.3390/v13010058