
Gaps in Serologic Immunity against Contemporary Swine-Origin Influenza A Viruses among Healthy Individuals in the United States

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
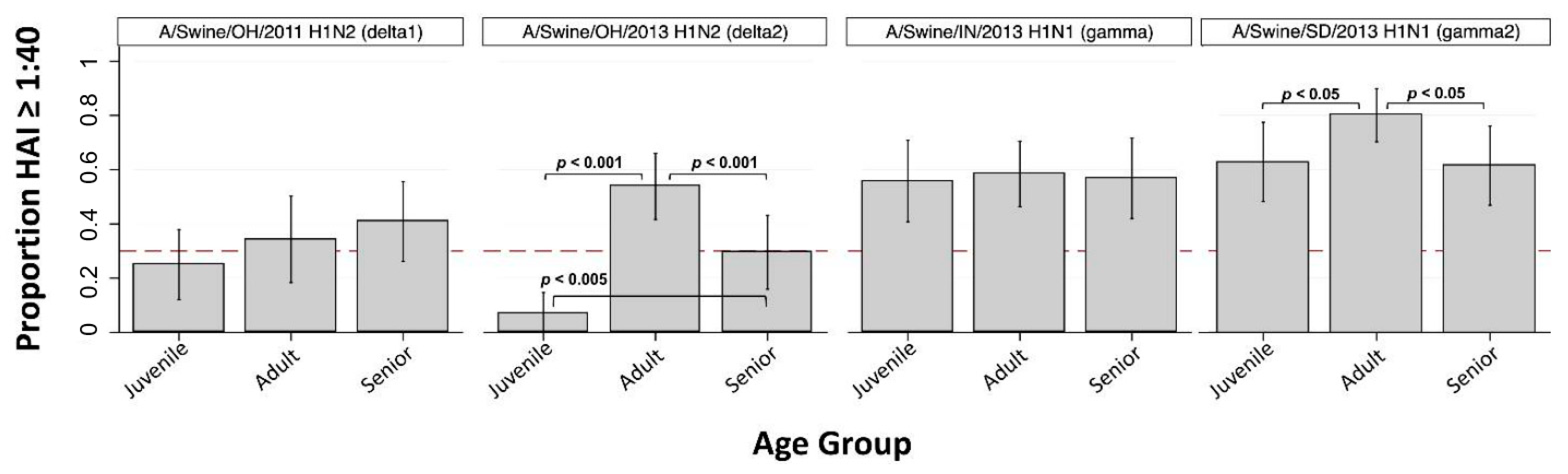
3.1. H1 Subtype IAV-S
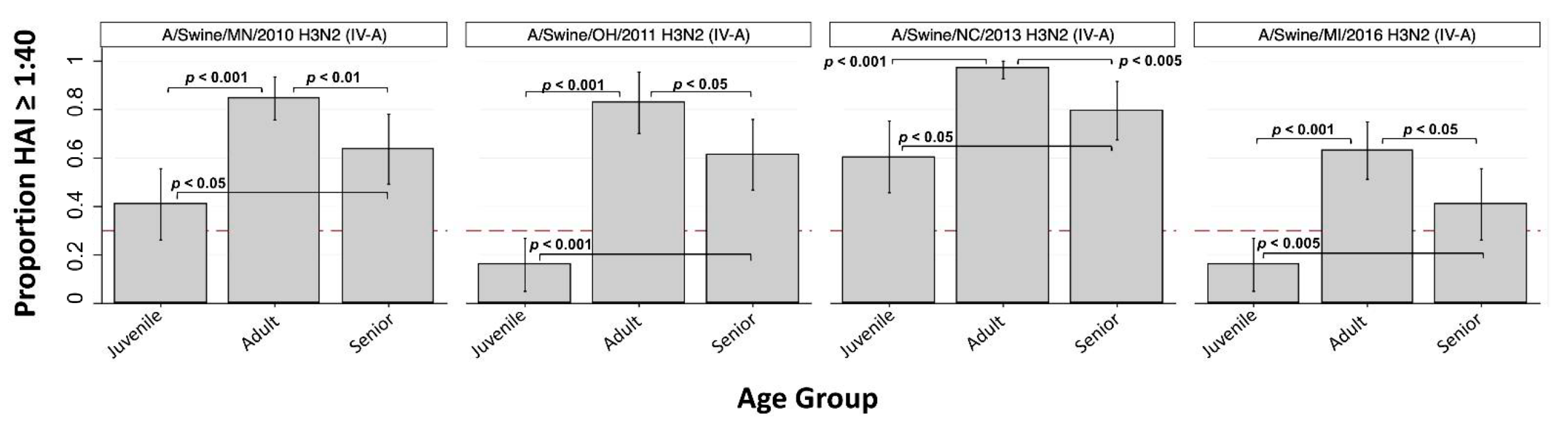
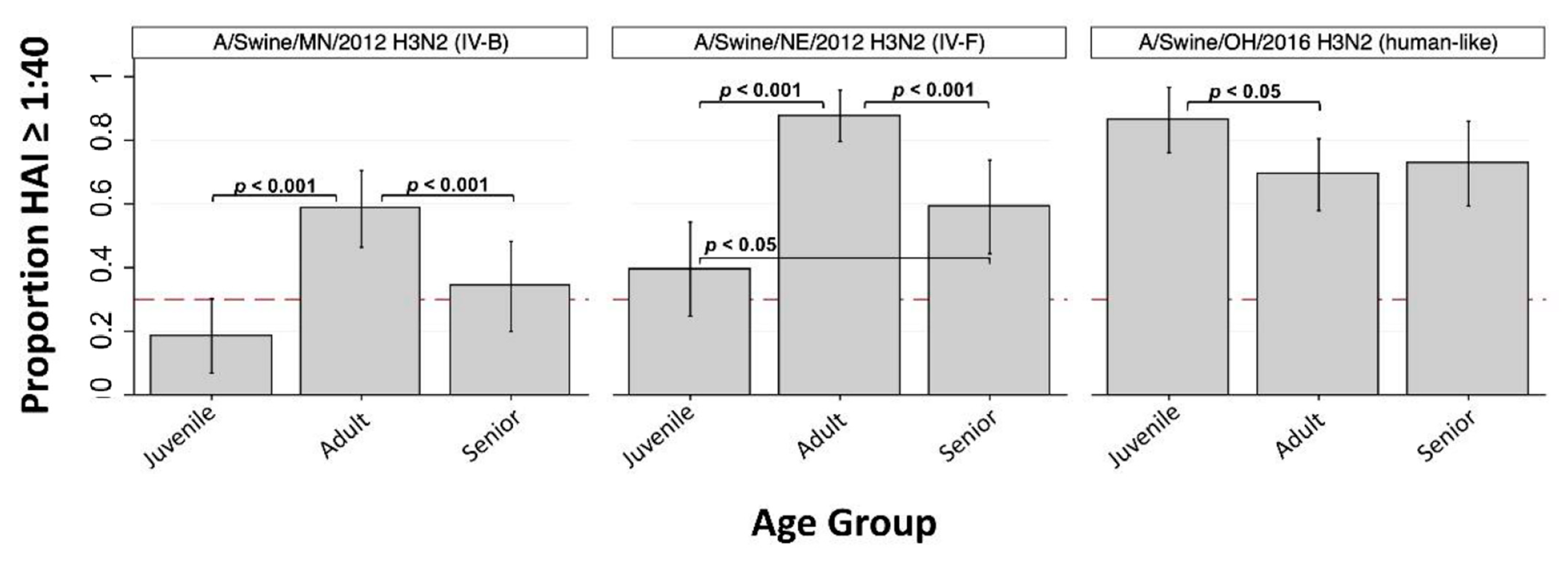
3.2. H3 Subtype IAV-S
3.3. Seroprotection within Decades of Birth
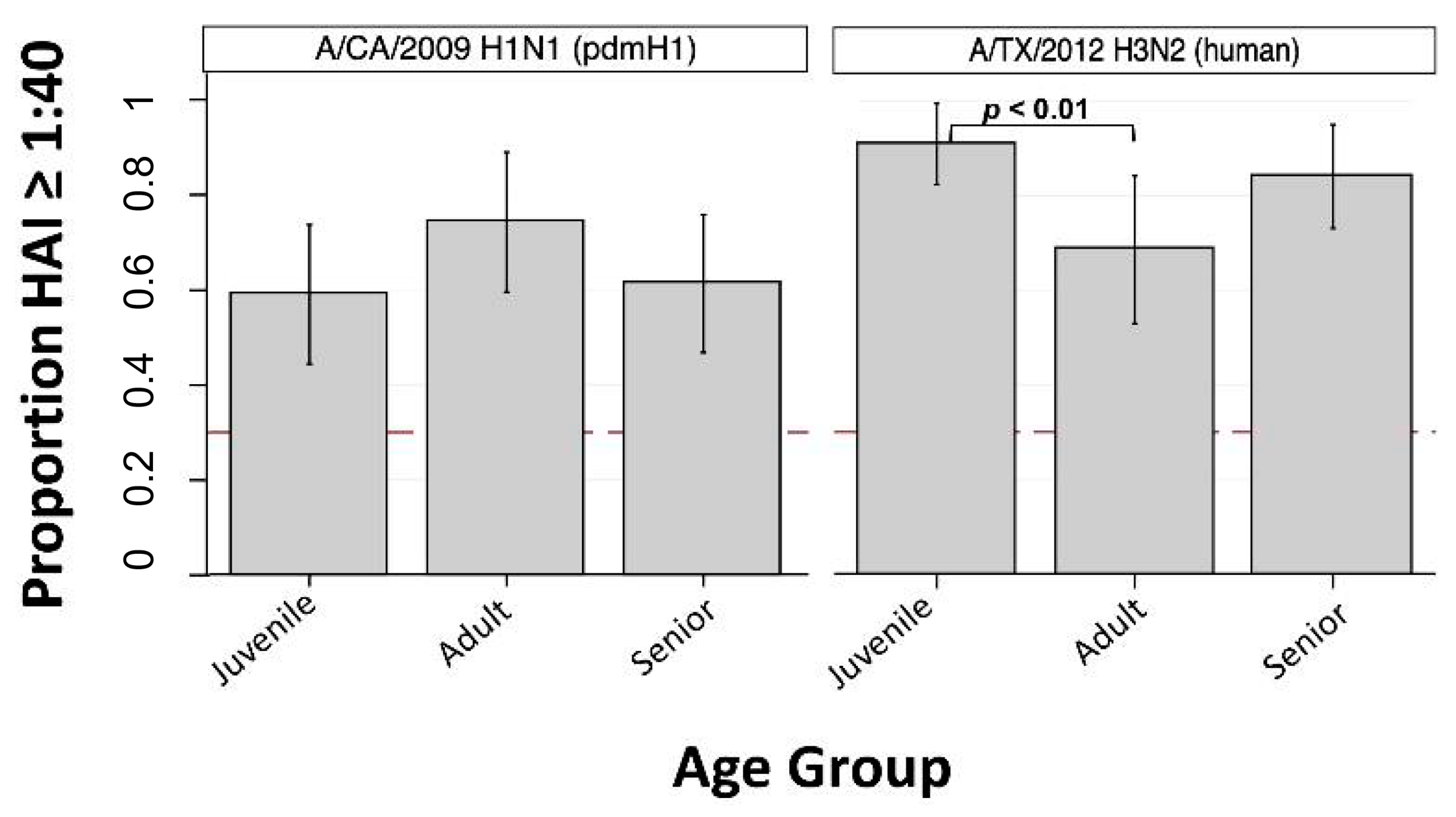
3.4. Human Seasonal IAVs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Horimoto, T.; Kawaoka, Y. Influenza: Lessons from past pandemics, warnings from current incidents. Nat. Rev. Microbiol. 2005, 3, 591–600. [Google Scholar] [CrossRef]
- Mena, I.; I Nelson, M.; Quezada-Monroy, F.; Dutta, J.; Cortes-Fernández, R.; Lara-Puente, J.H.; Castro-Peralta, F.; Cunha, L.F.; Trovão, N.S.; Lozano-Dubernard, B.; et al. Origins of the 2009 H1N1 influenza pandemic in swine in Mexico. eLife 2016, 5, 16777. [Google Scholar] [CrossRef]
- Kilbourne, E.D. Influenza Pandemics of the 20th Century. Emerg. Infect. Dis. 2006, 12, 9–14. [Google Scholar] [CrossRef]
- Smith, G.J.D.; Vijaykrishna, D.; Bahl, J.; Lycett, S.J.; Worobey, M.; Pybus, O.G.; Ma, S.K.; Cheung, C.L.; Raghwani, J.; Bhatt, S.; et al. Origins and evolutionary genomics of the 2009 swine-origin H1N1 influenza A epidemic. Nat. Cell Biol. 2009, 459, 1122–1125. [Google Scholar] [CrossRef] [Green Version]
- Dawood, F.S.; Iuliano, A.D.; Reed, C.; Meltzer, M.I.; Shay, D.K.; Cheng, P.Y.; Widdowson, M.A. Estimated global mortality associated with the first 12 months of 2009 pandemic in-fluenza A H1N1 virus circulation: A modelling study. Lancet Infect. Dis. 2012, 12, 687–695. [Google Scholar] [CrossRef] [Green Version]
- Van Reeth, K.; Vincent, A.L. Influenza Viruses. In Diseases of Swine, 11th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 576–593. [Google Scholar]
- Nelson, M.I.; Vincent, A.L. Reverse zoonosis of influenza to swine: New perspectives on the human–animal interface. Trends Microbiol. 2015, 23, 142–153. [Google Scholar] [CrossRef] [Green Version]
- Nelson, M.I.; Stratton, J.; Killian, M.L.; Janas-Martindale, A.; Vincent, A.L. Continual Reintroduction of Human Pandemic H1N1 Influenza A Viruses into Swine in the United States, 2009 to 2014. J. Virol. 2015, 89, 6218–6226. [Google Scholar] [CrossRef] [Green Version]
- Schicker, R.S.; Rossow, J.A.; Eckel, S.; Fisher, N.; Bidol, S.; Tatham, L.; Matthews-Greer, J.; Sohner, K.; Bowman, A.S.; Avrill, J.; et al. Outbreak of Influenza A(H3N2) Variant Virus Infections Among Persons Attending Agricultural Fairs Housing Infected Swine—Michigan and Ohio, July–August 2016. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 1157–1160. [Google Scholar] [CrossRef] [Green Version]
- Bowman, A.S.; Walia, R.R.; Nolting, J.M.; Vincent, A.L.; Killian, M.L.; Zentkovich, M.M.; Forshey, T. Influenza A(H3N2) Virus in Swine at Agricultural Fairs and Transmission to Hu-mans, Michigan and Ohio, USA, 2016. Emerg. Infect. Dis. 2017, 23, 1551–1555. [Google Scholar] [CrossRef]
- Rajão, D.S.; Gauger, P.C.; Anderson, T.K.; Lewis, N.S.; Abente, E.J.; Killian, M.L.; Perez, D.R.; Sutton, T.C.; Zhang, J.; Vincent, A. Novel Reassortant Human-Like H3N2 and H3N1 Influenza A Viruses Detected in Pigs Are Virulent and Antigenically Distinct from Swine Viruses Endemic to the United States. J. Virol. 2015, 89, 11213–11222. [Google Scholar] [CrossRef] [Green Version]
- Rajao, D.; Walia, R.; Campbell, B.; Gauger, P.C.; Janas-Martindale, A.; Killian, M.L.; Vincent, A. Reassortment between Swine H3N2 and 2009 Pandemic H1N1 in the United States Resulted in Influenza A Viruses with Diverse Genetic Constellations with Variable Virulence in Pigs. J. Virol. 2016, 91, e01763-16. [Google Scholar] [CrossRef] [Green Version]
- Rajao, D.S.; Anderson, T.K.; Kitikoon, P.; Stratton, J.; Lewis, N.S.; Vincent, A. Antigenic and genetic evolution of contemporary swine H1 influenza viruses in the United States. Virology 2018, 518, 45–54. [Google Scholar] [CrossRef]
- De Jong, J.C.; Van Nieuwstadt, A.P.; Kimman, T.G.; Loeffen, W.L.A.; Bestebroer, T.M.; Bijlsma, K.; Claas, E.C.J. Antigenic drift in swine influenza H3 haemagglutinins with implica-tions for vaccination policy. Vaccine 1999, 17, 1321–1328. [Google Scholar] [CrossRef]
- Anderson, T.K.; Campbell, B.A.; Nelson, M.I.; Lewis, N.S.; Janas-Martindale, A.; Killian, M.L.; Vincent, A.L. Characterization of co-circulating swine influenza A viruses in North Amer-ica and the identification of a novel H1 genetic clade with antigenic significance. Virus Res. 2015, 201, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Lorusso, A.; Vincent, A.; Harland, M.L.; Alt, D.; Bayles, D.O.; Swenson, S.L.; Gramer, M.R.; Russell, C.A.; Smith, D.J.; Lager, K.M.; et al. Genetic and antigenic characterization of H1 influenza viruses from United States swine from 2008. J. Gen. Virol. 2010, 92, 919–930. [Google Scholar] [CrossRef]
- Nelson, M.I.; Stucker, K.M.; Schobel, S.A.; Trovão, N.S.; Das, S.R.; Dugan, V.G.; Nelson, S.W.; Sreevatsan, S.; Killian, M.L.; Nolting, J.M.; et al. Introduction, Evolution, and Dissemination of Influenza A Viruses in Exhibition Swine in the United States during 2009 to 2013. J. Virol. 2016, 90, 10963–10971. [Google Scholar] [CrossRef] [Green Version]
- Nelson, M.I.; Wentworth, E.D.; Das, S.R.; Sreevatsan, S.; Killian, M.L.; Nolting, J.M.; Slemons, R.D.; Bowman, A.S. Evolutionary Dynamics of Influenza A Viruses in US Exhibition Swine. J. Infect. Dis. 2015, 213, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Webby, R.J.; Rossow, K.; Erickson, G.; Sims, Y.; Webster, R. Multiple lineages of antigenically and genetically diverse influenza A virus co-circulate in the United States swine population. Virus Res. 2004, 103, 67–73. [Google Scholar] [CrossRef]
- Joseph, U.; Su, Y.C.F.; Vijaykrishna, D.; Smith, G.J.D. The ecology and adaptive evolution of influenza A interspecies transmission. Influ. Other Respir. Viruses 2016, 11, 74–84. [Google Scholar] [CrossRef]
- Wong, K.K.; Greenbaum, A.; Moll, M.E.; Lando, J.; Moore, E.L.; Ganatra, R.; Biggerstaff, M.; Lam, E.; Smith, E.E.; Storms, A.D.; et al. Outbreak of Influenza A (H3N2) Variant Virus Infection among Attendees of an Agricultural Fair, Pennsylvania, USA, 2011. Emerg. Infect. Dis. 2012, 18, 1937–1944. [Google Scholar] [CrossRef]
- Jhung, M.A.; Epperson, S.; Biggerstaff, M.; Allen, D.; Balish, A.; Barnes, N.; Beaudoin, A.; Berman, L.; Bidol, S.; Blanton, L.; et al. Outbreak of Variant Influenza A(H3N2) Virus in the United States. Clin. Infect. Dis. 2013, 57, 1703–1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, A.S.; Nelson, S.W.; Page, S.L.; Nolting, J.M.; Killian, M.L.; Sreevatsan, S.; Slemons, R.D. Swine-to-Human Transmission of Influenza A(H3N2) Virus at Agricultural Fairs, Ohio, USA, 2012. Emerg. Infect. Dis. 2014, 20, 1472–1480. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Tool for Influenza Pandemic Risk Assessment (TIPRA); WHO Press: Geneva, Switzerland, 2016; Volume 1. [Google Scholar]
- Cox, N.; Trock, S.C.; Burke, S.A. Pandemic Preparedness and the Influenza Risk Assessment Tool (IRAT). Curr. Top. Microbiol. Immunol. 2014, 385, 119–136. [Google Scholar] [CrossRef] [PubMed]
- Nachbagauer, R.; Choi, A.; Hirsh, A.; Margine, I.; Iida, S.; Barrera, A.; Ferres, M.; Albrecht, R.A.; García-Sastre, A.; Bouvier, N.M.; et al. Defining the antibody cross-reactome directed against the influenza virus surface glycoproteins. Nat. Immunol. 2017, 18, 464–473. [Google Scholar] [CrossRef]
- Skowronski, D.M.; Janjua, N.Z.; De Serres, G.; Purych, D.; Gilca, V.; Scheifele, D.W.; Dionne, M.; Sabaiduc, S.; Gardy, J.L.; Li, G.; et al. Cross-reactive and Vaccine-Induced Antibody to an Emerging Swine-Origin Variant of Influenza A Virus Subtype H3N2 (H3N2v). J. Infect. Dis. 2012, 206, 1852–1861. [Google Scholar] [CrossRef] [Green Version]
- Epperson, S.; Jhung, M.; Richards, S.; Quinlisk, P.; Ball, L.; Moll, M.; Boulton, R.; Haddy, L.; Biggerstaff, M.; Brammer, L.; et al. Human Infections With Influenza A(H3N2) Variant Virus in the United States, 2011–2012. Clin. Infect. Dis. 2013, 57, S4–S11. [Google Scholar] [CrossRef] [Green Version]
- Kitikoon, P.; Nelson, M.I.; Killian, M.L.; Anderson, T.K.; Koster, L.; Culhane, M.R.; Vincent, A.L. Genotype patterns of contemporary reassorted H3N2 virus in US swine. J. Gen. Virol. 2013, 94, 1236–1241. [Google Scholar] [CrossRef]
- Anderson, T.K.; Macken, C.A.; Lewis, N.S.; Scheuermann, R.H.; Van Reeth, K.; Brown, I.H.; Swenson, S.L.; Simon, G.; Saito, T.; Berhane, Y.; et al. A Phylogeny-Based Global Nomenclature System and Automated Annotation Tool for H1 Hemagglutinin Genes from Swine Influenza A Viruses. mSphere 2016, 1, e00275-16. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Anderson, T.K.; Zeller, M.A.; Gauger, P.C.; Vincent, A. octoFLU: Automated Classification for the Evolutionary Origin of Influenza A Virus Gene Sequences Detected in U.S. Swine. Microbiol. Resour. Announc. 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Aevermann, B.D.; Anderson, T.K.; Burke, D.F.; Dauphin, G.; Gu, Z.; Scheuermann, R.H. Influenza Research Database: An integrated bioinformatics resource for in-fluenza virus research. Nucleic Acids Res. 2016, 45, D466–D474. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Katz, J.M.; Hancock, K.; Xu, X. Serologic assays for influenza surveillance, diagnosis and vaccine evaluation. Expert Rev. Anti-Infect. Ther. 2011, 9, 669–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yen, H.-L.; Webster, R.G. Pandemic Influenza as a Current Threat. Curr. Top. Microbiol. Immunol. 2009, 333, 3–24. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.; Hoschler, K.; Hardelid, P.; Stanford, E.; Andrews, N.; Zambon, M. Incidence of 2009 pandemic influenza A H1N1 infec-tion in England: A cross-sectional serological study. Lancet 2010, 375, 1100–1108. [Google Scholar] [CrossRef]
- Annual Estimates of the Resident Population by Single Year of Age and Sex for the United States: 1 April 2010 to 1 July 2018; United States Census Bureau: Washington, DC, USA, 2018. Available online: factfinder.census.gov (accessed on 26 March 2020).
- Verma, N.; Dimitrova, M.; Carter, N.M.; Crevar, C.J.; Ross, T.M.; Golding, H.; Khurana, S. Influenza Virus H1N1pdm09 Infections in the Young and Old: Evidence of Greater Antibody Diversity and Affinity for the Hemagglutinin Globular Head Domain (HA1 Domain) in the Elderly than in Young Adults and Children. J. Virol. 2012, 86, 5515–5522. [Google Scholar] [CrossRef] [Green Version]
- Lewis, N. (Nicola); A Russell, C.; Langat, P.; Anderson, T.K.; Berger, K.; Bielejec, F.; Burke, D.; Dudas, G.; Fonville, J.M.; Fouchier, R.A.; et al. Author response: The global antigenic diversity of swine influenza A viruses. Author Response 2016, 5, e12217. [Google Scholar] [CrossRef]
- Webster, R.G. Original antigenic sin in ferrets: The response to sequential infections with influenza viruses. J. Immunol. 1966, 97, 177–183. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. Samples | ||
|---|---|---|
| Birth year | 1934–1943 | 10 |
| 1944–1953 | 15 | |
| 1954–1963 | 22 | |
| 1964–1973 | 14 | |
| 1974–1983 | 14 | |
| 1984–1993 | 34 | |
| 1994–2003 | 25 | |
| 2004–2013 | 19 | |
| Age group a | Juvenile b | 44 |
| Adult c | 65 | |
| Senior d | 44 | |
| Overall | 153 |
| HAI ≥ 1:40 [Proportion (95% CI)] | |||||
|---|---|---|---|---|---|
| Virus | HA Subclade | Juvenile c | Adult d | Senior e | Overall f |
| A/Swine/OH/2011 (H1N2) | delta1 | 25.0% ** (12.1, 37.9) | 34.3% ** (18.3, 50.2) | 40.9% (26.2, 55.6) | 32.2% |
| A/Swine/OH/2013 (H1N2) | delta2 | 7.0% * (0, 14.7) | 53.8% (41.6, 66.1) | 29.5% (15.9, 43.2) | 33.3% |
| A/Swine/IN/2013 (H1N1) | gamma | 55.8% (40.8, 70.8) | 58.5% (46.4, 70.5) | 56.8% (42.0, 71.6) | 53.5% |
| A/Swine/SD/2013 (H1N1) | gamma2 | 62.8% (48.2, 77.4) | 80.0% (70.2, 89.8) | 61.4% (46.8, 75.9) | 65.4% |
| A/CA/2009 (H1N1) | pdmH1 | 59.1% (44.4, 73.8) | 74.3% (59.6, 89.0) | 61.4% (46.8, 75.9) | 62.2% |
| A/Swine/MN/2010 (H3N2) | C_IV-A | 40.9% * (26.2, 55.6) | 84.6% (75.8, 93.5) | 63.6% (49.3, 78.0) | 63.7% |
| A/Swine/OH/2011 (H3N2) | C_IV-A | 15.9% * (5.0, 26.8) | 82.9% (70.2, 95.5) | 61.4% (46.8, 75.9) | 57.2% |
| A/Swine/NC/2013 (H3N2) | C_IV-A | 60.5% (45.7, 75.3) | 96.9% (92.7, 100) | 79.5% (67.5, 91.6) | 77.7% |
| A/Swine/MI/2016 (H3N2) | C_IV-A | 15.9% * (5.0, 26.8) | 63.1% (51.3, 74.9) | 40.9% (26.2, 55.6) | 42.5% |
| A/Swine/MN/2012 (H3N2) | C_IV-B | 18.6% * (6.8, 30.4) | 58.5% (46.4, 70.5) | 34.1% (19.9, 48.3) | 39.0% |
| A/Swine/NE/2012 (H3N2) | C_IV-F | 39.5% * (24.7, 54.3) | 87.7% (79.6, 95.7) | 59.1% (44.4, 73.8) | 63.3% |
| A/Swine/OH/2016 (H3N2) | human-like | 86.4% (76.1, 96.6) | 69.2% (57.9, 80.5) | 72.7% (59.4, 86.0) | 69.1% |
| A/TX/2012 (H3N2) | human | 90.9% (82.3, 99.5) | 68.6% (53.0, 84.2) | 84.1% (73.2, 95.0) | 73.3% |
| GMT [Mean (95% CI)] | |||||
|---|---|---|---|---|---|
| Virus | Subclade | Juvenile c | Adult d | Senior e | Overall |
| A/Swine/OH/2011 (H1N2) | delta1 | 13 (9, 18) | 17 (11, 27) | 24 (17, 34) | 18 (14, 22) |
| A/Swine/OH/2013 (H1N2) | delta2 | 8 (6, 10) | 31 (20, 48) | 14 (10, 21) | 17 (13, 21) |
| A/Swine/IN/2013 (H1N1) | gamma | 35 (20, 61) | 39 (25, 60) | 30 (20, 44) | 35 (27, 46) |
| A/Swine/SD/2013 (H1N1) | gamma2 | 49 (29, 85) | 91 (62, 134) | 34 (23, 51) | 58 (44, 75) |
| A/CA/2009 (H1N1) | pdmH1 | 47 (30, 74) | 78 (45, 137) | 36 (24, 54) | 49 (38, 65) |
| A/Swine/MN/2010 (H3N2) | C_IV-A | 19 (13, 29) | 136 (95, 197) | 29 (20, 41) | 50 (38, 64) |
| A/Swine/OH/2011 (H3N2) | C_IV-A | 11 (8, 15) | 110 (64, 187) | 39 (27, 56) | 33 (25, 44) |
| A/Swine/NC/2013 (H3N2) | C_IV-A | 46 (29, 72) | 267 (200, 357) | 94 (63, 139) | 120 (94, 152) |
| A/Swine/MI/2016 (H3N2) | C_IV-A | 9 (7, 12) | 48 (31, 73) | 25 (16, 37) | 24 (19, 31) |
| A/Swine/MN/2012 (H3N2) | C_IV-B | 10 (8, 14) | 30 (22, 42) | 18 (13, 24) | 19 (16, 23) |
| A/Swine/NE/2012 (H3N2) | C_IV-F | 19 (12, 31) | 163 (109, 244) | 42 (27, 66) | 60 (45, 80) |
| A/Swine/OH/2016 (H3N2) | human-like | 95 (62, 146) | 77 (48, 121) | 64 (43, 95) | 78 (60, 100) |
| A/TX/2012 (H3N2) | human | 179 (119, 269) | 71 (40, 127) | 83 (57, 120) | 104 (80, 136) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lorbach, J.N.; Fitzgerald, T.; Nolan, C.; Nolting, J.M.; Treanor, J.J.; Topham, D.J.; Bowman, A.S. Gaps in Serologic Immunity against Contemporary Swine-Origin Influenza A Viruses among Healthy Individuals in the United States. Viruses 2021, 13, 127. https://0-doi-org.brum.beds.ac.uk/10.3390/v13010127
Lorbach JN, Fitzgerald T, Nolan C, Nolting JM, Treanor JJ, Topham DJ, Bowman AS. Gaps in Serologic Immunity against Contemporary Swine-Origin Influenza A Viruses among Healthy Individuals in the United States. Viruses. 2021; 13(1):127. https://0-doi-org.brum.beds.ac.uk/10.3390/v13010127
Chicago/Turabian StyleLorbach, Joshua N., Theresa Fitzgerald, Carolyn Nolan, Jacqueline M. Nolting, John J. Treanor, David J. Topham, and Andrew S. Bowman. 2021. "Gaps in Serologic Immunity against Contemporary Swine-Origin Influenza A Viruses among Healthy Individuals in the United States" Viruses 13, no. 1: 127. https://0-doi-org.brum.beds.ac.uk/10.3390/v13010127