
In Vitro Evaluation of Anti-Rift Valley Fever Virus, Antioxidant and Anti-Inflammatory Activity of South African Medicinal Plant Extracts


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Viruses and Reagents
2.2. Plant Collection and Extraction
2.3. Cytotoxicity Assays
2.4. Antiviral Activity Assay
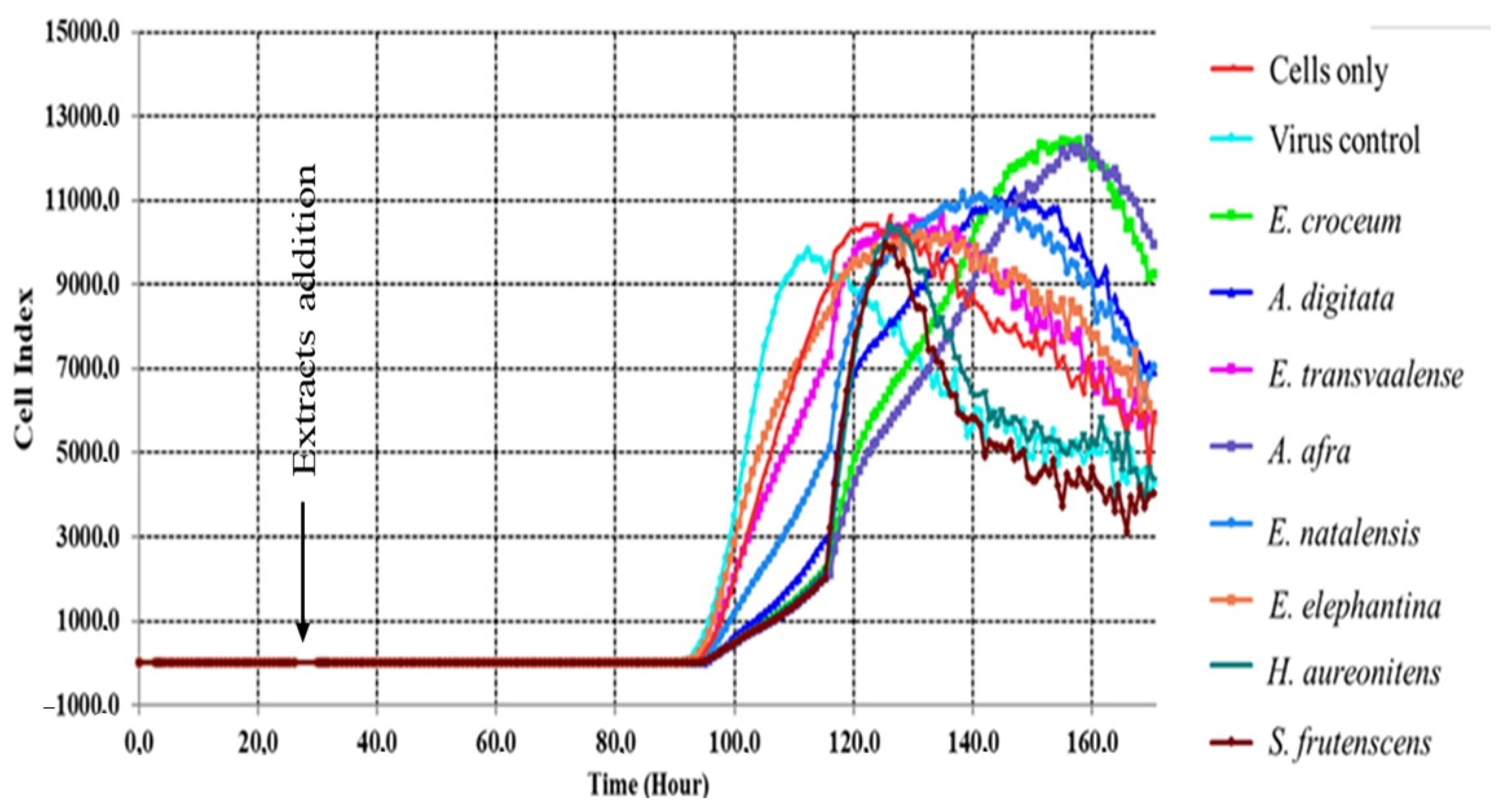
2.5. Real-Time Cell Analyzer (RTCA)
2.6. DPPH (2,2-Diphenyl-1-picrylhydrazyl) Radical-Scavenging Activity
2.7. ABTS+ (2,2′-Azino-bis-3-ethylbenzthiazoline-6-sulphonic acid) Radical Scavenging Assay
2.8. Lipopolysaccharides (LPS)-Induced Intracellular Reactive Oxygen/Nitrogen Species Measurement
2.9. Nitrite Concentrations as a Measure of Reactive Nitrogen Species
2.10. Statistical Analyses
3. Results
3.1. Cytotoxicity Results
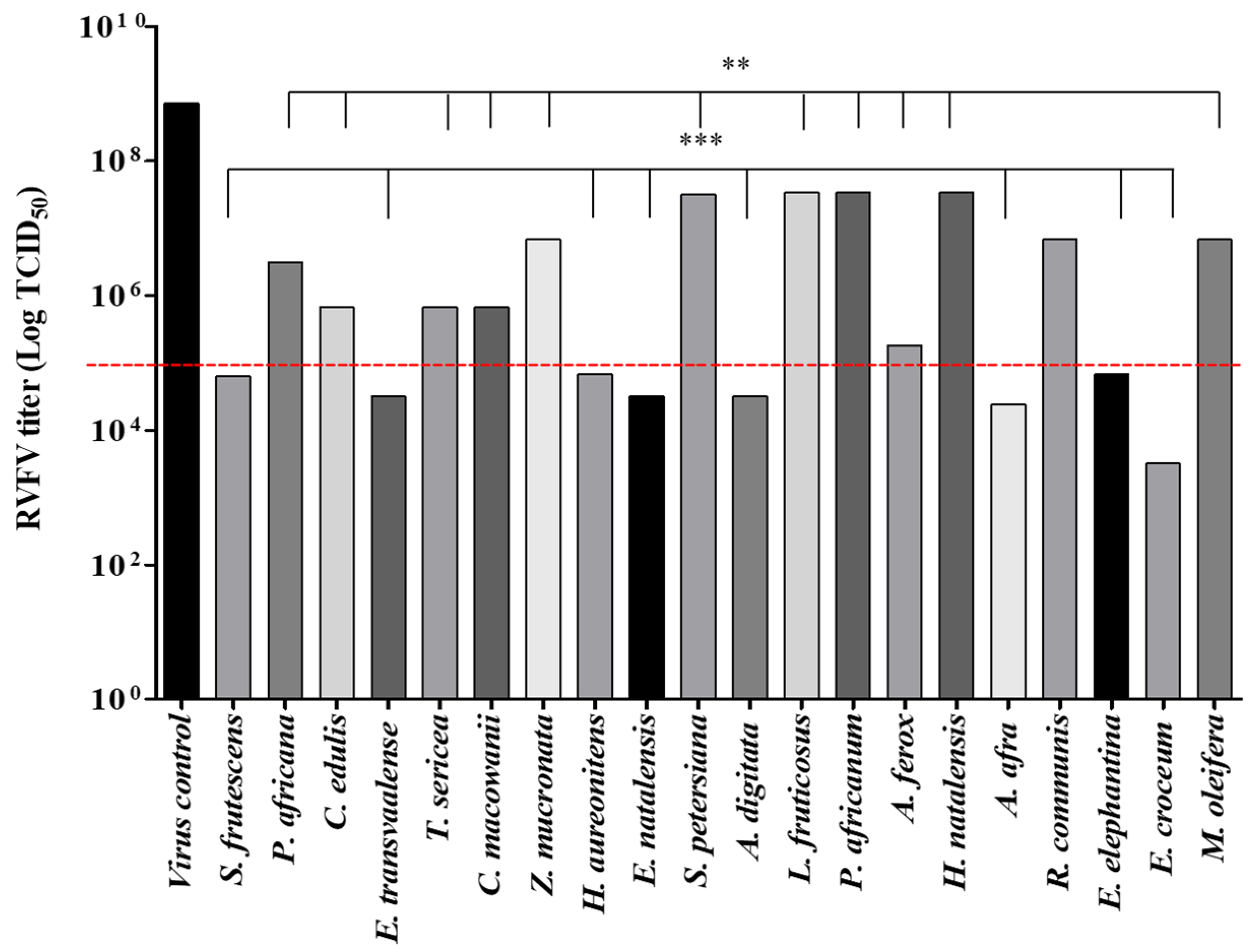
3.2. Antiviral Results
3.3. Real-Time Cell Analyzer (RTCA)
3.4. DPPH Radical-Scavenging Activity
3.5. ABTS+ Radical Scavenging Acvtiity
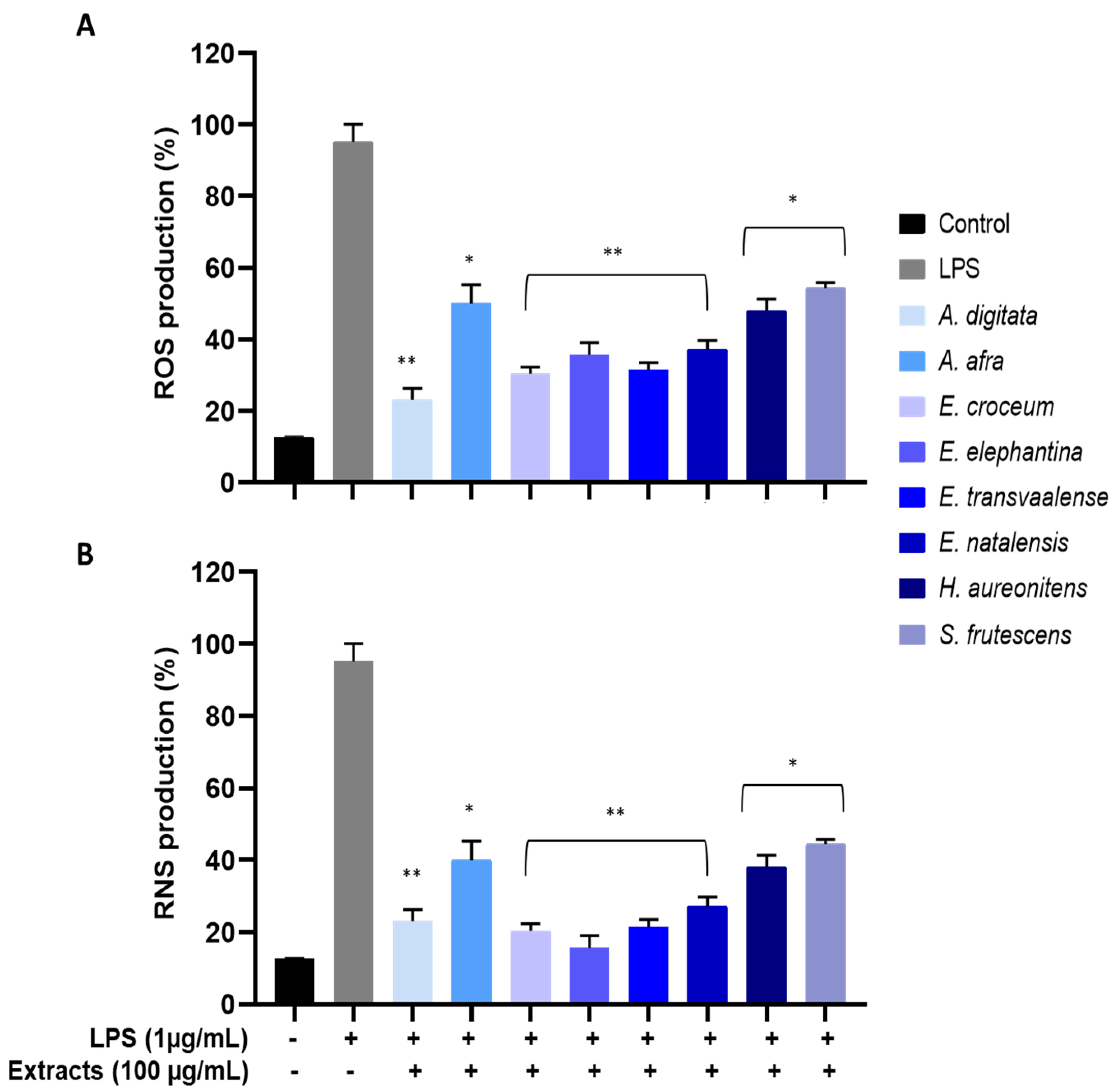
3.6. Measurement of LPS-Induced Intracellular ROS
3.7. Measurement of LPS-Induced Intracellular RNS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kalveram, B.; Lihoradova, O.; Indran, S.V.; Lokugamage, N.; Head, J.A.; Ikegami, T. Rift valley fever virus NSs inhibits host transcription independently of the degradation of dsRNA-dependent protein kinase PKR. Virology 2013, 435, 415–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fawzy, M.; Helmy, Y.A. The one health approach is necessary for the control of Rift Valley fever infections in Egypt. Viruses 2019, 11, 139. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.K.; Strand, M.; Saleeb, M.; Svensson, R.; Baranczewski, P.; Artursson, P.; Wadell, G.; Ahlm, C.; Elofsson, M.; Evander, M. Anti-Rift Valley fever virus activity in vitro, pre-clinical pharmacokinetics and oral bioavailability of benzavir-2, a broad-acting antiviral compound. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bird, B.H.; McElroy, A.K. Rift Valley fever virus: Unanswered questions. Antivir. Res. 2016, 132, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Rolin, A.I.; Berrang-ford, L.; Kulkarni, M.A.; Rolin, A.I.; Berrang-ford, L.; Kulkarni, M.A. The risk of Rift Valley fever virus introduction and establishment in the United States and European Union The risk of Rift Valley fever virus introduction and establishment in the United States and European Union. J. Microb. Infect. 2013, 2, 1–8. [Google Scholar]
- Liu, J.; Sun, Y.; Shi, W.; Tan, S.; Pan, Y.; Cui, S.; Zhang, Q.; Dou, X.; Lv, Y.; Li, X.; et al. The first imported case of Rift Valley fever in China reveals a genetic reassortment of different viral lineages. Emerg. Microb. Infect. 2017, 6, 4–7. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Rapid Risk Assessment Rift Valley Fever Outbreak in Mayotte, France Main Conclusions and Options for Response; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2019; pp. 1–12. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/RRA-Rift-Valley-fever-Mayotte-France-March-2019.pdf (accessed on 31 January 2021).
- Pereira-Dos-Santos, T.; Roiz, D.; Lourenço-De-Oliveira, R.; Paupy, C. A systematic review: Is Aedes albopictus an efficient bridge vector for zoonotic arboviruses? Pathogens 2020, 9, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization Regional Office for Africa. Weekly Bulletin on Outbreaks; WHO: Zeneva, Switzerland, 2020; pp. 1–20. [Google Scholar]
- Zhang, Z.; Rong, L.; Li, Y.P. Flaviviridae viruses and oxidative stress: Implications for viral pathogenesis. Oxid. Med. Cell. Longev. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- More, G.K.; Makola, R.T. In vitro analysis of free radical scavenging activities and suppression of LPS-induced ROS production in macrophage cells by Solanum sisymbriifolium extracts. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Huang, W.E.N.J.; Zhang, X.I.A.; Chen, W.E.I. Role of oxidative stress in Alzheimer’s disease (Review). Biomed. Rep. 2016, 4, 519–522. [Google Scholar] [CrossRef] [Green Version]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS sources in physiological and pathological conditions. Oxid. Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef]
- Collin, F. Chemical basis of reactive oxygen species reactivity and involvement in neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20, 2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Villani, R.M.; Wang, H.; Simpson, M.J.; Roberts, M.S.; Tang, M. The role of cellular reactive oxygen species in cancer chemotherapy. J. Exp. Clin. Can. Res. 2018, 37, 2661–2710. [Google Scholar] [CrossRef] [PubMed]
- Lee, C. Therapeutic modulation of virus-induced oxidative stress via the Nrf2-dependent antioxidative pathway. Oxid. Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, A.; Amaya, M.; Voss, K.; Chung, M.; Benedict, A.; Sampey, G.; Kehn-Hall, K.; Luchini, A.; Liotta, L.; Bailey, C.; et al. Reactive oxygen species activate NFκB (p65) and p53 and induce apoptosis in RVFV infected liver cells. Virology 2014, 449, 270–286. [Google Scholar] [CrossRef] [Green Version]
- Davinelli, S.; Scapagnini, G.; Denaro, F.; Calabrese, V.; Zella, D. Altered expression pattern of Nrf2/HO-1 axis during accelerated-senescence in HIV-1 transgenic rat. Biogerontology 2014, 15, 449–461. [Google Scholar] [CrossRef]
- Cymerys, J.; Kowalczyk, A.; Mikołajewicz, K.; Słońska, A.; Krzyzowska, M. Nitric oxide influences HSV-1-induced neuroinflammation. Oxid. Med. Cell. Longev. 2019, 2019. [Google Scholar] [CrossRef]
- Ivanov, A.V.; Valuev-Elliston, V.T.; Ivanova, O.N.; Kochetkov, S.N.; Starodubova, E.S.; Bartosch, B.; Isaguliants, M.G. Oxidative stress during HIV Infection: Mechanisms and consequences. Oxid. Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Zablockienė, B.; Ambrozaitis, A.; Kačergius, T.; Gravenstein, S. Implication of nitric oxide in the pathogenesis of influenza virus infection. Biologija 2012, 58, 15–25. [Google Scholar] [CrossRef]
- Jansen van Vuren, P.; Tiemessen, C.T.; Paweska, J.T. Anti-nucleocapsid protein immune responses counteract pathogenic effects of Rift Valley fever virus infection in mice. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2011, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedoreyev, S.A.; Krylova, N.V.; Mishchenko, N.P.; Vasileva, E.A.; Pislyagin, E.A.; Iunikhina, O.V.; Lavrov, V.F.; Svitich, O.A.; Ebralidze, L.K.; Leonova, G.N. Antiviral and antioxidant properties of echinochrome A. Mar. Drugs 2018, 16, 509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, T.M.; Espina, V.; Lundberg, L.; Pinkham, C.; Brahms, A.; Carey, B.D.; Lin, S.C.; Dahal, B.; Woodson, C.; de la Fuente, C.; et al. Combination kinase inhibitor treatment suppresses Rift Valley fever virus replication. Viruses 2018, 10, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keck, F.; Amaya, M.; Kehn-Hall, K.; Roberts, B.; Bailey, C.; Narayanan, A. Characterizing the effect of Bortezomib on rift valley fever virus multiplication. Antivir. Res. 2015, 120, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Brahms, A.; Mudhasani, R.; Pinkham, C.; Kota, K.; Nasar, F.; Zamani, R.; Bavari, S.; Kehn-Hall, K. Sorafenib impedes Rift Valley Fever Virus egress by inhibiting Valosin-Containing protein function in the cellular secretory pathway. J. Virol. 2017, 91, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benedict, A.; Bansal, N.; Senina, S.; Hooper, I.; Lundberg, L.; Mcelroy, A. Repurposing FDA-approved drugs as therapeutics to treat Rift valley fever virus infection. Front Microbiol. 2015, 6, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ly, H.J.; Ikegami, T. Rift valley fever virus NSs protein functions and the similarity to other bunyavirus NSs proteins. Virol. J. 2016, 13, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Atkins, C.; Freiberg, A.N. Recent advances in the development of antiviral therapeutics for Rift valley fever virus infection. Future Virol. 2017, 12, 651–665. [Google Scholar] [CrossRef]
- Hills, C.; Musili, P.M.; Walter, N.O.; Hu, G.; Zhou, Y. Medicinal plants and their traditional uses in local communities around Cherangani Hills, Western Kenya. Plants 2020, 9, 1–16. [Google Scholar]
- Bagley, M.A. Toward an effective indigenous knowledge protection regime case study of South Africa. CIGI 2018, 207. [Google Scholar]
- Karimi, A.; Majlesi, M.; Rafieian-kopaei, M. Herbal versus synthetic drugs: Beliefs and facts. J. Nephropharmacol. 2015, 4, 27–30. [Google Scholar] [PubMed]
- Salehi, B.; Kumar, N.V.A.; Şener, B.; Sharifi-Rad, M.; Kılıç, M.; Mahady, G.B.; Vlaisavljevic, S.; Iriti, M.; Kobarfard, F.; Setzer, W.N.; et al. Medicinal plants used in the treatment of human immunodeficiency virus. Int. J. Mol. Sci. 2018, 19, 1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathore, B.; Ali Mahdi, A.; Paul, B.N.; Narayan, S.P.; Kumar, S.D. Indian herbal medicines: Possible potent therapeutic agents for rheumatoid arthritis. J. Clin. Biochem. Nutr. 2007, 41, 12–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulaiman, L.K.; Oladele, O.A.; Shittu, I.A.; Emikpe, B.O.; Oladokun, A.T.; Meseko, C.A. In-vivo evaluation of the antiviral activity of methanolic root-bark extract of the African Baobab (A. digitata Lin). Afr. J. Biotechnol. 2011, 10, 4256–4258. [Google Scholar]
- Silva, O.; Barbosa, S.; Diniz, A.; Valdeira, M.L.; Gomes, E. Plant extracts with antiviral activity against Herpes simplex virus type 1 and African swine fever virus. Int. J. Pharmacogn. 1997, 38, 12–16. [Google Scholar] [CrossRef]
- Sharma, A.; Rangari, V. HIV-1 reverse transcriptase and protease assay of methanolic extracts of A. digitata L. Int. J. Pharm. Pharm. Sci. 2016, 8, 124–127. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.Q.; Van der Kooy, F.; Verpoorte, R. Artemisia afra: A potential flagship for African medicinal plants? SA J. Bot. 2009, 75, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Ferox, L.O.E. and Somnifera, W. Anti-viral effects of aqueous extracts of Aloe ferox and Withania somnifera on herpes simplex virus type 1 in cell culture. S. Afr. J. Sci. 2007, 103, 9–10. [Google Scholar]
- Asres, K.; Bucar, F.; Kartnig, T.; Witvrouw, M.; Pannecouque, C.; De Clercq, E. Antiviral activity against human immunodeficiency virus type 1 (HIV-1) and type 2 (HIV-2) of ethnobotanically selected Ethiopian medicinal plants. Phyther. Res. 2001, 15, 62–69. [Google Scholar] [CrossRef]
- Mukhtar, M.; Arshad, M.; Ahmad, M.; Pomerantz, R.J.; Wigdahl, B.; Parveen, Z. Antiviral potentials of medicinal plants. Virus Res. 2008, 131, 111–120. [Google Scholar] [CrossRef]
- Tolo, F.M.; Rukunga, G.M.; Muli, F.W.; Njagi, E.N.M.; Njue, W.; Kumon, K.; Mungai, G.M.; Muthaura, C.N.; Muli, J.M.; Keter, L.K.; et al. Anti-viral activity of the extracts of a Kenyan medicinal plant Carissa edulis against herpes simplex virus. J. Ethnopharmacol. 2006, 104, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Robin, V.; Le Bosse, B.; Boustie, J.; Amoros, M. In vitro antiviral activity of thirty-six plants. Fitoterapia 2002, 73, 346–350. [Google Scholar]
- Klos, M.; van de Venter, M.; Milne, P.J.; Traore, H.N.D.; Oosthuizen, M.V. In vitro anti-HIV activity of five selected South African medicinal plant extracts. J. Ethnopharmacol. 2009, 124, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Prinsloo, G.; Meyer, J.J.M.; Hussein, A.A.; Munoz, E.; Sanchez, R. A cardiac glucoside with in vitro anti-HIV activity isolated from E. croceum. Nat. Prod. Res. 2010, 24, 1743–1766. [Google Scholar] [CrossRef]
- Tolo, F.; Rukunga, G.; Muli, F.; Ochora, J.; Eizuru, Y. In vitro anti-viral activity of aqueous extracts of Kenyan C. edulis, P. africana and Melia azedarach against human cytomegalovirus. Afr. J. Health Sci. 2007, 14, 143–148. [Google Scholar]
- Sigidi, M.T.; Traoré, A.N.; Boukandou, M.M.; Tshisikhawe, M.P.; Ntuli, S.S.; Potgieter, N. Anti-HIV, pro-inflammatory and cytotoxicity properties of selected Venda plants. Indian J. Tradit. Knowl. 2017, 16, 545–552. [Google Scholar]
- Lall, N.; Meyer, J.J.M.; Taylor, M.B. Anti-HSV-1 activity of E. natalensis. S. Afr. J. Bot. 2005, 71, 444–446. [Google Scholar] [CrossRef] [Green Version]
- Meyer, J.J.M.; Afolayan, A.J.; Taylor, M.B.; Erasmus, D. Antiviral activity of galangin isolated from the aerial parts of H. aureonitens. J. Ethnopharmacol. 1997, 56, 165–169. [Google Scholar] [CrossRef]
- Hurinanthan, V. Anti-HIV Activity of Selected South African Medicinal Plants. Ph.D. Thesis, Durban University of Technology, KwaZulu Natal, South Africa, 2013. [Google Scholar]
- Lunat, I. Traditional, Complementary and Alternative Medicine Use in HIV-Positive Patients Imran Lunat. Ph.D. Thesis, Nelson Mandela Metropolitan University (NMMU), Port Elizabeth, South Africa, 2011. [Google Scholar]
- Hafidh, R.R.; Abdulamir, A.S.; Jahanshiri, F.; Abas, F.; Abu Bakar, F.; Sekawi, Z. Asia is the mine of natural antiviral products for public health. Open Complement. Med. J. 2009, 1, 58–68. [Google Scholar]
- Lipipun, V.; Kurokawa, M.; Suttisri, R.; Taweechotipatr, P.; Pramyothin, P.; Hattori, M.; Shiraki, K. Efficacy of Thai medicinal plant extracts against herpes simplex virus type 1 infection in vitro and in vivo. Antivir. Res. 2003, 60, 175–180. [Google Scholar] [CrossRef]
- Ali, H.; König, G.M.; Khalid, S.A.; Wright, A.D.; Kaminsky, R. Evaluation of selected Sudanese medicinal plants for their in vitro activity against hemoflagellates, selected bacteria, HIV-1-RT and tyrosine kinase inhibitory, and for cytotoxicity. J. Ethnopharmacol. 2002, 83, 219–228. [Google Scholar] [CrossRef]
- Younus, I.; Siddiq, A.; Assad, T.; Badar, S.; Jameel, S.; Ashraf, M. Screening antiviral activity of Moringa oliefera L. leaves against foot and mouth disease virus. Glob. Vet. 2015, 15, 409–413. [Google Scholar]
- Feustel, S.; Ayón-Pérez, F.; Sandoval-Rodriguez, A.; Rodríguez-Echevarría, R.; Contreras-Salinas, H.; Armendáriz-Borunda, J.; Sánchez-Orozco, L.V. Protective effects of Moringa oleifera on HBV Genotypes C and H Transiently Transfected Huh7 Cells. J. Immunol. Res. 2017, 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tshikalange, T.E.; Meyer, J.J.M.; Lall, N.; Muñoz, E.; Sancho, R.; Van de Venter, M.; Oosthuizen, V. In vitro anti-HIV-1 properties of ethnobotanically selected South African plants used in the treatment of sexually transmitted diseases. J. Ethnopharmacol. 2008, 119, 478–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terasaki, K.; Murakami, S.; Lokugamage, K.G.; Makino, S. Mechanism of tripartite RNA genome packaging in Rift Valley fever virus. Proc. Natl. Acad. Sci. USA 2011, 108, 804–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bessong, P.O.; Obi, C.L.; Andréola, M.L.; Rojas, L.B.; Pouységu, L.; Igumbor, E.; Meyer, J.J.M.; Quideau, S.; Litvak, S. Evaluation of selected South African medicinal plants for inhibitory properties against human immunodeficiency virus type 1 reverse transcriptase and integrase. J. Ethnopharmacol. 2005, 99, 83–91. [Google Scholar] [CrossRef]
- Martín-Folgar, R.; Lorenzo, G.; Boshra, H.; Iglesias, J.; Mateos, F.; Borrego, B.; Brun, A. Development and characterization of monoclonal antibodies against Rift Valley fever virus nucleocapsid protein generated by DNA immunization. MAbs 2010, 2, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Bergren, N.A.; Patterson, E.I.; Blair, H.; Ellis, R.P.; Kading, R.C. Methods for successful inactivation of Rift Valley fever virus in infected mosquitoes. J. Virol. Methods 2020, 276, 113794. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Lubisi, B.A.; Ndouvhada, P.N.; Neiffer, D.; Penrith, M.L.; Sibanda, D.R.; Bastos, A.D.S. Evaluation of a virus neutralisation test for detection of rift valley fever antibodies in Suid Sera. Trop. Med. Infect. Dis. 2019, 4, 52. [Google Scholar] [CrossRef] [Green Version]
- Volsteedt, Y.; Apostolides, Z. Comparison of the antioxidant content of fruits, vegetables and teas measured as vitamin C equivalents. Toxicology 2001, 166, 63–69. [Google Scholar]
- Elisha, I.L.; Dzoyem, J.; Mcgaw, L.J.; Botha, F.S.; Eloff, J.N. Antioxidant activity and relationships with total phenolics and total flavonoids of nine South African plants used traditionally to treat arthritis. BMC Complement. Altern. Med. 2016, 16, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rock, L.; Brunswick, N. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 10, 4290–4302. [Google Scholar]
- Wu, W.Y.; Wu, Y.Y.; Huang, H.; He, C.; Li, W.Z.; Wang, H.L.; Chen, H.Q.; Yin, Y.Y. Biochanin A attenuates LPS-induced pro-inflammatory responses and inhibits the activation of the MAPK pathway in BV2 microglial cells. Int. J. Mol. Med. 2015, 35, 391–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, J.; Fiji, N.; Roy, D.; Andrew, D.M.G.; Shihabudeen, M.S.; Chattopadhyay, D.; Thirumurugan, K. A rapid method to assess reactive oxygen species in yeast using H2DCF-DA. Anal. Methods 2015, 7, 8572–8575. [Google Scholar] [CrossRef]
- Shen, C.F.; Guilbault, C.; Li, X.; Elahi, S.M.; Ansorge, S.; Kamen, A.; Gilbert, R. Development of suspension adapted Vero cell culture process technology for production of viral vaccines. Vaccine 2019, 37, 6996–7002. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Orr-Burks, N.; Karpilow, J.; Tripp, R.A. Development of improved vaccine cell lines against rotavirus. Sci. Data 2017, 4, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ianevski, A.; Yao, R.; Fenstad, M.H.; Biza, S.; Zusinaite, E.; Reisberg, T.; Lysvand, H.; Løseth, K.; Landsem, V.M.; Malmring, J.F.; et al. Potential antiviral options against SARS-CoV-2 Infection. Viruses 2020, 12, 642. [Google Scholar] [CrossRef] [PubMed]
- Yelani, T.; Hussein, A.A.; Meyer, J.J.M. Isolation and identification of poisonous triterpenoids from E. croceum. Nat. Prod. Res. 2010, 6419, 1418–1425. [Google Scholar] [CrossRef]
- Celestino, V.R.L.; Maranhão, H.M.L.; Carlos, F.; Vasconcelos, B.; Lima, C.R.; Medeiros, G.C.R. Acute toxicity and laxative activity of Aloe ferox resin. Rev. Bras. Farmacogn. 2013, 23, 279–283. [Google Scholar] [CrossRef] [Green Version]
- Petiot, E.; Fournier, F.; Gény, C.; Pinton, H.; Marc, A. Rapid screening of serum-free media for the growth of adherent vero cells by using a small-scale and non-invasive tool. Appl. Biochem. Biotechnol. 2010, 160, 1600–1615. [Google Scholar] [CrossRef]
- Wu, W.; Li, R.; Li, X.; He, J.; Jiang, S.; Liu, S.; & Yang, J. Quercetin as an antiviral agent inhibits influenza a virus (IAV) Entry. Viruses 2015, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bagla, V.P.; Mcgaw, L.J.; Eloff, J.N. The antiviral activity of six South African plants traditionally used against infections in ethnoveterinary medicine. Vet. Microbiol. 2012, 115, 198–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhanani, T.; Singh, R.; Kumar, S. Extraction optimization of gallic acid, (+)-catechin, procyanidin-B2, (−)-epicatechin, (−)-epigallocatechin gallate, and (−)-epicatechin gallate: Their simultaneous identification and quantification in Saraca asoca. J. Food. Drug. Anal. 2017, 25, 4–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irondi, E.A.; Akintunde, J.K.; Agboola, S.O.; Boligon, A.A.; Athayde, M.L. Blanching influences the phenolics composition, antioxidant activity, and inhibitory effect of A. digitata leaves extract on α-amylase, α-glucosidase, and aldose reductase. Food Sci. Nutr. 2017, 5, 233–242. [Google Scholar] [CrossRef] [PubMed]
- De Torre, M.P.; Cavero, R.Y.; Calvo, M.I. and Vizmanos, J.L.W. A simple and a reliable method to quantify antioxidant activity in vivo. Antioxidants 2019, 8, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilyasov, I.R.; Beloborodov, V.L.; Selivanova, I.A.; Terekhov, R.P. ABTS/PP decolorization assay of antioxidant capacity reaction pathways. Int. J. Mol. Sci. 2020, 21, 1131. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.C. The endotoxin hypothesis of neurodegeneration. J. Neuroinflammation. 2019, 16, 180. [Google Scholar] [CrossRef] [Green Version]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compos. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Maroyi, A. Medicinal uses, phytochemistry, pharmacology and toxicological properties of Elaeodendron croceum. Trop. J. Pharm. Res. 2019, 18, 669–676. [Google Scholar]
- Odeyemi, S.W.; Afolayan, A.J. Biological activities and phytochemical screening of E. croceum (Thunb.) DC leaves and stem barks extracts. Int. J. Phytomed. 2017, 9, 566–575. [Google Scholar] [CrossRef] [Green Version]
- Carrasco-Torres, G.; Baltiérrez-Hoyos, R.; Andrade-Jorge, E.; Villa-Treviño, S.; Trujillo-Ferrara, J.G.; Vásquez-Garzón, V.R. Cytotoxicity, oxidative stress, cell cycle arrest, and mitochondrial apoptosis after combined treatment of Hepatocarcinoma cells with Maleic Anhydride derivatives and quercetin. Oxid. Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Kesic, M.J.; Simmons, S.O.; Bauer, R.; Jaspers, I. Free radical biology & medicine Nrf2 expression modifies influenza A entry and replication in nasal epithelial cells. Free Radic. Biol. Med. 2011, 51, 444–453. [Google Scholar] [PubMed]
- Dai, J.; Gu, L.; Su, Y.; Wang, Q.; Zhao, Y.; Chen, X.; Deng, H.; Li, W.; Wang, G.; Li, K. International immunopharmacology inhibition of curcumin on in fluenza A virus infection and influenzal pneumonia via oxidative stress, TLR2/4, p38/JNK MAPK and NF- κ B pathways. Int. Immunopharmacol. 2018, 54, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.W.; Ou, Y.D.; Chu, Y.Y.; Wang, J.M.; Yu, C.Y.; Lin, C.F. Activation of Nrf2 by the dengue virus causes an increase in CLEC5A, which enhances TNF-α production by mononuclear phagocytes. Nat. Publ. Gr. 2016, 6, 1–15. [Google Scholar]
- Wu, Y.H.; Hao, B.J.; Cao, H.C.; Xu, W.; Li, Y.J.; Li, L.J. Anti-hepatitis B virus effect and possible mechanism of action of 3,4-O-dicaffeoylquinic acid in vitro and in vivo. Evid. Based Complement. Altern. Med. 2012, 2012, 1–10. [Google Scholar]
- Yazdi, S.E.; Prinsloo, G.; Heyman, H.M.; Oosthuizen, C.B.; Klimkait, T.; Meyer, J.J.M. Anti-HIV-1 activity of quinic acid isolated from Helichrysum mimetes using NMR-based metabolomics and computational analysis. S. Afr. J. Bot. 2019, 126, 328–339. [Google Scholar] [CrossRef]
- Kapoor, R.; Sharma, B.; Kanwar, S.S. Antiviral Phytochemicals: An Overview. Biochem. Physiol. Open Access. 2017, 6, 7. [Google Scholar] [CrossRef]
- Di Sotto, A.; Checconi, P.; Celestino, I.; Locatelli, M.; Carissimi, S.; De Angelis, M.; Rossi, V.; Limongi, D.; Toniolo, D.; Martinoli, L. Antiviral and antioxidant activity of a hydroalcoholic extract from H. lupulus L. Oxid. Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Plant Names | Family | Antiviral Activities | References |
|---|---|---|---|
| Adansonia digitata | Malvaceae | HSV-1, NDV, HSV-1; ASFV, HIV1 RT, HIV-FRET, PR | [33,34,35,36,37,38] |
| Artemisia afra | Asteraceae | HIV-1/2 | [37,39] |
| Aloe ferox | Asphodelaceae | HSV-1 | [40] |
| Carissa edulis | Apocynaceae | HSV-1, CDV, CPIV, FHV, LSDV, PV-2, CMV | [40,41,42,43,44] |
| Crinum macowanii | Amaryllidaceae | HIV-1 RT, PR | [45] |
| Elaeodendron croceum | Celastraceae | HIV-CB | [46] |
| Elaeodendron transvaalense | Celastraceae | HIV-1 a-Glucosidase, RT, CB, NF-kB, Tat, IN | [45,46,47] |
| Elephantorrhiza elephantina | Fabaceae | HIV-RT | [48] |
| Euclea natalensis | Ebenaceae | HIV-1 RT, HSV-1 | [49] |
| Helichrysum aureonitens | Asteraceae | HSV-1, Cox B-1, Ad31 reovirus | [50] |
| Heteropyxis natalensis | Heteropyxidaceae | HIV-1 RT | [51] |
| Lobostemon fruticosus | Boraginaceae | HIV-1 | [52] |
| Moringa oleifera | Moringaceae | HSV1, HIV-1 RT, FMDV, HBV, EBV | [50,51,52,53,54,55] |
| Peltophorum africanum | Fabaceae | HIV1-RT | [56,46] |
| Prunus africana | Rosaceae | CMV | [47] |
| Ricinus communis | Euphorbiaceae | HIV1- RT, RNase H, HIV-1 IN | [45,46,57] |
| Senna petersiana | Fabaceae | HIV1-RT | [58] |
| Sutherlandia frutescens | Fabaceae | HIV1 RT, IN, RNase H | [45,46,59] |
| Terminalia sericea | Combretaceae | HIV1 RT, HIV-1 RNA-dependent-DNA polymerase (RDDP) | [47,46] |
| Ziziphus mucronata | Rhamnaceae | HIV-1 RT, RNase H | [60] |
| Plant Names | LC50 (µg/mL) | DPPH EC50 (µg/mL) | ABTS EC50 (µg/mL) |
|---|---|---|---|
| Adansonia digitata | 291.5 | 4.64 | 5.04 |
| Artemisia afra | 151.8 | 20.41 | 16.39 |
| Aloe ferox | 330.3 | NT | NT |
| Carissa edulis | 400.6 | NT | NT |
| Crinum macowanii | 389.4 | NT | NT |
| Elaeodendron croceum | 394.4 | 6.00 | 4.12 |
| Elaeodendron ransvaalense | 336.9 | 11.64 | 15.00 |
| Elephantorrhiza elephantina | 225.2 | 6.54 | 7.40 |
| Euclea natalensis | 214.3 | 5.30 | 5.00 |
| Helichrysum aureonitens | 305.2 | 8.25 | 11.40 |
| Heteropyxis natalensis | 259.7 | NT | NT |
| Lobostemon fruticosus | 313.4 | NT | NT |
| Moringa oleifera | 271.6 | NT | NT |
| Peltophorum africanum | 332.5 | NT | NT |
| Prunus africana | 249.1 | NT | NT |
| Ricinus communis | 82.0 | NT | NT |
| Senna petersiana | 328.7 | NT | NT |
| Sutherlandia frutescens | 301.9 | 32.20 | 42.30 |
| Terminalia sericea | 232.2 | NT | NT |
| Ziziphus mucronata | 271.6 | NT | NT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
More, G.K.; Makola, R.T.; Prinsloo, G. In Vitro Evaluation of Anti-Rift Valley Fever Virus, Antioxidant and Anti-Inflammatory Activity of South African Medicinal Plant Extracts. Viruses 2021, 13, 221. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020221
More GK, Makola RT, Prinsloo G. In Vitro Evaluation of Anti-Rift Valley Fever Virus, Antioxidant and Anti-Inflammatory Activity of South African Medicinal Plant Extracts. Viruses. 2021; 13(2):221. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020221
Chicago/Turabian StyleMore, Garland K., Raymond T. Makola, and Gerhard Prinsloo. 2021. "In Vitro Evaluation of Anti-Rift Valley Fever Virus, Antioxidant and Anti-Inflammatory Activity of South African Medicinal Plant Extracts" Viruses 13, no. 2: 221. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020221