
Genetic Characterization and Pathogenesis of Avian Influenza Virus H7N3 Isolated from Spot-Billed Ducks in South Korea, Early 2019

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation of Influenza Virus from Wildbird Feces
2.3. Host Identification
2.4. Reverse Transcription (RT)-PCR and Sequencing
2.5. Next Generation Sequencing (NGS) by Illumina Hiseq X Method
2.6. Molecular Characterization and Phylogenetic Analysis
2.7. Determination of 50% Tissue Culture Infectious Dose (TCID50) and 50% Egg Infectious Dose (EID50)
2.8. Viral Growth Kinetics in MDCK Cells
2.9. Animal Experiment
2.10. Statistics
3. Results
3.1. Genome Characterization of H7N3 (WKU2019-1) Isolate
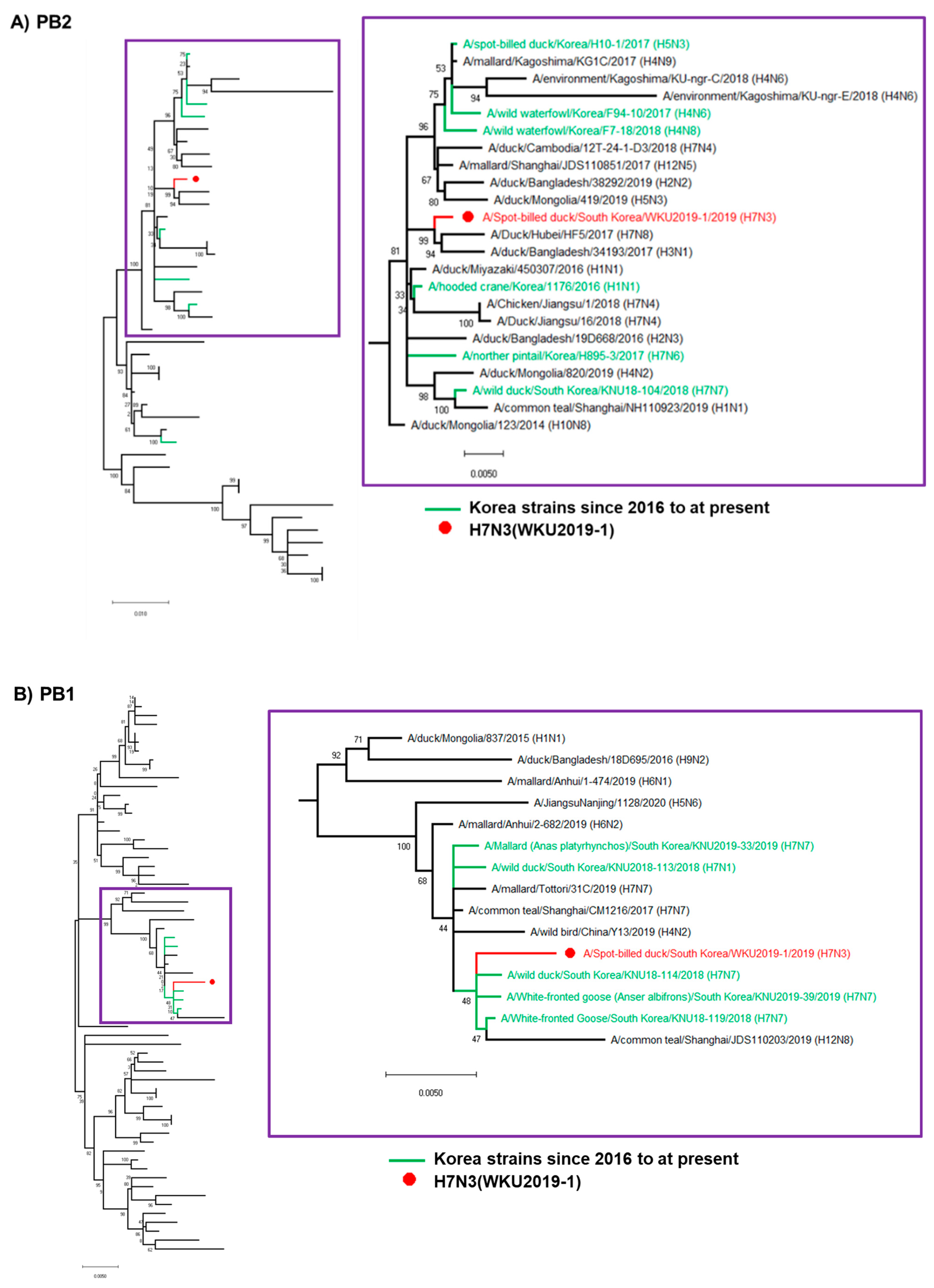
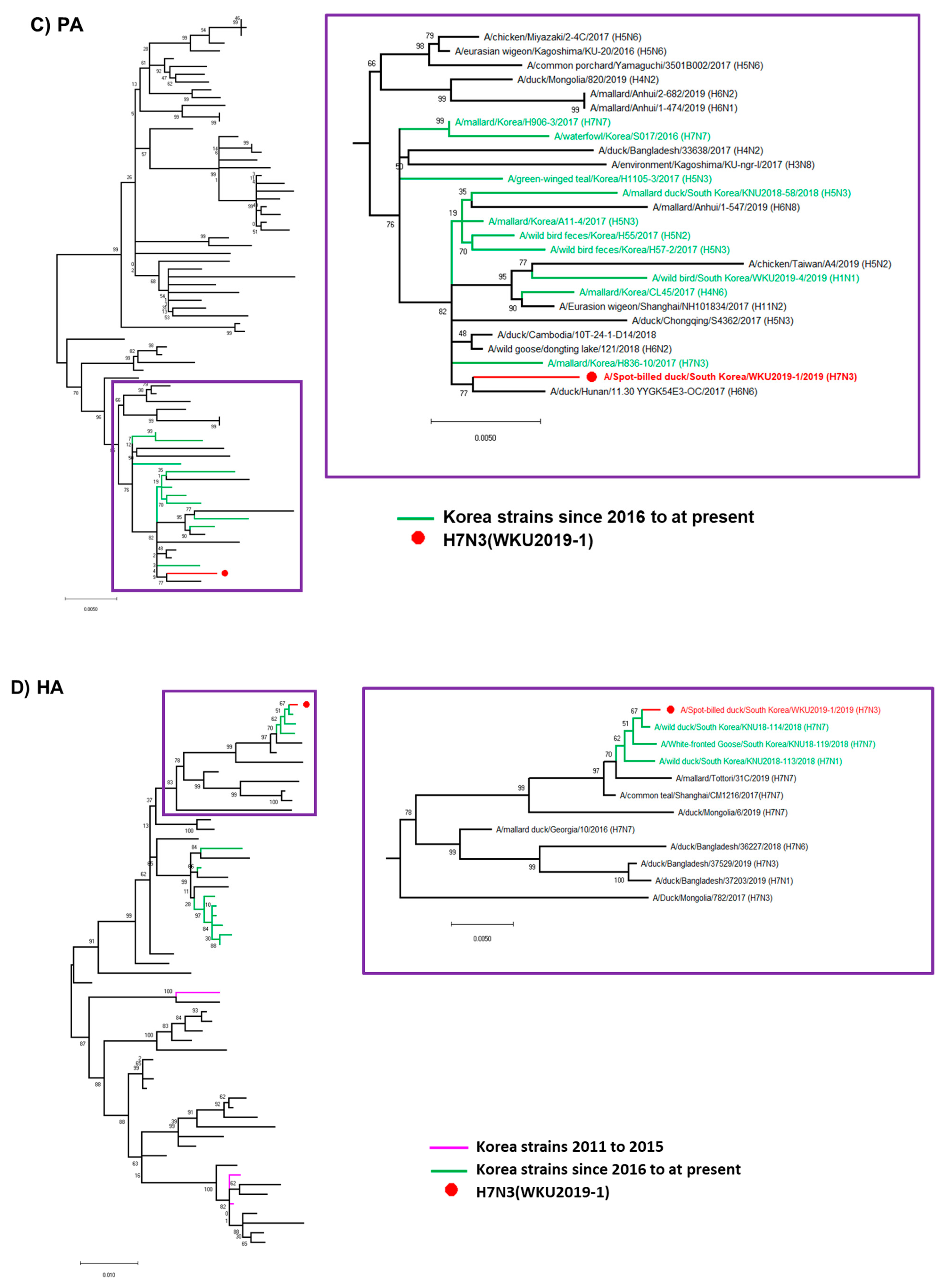
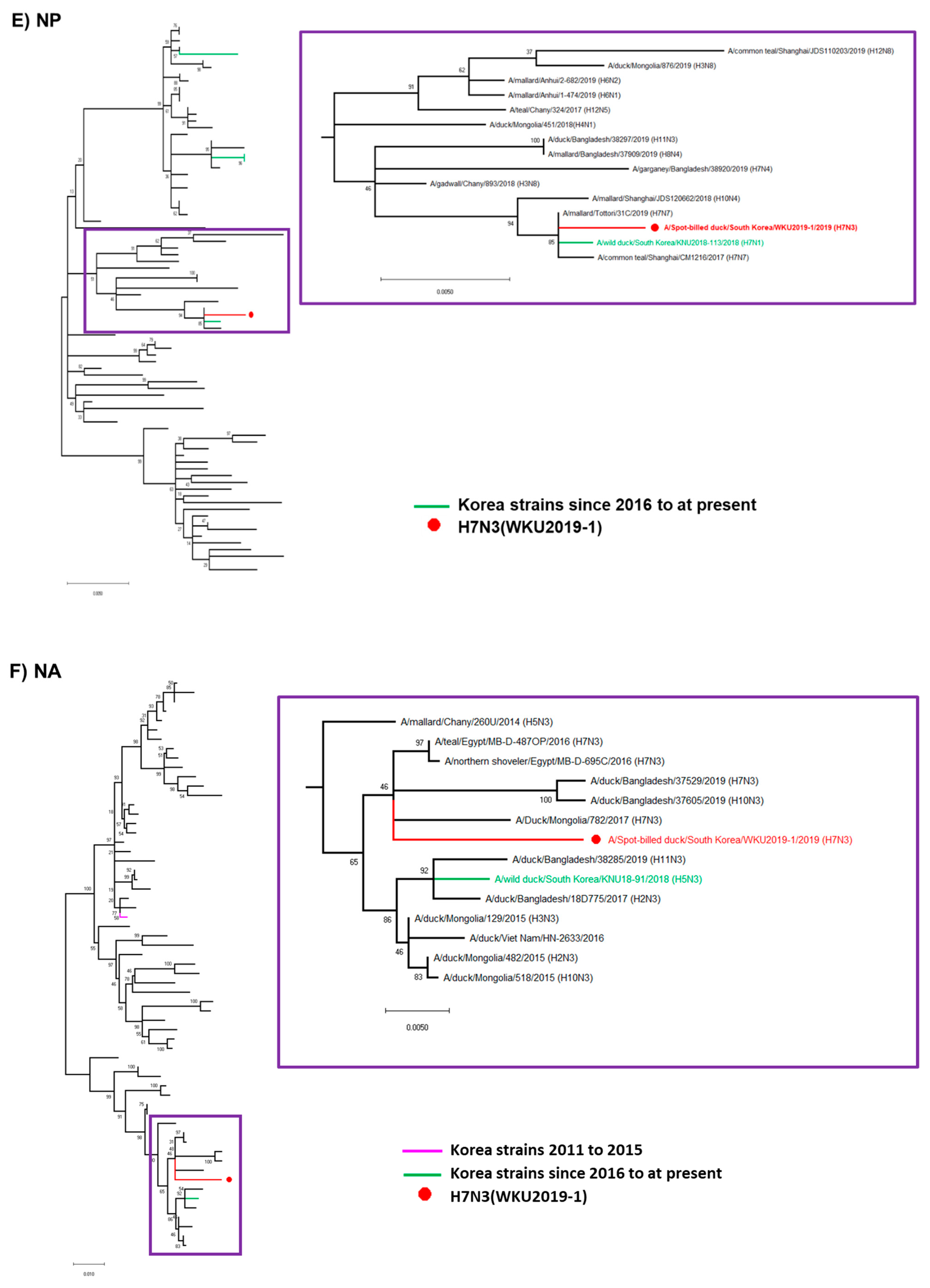
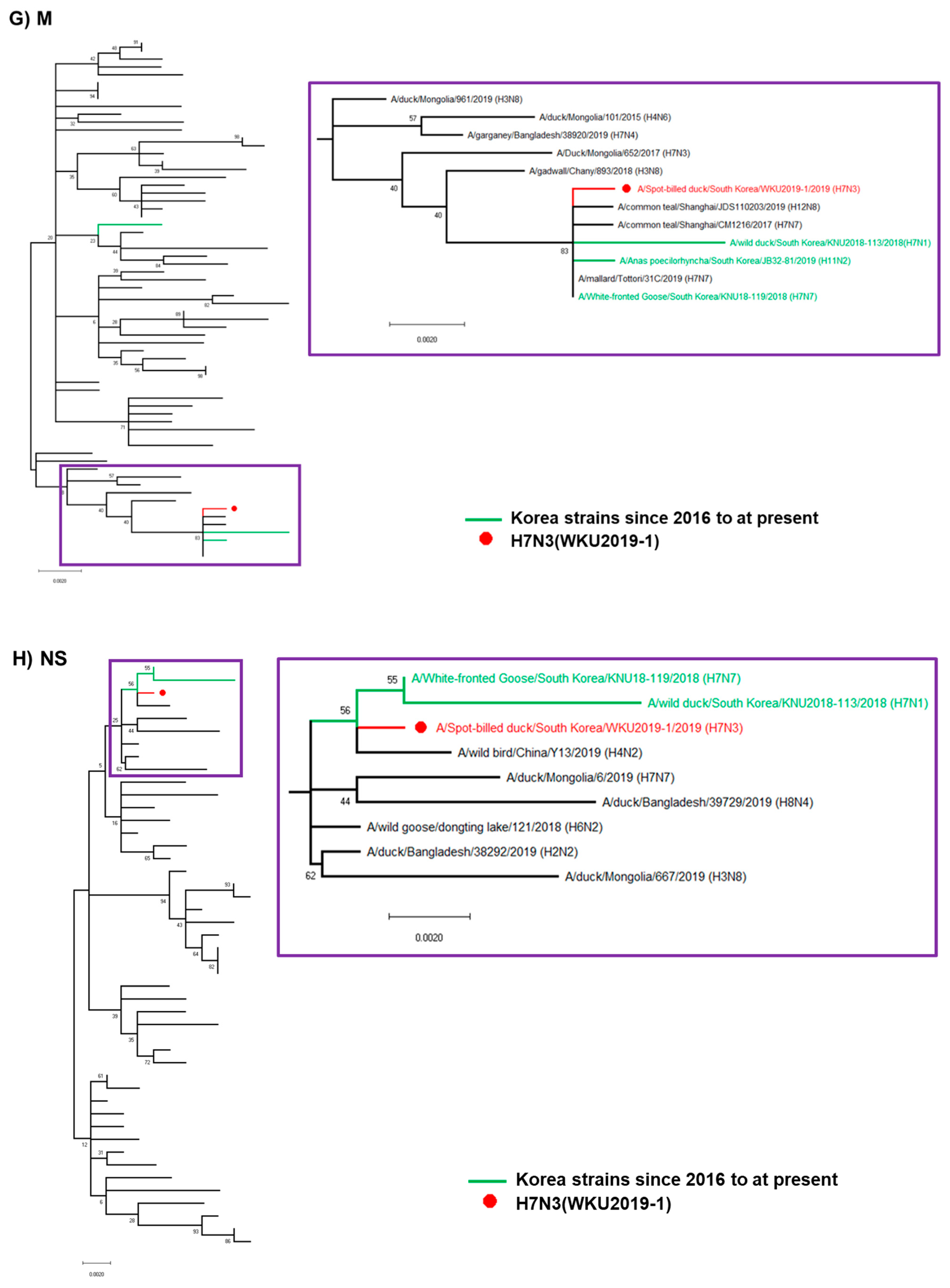
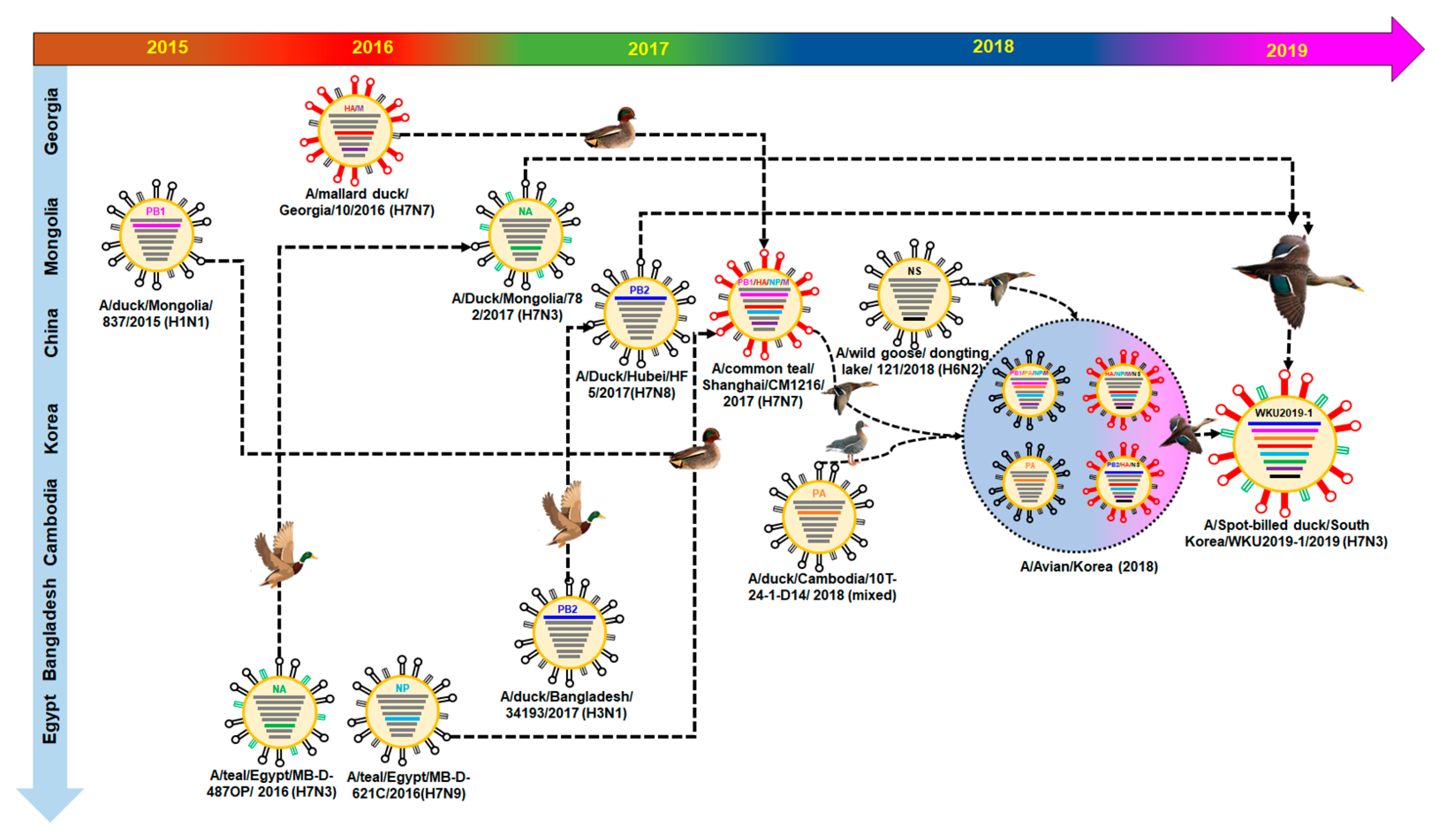
3.2. Hypothesis for Reassortment Event for Each Gene Segment
3.3. Molecular Characterization of the H7N3 (WKU2019-1) Isolate
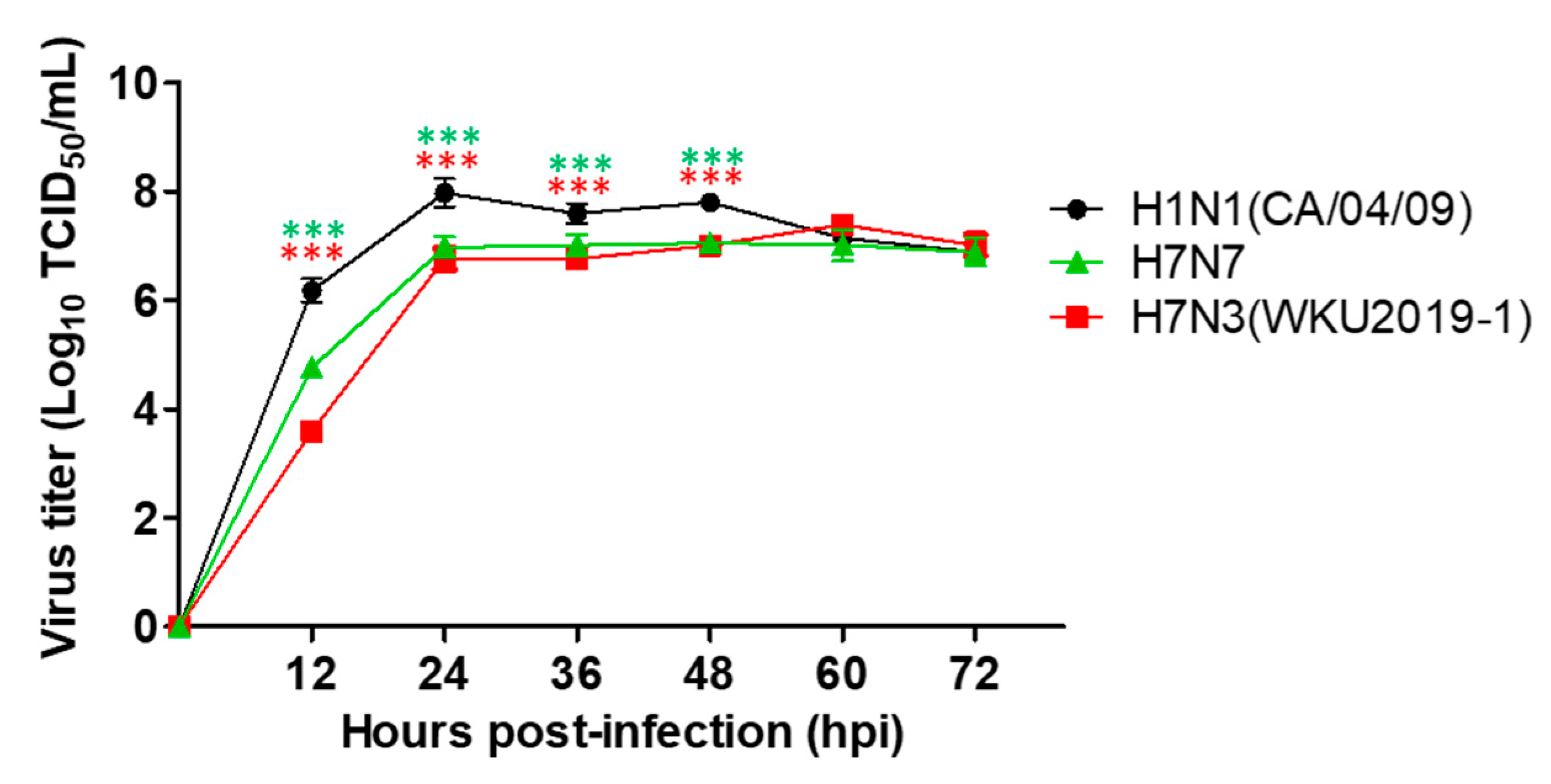
3.4. Growth Kinetics of H7N3 (WKU2019-1) Isolate in Mammalian Cell Culture
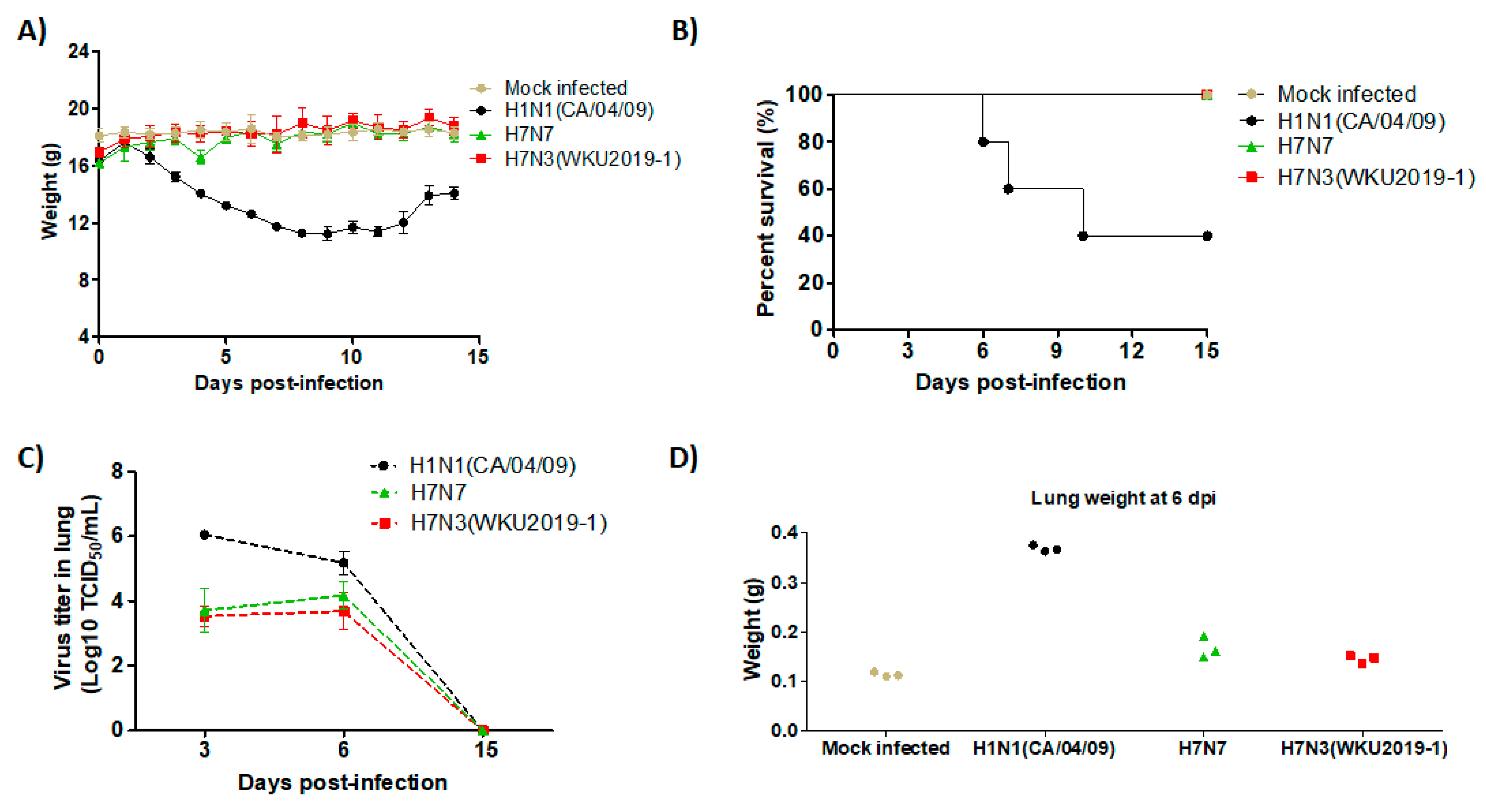
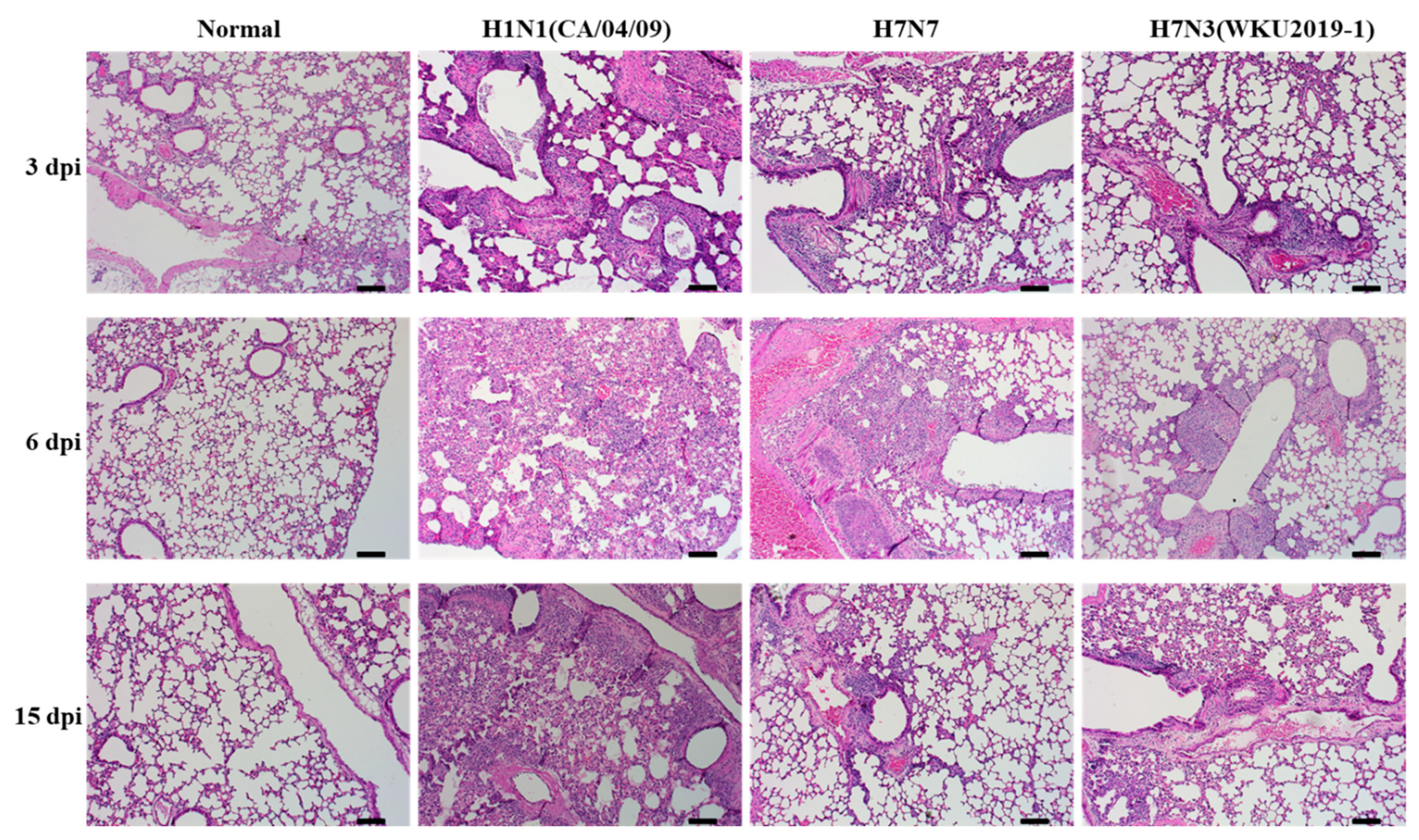
3.5. Pathogenicity in Mice
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shi, Y.; Cui, H.; Wang, J.; Chi, Q.; Li, X.; Teng, Q.; Chen, H.; Yang, J.; Liu, Q.; Li, Z. Characterizations of H4 avian influenza viruses isolated from ducks in live poultry markets and farm in Shanghai. Sci. Rep. 2016, 6, 37843. [Google Scholar] [CrossRef] [Green Version]
- Naguib, M.M.; Verhagen, J.H.; Mostafa, A.; Wille, M.; Li, R.; Graaf, A.; Järhult, J.D.; Ellström, P.; Zohari, S.; Lundkvist, Å.; et al. Global patterns of avian influenza A (H7): Virus evolution and zoonotic threats. FEMS Microbiol. Rev. 2019, 43, 608–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, J.W.; Ko, S.-Y.; Byoun, M.S.; Sung, H.W.; Lim, C.S.; Byun, M.S. GENEDIA Multi Influenza Ag Rapid Test for detection and H1, H3, and H5 subtyping of influenza viruses. J. Clin. Virol. 2015, 73, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Lupiani, B.; Reddy, S.M. The history of avian influenza. Comp. Immunol. Microbiol. Infect. Dis. 2009, 32, 311–323. [Google Scholar] [CrossRef]
- Zanin, M.; Koçer, Z.A.; Poulson, R.L.; Gabbard, J.D.; Howerth, E.W.; Jones, C.A.; Friedman, K.; Seiler, J.; Danner, A.; Kercher, L.; et al. Potential for Low-Pathogenic Avian H7 Influenza A Viruses to Replicate and Cause Disease in a Mammalian Model. J. Virol. 2016, 91. [Google Scholar] [CrossRef] [Green Version]
- Krauss, S.; Stucker, K.M.; Schobel, S.A.; Danner, A.; Friedman, K.; Knowles, J.P.; Kayali, G.; Niles, L.J.; Dey, A.D.; Raven, G.; et al. Long-term surveillance of H7 influenza viruses in American wild aquatic birds: Are the H7N3 influenza viruses in wild birds the precursors of highly pathogenic strains in domestic poultry? Emerg. Microbes Infect. 2015, 4, 1–9. [Google Scholar] [CrossRef]
- Subbarao, K.; Katz, J. Avian influenza viruses infecting humans. Cell. Mol. Life Sci. 2000, 57, 1770–1784. [Google Scholar] [CrossRef]
- Naeem, K. The Avian Influenza H7N3 Outbreak in South Central Asia. Avian Dis. 2003, 47, 31–35. [Google Scholar]
- Aamir, U.B.; Naeem, K.; Ahmed, Z.; Obert, C.A.; Franks, J.; Krauss, S.; Seiler, P.; Webster, R.G. Zoonotic potential of highly pathogenic avian H7N3 influenza viruses from Pakistan. Virology 2009, 390, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Lu, R.; Wo, E.; You, J.; Wang, Y.; Wang, Q.; Xu, L.; Wu, N.; Guo, C. Sequence and phylogenetic analysis of H7N3 avian influenza viruses isolated from poultry in China in 2011. Arch. Virol. 2012, 157, 2017–2021. [Google Scholar] [CrossRef]
- Suttie, A.; Yann, S.; Y, P.; Tum, S.; Deng, Y.-M.; Hul, V.; Horm, V.S.; Barr, I.; Greenhill, A.; Horwood, P.F.; et al. Detection of Low Pathogenicity Influenza A(H7N3) Virus during Duck Mortality Event, Cambodia, 2017. Emerg. Infect. Dis. 2018, 24, 1103–1107. [Google Scholar] [CrossRef] [Green Version]
- Shibata, A.; Harada, R.; Okamatsu, M.; Matsuno, K.; Arita, T.; Suzuki, Y.; Shirakura, M.; Odagiri, T.; Takemae, N.; Uchida, Y.; et al. Characterization of a novel reassortant H7N3 highly pathogenic avian influenza virus isolated from a poultry meat product taken on a passenger flight to Japan. J. Vet. Med. Sci. 2019, 81, 444–448. [Google Scholar] [CrossRef]
- Kim, H.-R.; Park, C.-K.; Lee, Y.-J.; Oem, J.-K.; Kang, H.-M.; Choi, J.-G.; Lee, O.-S.; Bae, Y.-C. Low pathogenic H7 subtype avian influenza viruses isolated from domestic ducks in South Korea and the close association with isolates of wild birds. J. Gen. Virol. 2012, 93, 1278–1287. [Google Scholar] [CrossRef]
- Nguyen, N.M.; Sung, H.W.; Yun, K.-J.; Park, H.; Yeo, S.-J. Genetic Characterization of a Novel North American-Origin Avian Influenza A (H6N5) Virus Isolated from Bean Goose of South Korea in 2018. Viruses 2020, 12, 774. [Google Scholar] [CrossRef]
- WHO. WHO Information for the Molecular Detection of Influenza Viruses; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Pancasakti, H.; Utari, W.D.; Raha, A.R. Phylogenetic Analysis of Duck Species from Tegal Indonesia Using 18S Ribosomal RNA and Mitochondrial COI Gene. Int. J. Poult. Sci. 2018, 17, 392–404. [Google Scholar] [CrossRef] [Green Version]
- Ambardar, S.; Gupta, R.; Trakroo, D.; Lal, R.; Vakhlu, J. High Throughput Sequencing: An Overview of Sequencing Chemistry. Indian J. Microbiol. 2016, 56, 394–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.H.; Lee, H.J.; Lee, Y.J.; Kang, H.M.; Jeong, O.M.; Kim, M.C.; Kwon, J.S.; Kwon, J.H.; Kim, C.B.; Lee, J.B.; et al. DNA barcoding techniques for avian influenza virus surveillance in migratory bird habitats. J. Wildl. Dis. 2010, 46, 649–654. [Google Scholar] [CrossRef]
- Trinh, T.-T.T.; Duong, B.T.; Nguyen, A.T.V.; Tuong, H.T.; Hoang, V.T.; Than, D.D.; Nam, S.-J.; Sung, H.W.; Yun, K.-J.; Yeo, S.-J.; et al. Emergence of Novel Reassortant H1N1 Avian Influenza Viruses in Korean Wild Ducks in 2018 and 2019. Viruses 2020, 13, 30. [Google Scholar] [CrossRef]
- Yeo, S.-J.; Than, D.-D.; Park, H.-S.; Sung, H.W.; Park, H. Molecular Characterization of a Novel Avian Influenza A (H2N9) Strain Isolated from Wild Duck in Korea in 2018. Viruses 2019, 11, 1046. [Google Scholar] [CrossRef] [Green Version]
- Tuong, H.T.; Nguyen, N.M.; Sung, H.W.; Park, H.; Yeo, S.-J. Genetic Characterization of Avian Influenza A (H11N9) Virus Isolated from Mandarin Ducks in South Korea in 2018. Viruses 2020, 12, 203. [Google Scholar] [CrossRef] [Green Version]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Jiménez-Bluhm, P.; Karlsson, E.A.; Ciuoderis, K.A.; Cortez, V.; Marvin, S.A.; Hamilton-West, C.; Schultz-Cherry, S.; Osorio, J.E. Avian H11 influenza virus isolated from domestic poultry in a Colombian live animal market. Emerg. Microbes Infect. 2016, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Killian, M.L. Hemagglutination Assay for Influenza Virus. Methods Mol. Biol. 2014, 116, 3–9. [Google Scholar]
- Vertebrate Animal Research. 2018. Available online: https://animal.research.uiowa.edu/iacuc-guidelines-anesthesia (accessed on 1 January 2021).
- WHO Global Influenza Surveillance Network. Manual for the Laboratory Diagnosis and Virological Surveillance of Influenza; WHO: Geneva, Switzerland, 2011; p. 153. [Google Scholar]
- Heider, A.; Mochalova, L.; Harder, T.; Tuzikov, A.; Bovin, N.; Wolff, T.; Matrosovich, M.; Schweiger, B. Alterations in hemagglutinin receptor-binding specificity accompany the emergence of highly pathogenic avian influenza viruses. J. Virol. 2015, 89, 5395–5405. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Feng, H.; Xu, J.; Zhao, D.; Shi, J.; Li, Y.; Deng, G.; Jiang, Y.; Li, X.; Zhu, P.; et al. The PA Protein Directly Contributes to the Virulence of H5N1 Avian Influenza Viruses in Domestic Ducks. J. Virol. 2011, 85, 2180–2188. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Li, X.; Tang, L.; Wang, T.; He, G. Characterization of the low-pathogenic H7N7 avian influenza virus in Shanghai, China. Poult. Sci. 2021, 100, 565–574. [Google Scholar] [CrossRef]
- Li, J.; Li, Y.; Hu, Y.; Chang, G.; Sun, W.; Yang, Y.; Kang, X.; Wu, X.; Zhu, Q. PB1-mediated virulence attenuation of H5N1 influenza virus in mice is associated with PB2. J. Gen. Virol. 2011, 92, 1435–1444. [Google Scholar] [CrossRef]
- Li, J.; Ishaq, M.; Prudence, M.; Xi, X.; Hu, T.; Liu, Q.; Guo, D. Single mutation at the amino acid position 627 of PB2 that leads to increased virulence of an H5N1 avian influenza virus during adaptation in mice can be compensated by multiple mutations at other sites of PB2. Virus Res. 2009, 144, 123–129. [Google Scholar] [CrossRef]
- Prokopyeva, E.; Sobolev, I.; Prokopyev, M.; Shestopalov, A. Adaptation of influenza A(H1N1)pdm09 virus in experimental mouse models. Infect. Genet. Evol. 2016, 39, 265–271. [Google Scholar] [CrossRef]
- Govorkova, E.A.; Rehg, J.E.; Krauss, S.; Yen, H.-L.; Guan, Y.; Peiris, M.; Nguyen, T.D.; Hanh, T.H.; Puthavathana, P.; Long, H.T.; et al. Lethality to Ferrets of H5N1 Influenza Viruses Isolated from Humans and Poultry in 2004. J. Virol. 2005, 79, 2191–2198. [Google Scholar] [CrossRef] [Green Version]
- Salomon, R.; Franks, J.; Govorkova, E.A.; Ilyushina, N.A.; Yen, H.-L.; Hulse-Post, D.J.; Humberd, J.; Trichet, M.; Rehg, J.E.; Webby, R.J.; et al. The polymerase complex genes contribute to the high virulence of the human H5N1 influenza virus isolate A/Vietnam/1203/04. J. Exp. Med. 2006, 203, 689–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsurumura, T.; Qiu, H.; Yoshida, T.; Tsumori, Y.; Tsuge, H. Crystallization and preliminary X-ray diffraction studies of a surface mutant of the middle domain of PB2 from human influenza A (H1N1) virus. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2014, 70, 72–75. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, G.; Dauber, B.; Wolff, T.; Planz, O.; Klenk, H.-D.; Stech, J. The viral polymerase mediates adaptation of an avian influenza virus to a mammalian host. Proc. Natl. Acad. Sci. USA 2005, 102, 18590–18595. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, G.; Abram, M.; Keiner, B.; Wagner, R.; Klenk, H.-D.; Stech, J. Differential Polymerase Activity in Avian and Mammalian Cells Determines Host Range of Influenza Virus. J. Virol. 2007, 81, 9601–9604. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Sun, W.; Li, X.; Chen, Q.; Chai, H.; Gao, X.; Guo, J.; Zhang, K.; Wang, T.; Feng, N.; et al. Adaptive amino acid substitutions enhance the virulence of a reassortant H7N1 avian influenza virus isolated from wild waterfowl in mice. Virology 2015, 476, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Taubenberger, J.K.; Reid, A.H.; Lourens, R.M.; Wang, R.; Jin, G.; Fanning, T.G. Characterization of the 1918 influenza virus polymerase genes. Nature 2005, 437, 889–893. [Google Scholar] [CrossRef]
- Hulse-Post, D.J.; Franks, J.; Boyd, K.; Salomon, R.; Hoffmann, E.; Yen, H.L.; Webby, R.J.; Walker, D.; Nguyen, T.D.; Webster, R.G. Molecular Changes in the Polymerase Genes (PA and PB1) Associated with High Pathogenicity of H5N1 Influenza Virus in Mallard Ducks. J. Virol. 2007, 81, 8515–8524. [Google Scholar] [CrossRef] [Green Version]
- Wei, K.; Sun, H.; Sun, Z.; Sun, Y.; Kong, W.; Pu, J.; Ma, G.; Yin, Y.; Yang, H.; Guo, X.; et al. Influenza A Virus Acquires Enhanced Pathogenicity and Transmissibility after Serial Passages in Swine. J. Virol. 2014, 88, 11981–11994. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Hu, W.-B.; Xu, K.; He, Y.-X.; Wang, T.-Y.; Chen, Z.; Li, T.-X.; Liu, J.-H.; Buchy, P.; Sun, B. Amino acids 473V and 598P of PB1 from an avian-origin influenza A virus contribute to polymerase activity, especially in mammalian cells. J. Gen. Virol. 2012, 93, 531–540. [Google Scholar] [CrossRef]
- Conenello, G.M.; Tisoncik, J.R.; Rosenzweig, E.; Varga, Z.T.; Palese, P.; Katze, M.G. A Single N66S Mutation in the PB1-F2 Protein of Influenza A Virus Increases Virulence by Inhibiting the Early Interferon Response In Vivo. J. Virol. 2011, 85, 652–662. [Google Scholar] [CrossRef] [Green Version]
- Schmolke, M.; Manicassamy, B.; Pena, L.; Sutton, T.; Hai, R.; Varga, Z.T.; Hale, B.G.; Steel, J.; Pérez, D.R.; García-Sastre, A. Differential Contribution of PB1-F2 to the Virulence of Highly Pathogenic H5N1 Influenza A Virus in Mammalian and Avian Species. PLoS Pathog. 2011, 7, e1002186. [Google Scholar] [CrossRef] [Green Version]
- Ozawa, M.; Basnet, S.; Burley, L.M.; Neumann, G.; Hatta, M.; Kawaoka, Y. Impact of Amino Acid Mutations in PB2, PB1-F2, and NS1 on the Replication and Pathogenicity of Pandemic (H1N1) 2009 Influenza Viruses. J. Virol. 2011, 85, 4596–4601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanmuganatham, K.K.; Jones, J.C.; Marathe, B.M.; Feeroz, M.M.; Jones-Engel, L.; Walker, D.; Turner, J.; Rabiul Alam, S.M.; Kamrul Hasan, M.; Akhtar, S.; et al. The replication of Bangladeshi H9N2 avian influenza viruses carrying genes from H7N3 in mammals. Emerg. Microbes Infect. 2016, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lycett, S.J.; Ward, M.J.; Lewis, F.I.; Poon, A.F.Y.; Kosakovsky Pond, S.L.; Brown, A.J.L. Detection of Mammalian Virulence Determinants in Highly Pathogenic Avian Influenza H5N1 Viruses: Multivariate Analysis of Published Data. J. Virol. 2009, 83, 9901–9910. [Google Scholar] [CrossRef] [Green Version]
- Min, J.-Y.; Li, S.; Sen, G.C.; Krug, R.M. A site on the influenza A virus NS1 protein mediates both inhibition of PKR activation and temporal regulation of viral RNA synthesis. Virology 2007, 363, 236–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Jiang, Y.; Jiao, P.; Wang, A.; Zhao, F.; Tian, G.; Wang, X.; Yu, K.; Bu, Z.; Chen, H. The NS1 Gene Contributes to the Virulence of H5N1 Avian Influenza Viruses. J. Virol. 2006, 80, 11115–11123. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Zou, X.; Zhou, J.; Tang, J.; Shu, Y. Residues 41V and/or 210D in the NP protein enhance polymerase activities and potential replication of novel influenza (H7N9) viruses at low temperature. Virol. J. 2015, 12, 71. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Chen, H.; Huang, J.; Chen, Y.; Gu, M.; Wang, X.; Hu, S.; Liu, X.; Liu, X. A nonpathogenic duck-origin H9N2 influenza A virus adapts to high pathogenicity in mice. Arch. Virol. 2014, 159, 2243–2252. [Google Scholar] [CrossRef]
- Li, J.; Zheng, W.; Hou, L.; Chen, C.; Fan, W.; Qu, H.; Jiang, J.; Liu, J.; Gao, G.F.; Zhou, J.; et al. Differential nucleocytoplasmic shuttling of the nucleoprotein of influenza a viruses and association with host tropism. Cell. Microbiol. 2017, 19, e12692. [Google Scholar] [CrossRef] [Green Version]
- Yamayoshi, S.; Yamada, S.; Fukuyama, S.; Murakami, S.; Zhao, D.; Uraki, R.; Watanabe, T.; Tomita, Y.; Macken, C.; Neumann, G.; et al. Virulence-Affecting Amino Acid Changes in the PA Protein of H7N9 Influenza A Viruses. J. Virol. 2014, 88, 3127–3134. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, R.; Yamada, S.; Le, M.Q.; Ito, M.; Sakai-Tagawa, Y.; Kawaoka, Y. Mammalian Adaptive Mutations of the PA Protein of Highly Pathogenic Avian H5N1 Influenza Virus. J. Virol. 2015, 89, 4117–4125. [Google Scholar] [CrossRef] [Green Version]
- Leung, B.W.; Chen, H.; Brownlee, G.G. Correlation between polymerase activity and pathogenicity in two duck H5N1 influenza viruses suggests that the polymerase contributes to pathogenicity. Virology 2010, 401, 96–106. [Google Scholar] [CrossRef] [Green Version]
- Mei, K.; Liu, G.; Chen, Z.; Gao, Z.; Zhao, L.; Jin, T.; Yu, X.; Chen, Q. Deep sequencing reveals the viral adaptation process of environment-derived H10N8 in mice. Infect. Genet. Evol. 2016, 37, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Shaw, M.; Cooper, L.; Xu, X.; Thompson, W.; Krauss, S.; Guan, Y.; Zhou, N.; Klimov, A.; Cox, N.; Webster, R.; et al. Molecular changes associated with the transmission of avian influenza a H5N1 and H9N2 viruses to humans. J. Med. Virol. 2002, 66, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Xu, J.; Shi, J.; Li, Y.; Chen, H. Synergistic Effect of S224P and N383D Substitutions in the PA of H5N1 Avian Influenza Virus Contributes to Mammalian Adaptation. Sci. Rep. 2015, 5, 10510. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Hu, Z.; Zhang, X.; Chen, M.; Wang, Z.; Xu, G.; Bi, Y.; Tong, Q.; Wang, M.; Sun, H.; et al. An R195K Mutation in the PA-X Protein Increases the Virulence and Transmission of Influenza A Virus in Mammalian Hosts. J. Virol. 2020, 94. [Google Scholar] [CrossRef]
- Lee, M.S.; Deng, M.C.; Lin, Y.J.; Chang, C.Y.; Shieh, H.K.; Shiau, J.Z.; Huang, C.C. Characterization of an H5N1 avian influenza virus from Taiwan. Vet. Microbiol. 2007, 124, 193–201. [Google Scholar] [CrossRef]
- Katz, J.M.; Lu, X.; Tumpey, T.M.; Smith, C.B.; Shaw, M.W.; Subbarao, K. Molecular Correlates of Influenza A H5N1 Virus Pathogenesis in Mice. J. Virol. 2000, 74, 10807–10810. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Deng, G.; Song, J.; Tian, G.; Suo, Y.; Jiang, Y.; Guan, Y.; Bu, Z.; Kawaoka, Y.; Chen, H. Two amino acid residues in the matrix protein M1 contribute to the virulence difference of H5N1 avian influenza viruses in mice. Virology 2009, 384, 28–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez, D.L.; Senne, D.A.; Banks, J.; Brown, I.H.; Essen, S.C.; Lee, C.-W.; Manvell, R.J.; Mathieu-Benson, C.; Moreno, V.; Pedersen, J.C.; et al. Recombination Resulting in Virulence Shift in Avian Influenza Outbreak, Chile. Emerg. Infect. Dis. 2004, 10, 693–699. [Google Scholar] [CrossRef]
- Velkers, F.C.; Bouma, A.; Matthijs, M.G.R.; Koch, G.; Westendorp, S.T.; Stegeman, J.A. Outbreak of avian influenza H7N3 on a turkey farm in the Netherlands. Vet. Rec. 2006, 159, 403–405. [Google Scholar] [CrossRef] [Green Version]
- Tweed, S.A.; Skowronski, D.M.; David, S.T.; Larder, A.; Petric, M.; Lees, W.; Li, Y.; Katz, J.; Krajden, M.; Tellier, R.; et al. Human Illness from Avian Influenza H7N3, British Columbia. Emerg. Infect. Dis. 2004, 10, 2196–2199. [Google Scholar] [CrossRef]
- Youk, S.; Lee, D.-H.; Ferreira, H.L.; Afonso, C.L.; Absalon, A.E.; Swayne, D.E.; Suarez, D.L.; Pantin-Jackwood, M.J. Rapid evolution of Mexican H7N3 highly pathogenic avian influenza viruses in poultry. PLoS ONE 2019, 14, e0222457. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Peng, R.; Zhang, W.; Qi, J.; Song, H.; Liu, S.; Wang, H.; Wang, M.; Xiao, H.; Fu, L.; et al. Avian-to-Human Receptor-Binding Adaptation of Avian H7N9 Influenza Virus Hemagglutinin. Cell Rep. 2019, 29, 2217–2228.e5. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Bank ID | Reference Strain Accession ID | Origin | Per Ident (%) |
|---|---|---|---|---|
| PB2 | MT845654 | KY402062 | A/hooded crane/Korea/1176/2016 (H1N1) | 99.25 (2280/2280) |
| MH458919 | A/Duck/Hubei/HF5/2017 (H7N8) | 99.17 (2280/2280) | ||
| MH791830 | A/duck/Bangladesh/34193/2017 (H3N1) | 99.17 (2295/2280) | ||
| PB1 | MT845655 | MN602508 | A/White-fronted Goose/South Korea/KNU18-119/2018 (H7N7) | 99.61 (2287/2341) |
| MN602505 | A/wild duck/South Korea/KNU18-114/2018 (H7N7) | 99.56 (2287/2341) | ||
| MK554565 | A/common teal/Shanghai/CM1216/2017 (H7N7) | 99.56 (2252/2341) | ||
| PA | MT845656 | MN602506 | A/wild duck/South Korea/KNU18-114/2018 (H7N7) | 99.54 (2180/2223) |
| EPI_ISL_309223 | A/mallard/Korea/H836-10/2017 (H7N3) | 99.45 (2200/2223) | ||
| MN703036 | A/duck/Cambodia/10T-24-1-D14/2018 (mixed) | 99.36 (2209/2223) | ||
| HA | MT845657 | MN483232 | A/wild duck/South Korea/KNU18-114/2018 (H7N7) | 99.82 (1683/1731) |
| MN483237 | A/White-fronted Goose/South Korea/KNU18-119/2018 (H7N7) | 99.64 (1683/1731) | ||
| MN480525 | A/wild duck/South Korea/KNU2018-113/2018 (H7N1) | 99.53 (1708/1731) | ||
| MK554567 | A/common teal/Shanghai/CM1216/2017 (H7N7) | 99.51 (1639/1731) | ||
| NP | MT845658 | MN208011 | A/teal/Egypt/MB-D-487OP/2016 (H7N3) | 99.66 (1563/1497) |
| MN480533 | A/wild duck/South Korea/KNU2018-113/2018 (H7N1) | 99.53 (1541/1497) | ||
| MK554568 | A/common teal/Shanghai/CM1216/2017 (H7N7) | 99.53 (1499/1497) | ||
| NA | MT845659 | EPI_ISL_327473 | A/Duck/Mongolia/782/2017 (H7N3) | 97.85 (1446/1455) |
| MN208013 | A/teal/Egypt/MB-D-487OP/2016 (H7N3) | 99.48 (1450/1455) | ||
| MN208042 | A/northern shoveler/Egypt/MB-D-690C/2016 (H7N3) | 98.34 (1450/1455) | ||
| M2, M1 | MT845660 | MN483235 | A/wild duck/South Korea/KNU18-114/2018 (H7N7) | 100 (982/1028) |
| MN584917 | A/wild duck/South Korea/KNU2018-113/2018 (H7N1) | 99.90 (1006/1028) | ||
| MK554570 | A/common teal/Shanghai/CM1216/2017 (H7N7) | 99.90 (990/1028) | ||
| NEP, NS1 | MT845661 | MN483241 | A/White-fronted Goose/South Korea/KNU18-119/2018 (H7N7) | 99.77 (866/873) |
| MH727484 | A/wild goose/dongting lake/121/2018 (H6N2) | 99.65 (864/873) | ||
| MN480542 | A/wild duck/South Korea/KNU2018-113/2018 (H7N1) | 99.19 (865/873) |
| Virus Strain | HA Receptor-Binding Residues (H3 Numbering) | NA | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cleavage Sites | 138 | 158 | 183 | 186 | 190 | 221 | 225 | 226 | 228 | 391 | Deleted Range from 50–70 | 26 | 106 | 223 | 373 | 394 | |
| WKU2019-1 (H7N3) | PELPKGR↓GLF | A | T | H | G | E | P | G | Q | G | Q | No deletion | V | I | I | F | Q |
| Zhejiang-2011 (H7N3) | PEIPKGR↓GLF | A | T | H | G | E | P | G | Q | G | N | No deletion | I | I | I | F | Q |
| Japan-2018 (H7N3) | PEVPKRR↓TAR | A | T | H | V | E | P | G | Q | G | Q | No deletion | I | I | I | F | Q |
| Korea-2017 | PELPKGR↓GLF | A | T | H | G | E | P | G | Q | G | N | No deletion | I | I | I | F | Q |
| Zhejiang-2011 (H7N7) | PELPKGR↓GLF | A | T | H | G | E | P | G | Q | G | Q | - | - | - | - | - | - |
| Viral Protein | Amino Acid | H7N3 (WKU2019-1) | Zhejiang-2011 (H7N3) | Japan-2018 (H7N3) | Korea-2017 (H7N3) | Zhejiang-2011 (H7N7) | Phenotype | References |
|---|---|---|---|---|---|---|---|---|
| PB2 | T63I | I | I | I | I | I | Pathogenic in mice | [30] |
| L89V | V | E | V | V | V | Enhanced polymerase activity, Increased virulence in mice | [31] | |
| K251R | R | R | R | R | R | Increased virulence in mice | [32] | |
| G309D | D | D | D | D | D | Enhanced polymerase activity, Increased virulence in mice | [31] | |
| Q368R | R | R | R | R | R | Increased polymerase activity, Increased virulence in mammals | [33,34] | |
| H447Q | Q | Q | Q | Q | Q | Increased polymerase activity, Increased virulence in mammals | ||
| I471T | T | T | T | T | T | Change the surface electrostatic potential drastically | [35] | |
| R477G | G | G | G | G | G | Enhanced polymerase activity, Increased virulence in mice | [31] | |
| I495V | V | V | V | V | V | Enhanced polymerase activity, Increased virulence in mice | ||
| E627K | E | E | E | E | E | Human adaptation marker | ||
| A676T | T | T | T | T | T | Enhanced polymerase activity, Increased virulence in mice | [31] | |
| PB1 | D/A3V | V | V | V | V | - | Increased polymerase activity, Increased virulence in mammals | [33,34] |
| L13P | P | P | P | P | P | Increased polymerase activity, Increased virulence in mammals, Mammalian host marker, Amantadine resistance | [35,36] | |
| R207K | K | K | K | K | K | Increased polymerase activity in mammalian cells | [37] | |
| K328N | N | N | N | N | N | Increased polymerase activity, Increased virulence in mammals | [33,34] | |
| S375N | N | N | N | N | N | Increased polymerase activity, Increased virulence in mammals, Human host marker | [33,34,38] | |
| H436Y | Y | Y | Y | Y | Y | Increased polymerase activity and virulence in mallards, ferrets and mice | [39] | |
| A469T | T | T | T | T | T | Conferred in contact transmissibility in guinea pigs. | [40] | |
| L473V | V | V | V | V | V | Increased polymerase activity and replication efficiency | [41] | |
| V652A | A | A | A | A | A | Increased virulence in mice | [32] | |
| M677T | T | T | T | T | T | Pathogenic in mice | [30] | |
| PB1-F2 | N66S | S | N | N | N | S | Increased virulence in mammals | [42,43,44] |
| T68I | T | T | - | T | T | Increased virulence in mammals | [45] | |
| NEP | A/P42S | S | S | A | S | - | Increased virulence in mammals, Antagonism of interferon induction | [46] |
| T/D/V/R/A127N | N | N | R | N | - | Increased virulence in mammals | [46,47] | |
| V149A | A | A | A | A | - | Pathogenicity in mice, Antagonism of interferon induction | [48] | |
| T47A | E | E | E | E | E | |||
| T48N | A | A | S | A | A | |||
| M51I | R | R | R | R | R | |||
| NP | V41I | I | I | I | I | I | Might contribute to viral transmissibility | [49] |
| I109V | V | I | I | I | I | Mammalian host specific mutation | [45] | |
| R214K | R | N | R | R | R | Mammalian host specific mutation | ||
| F313Y | F | A | F | F | F | Mammalian host specific mutation | ||
| E372D | T | S | T | T | T | Mammalian host specific mutation | ||
| V105M | V | M | M | M | V | Contribute to the increased virulence of the H9N2 | [50] | |
| D210E | E | E | E | E | E | Might contribute to viral transmissibility | [49] | |
| F253I | I | A | I | I | I | Results in attenuated pathogenicity of the virus in mice | [51] | |
| I353V | V | V | I | I | V | Increased virulence in mice | [32] | |
| PA | D3V | D | E | E | E | - | Contribute to the increased virulence of the H9N2 | [50] |
| S37A | A | A | A | A | A | Significantly increased viral growth and polymerase activity in mammalian cells | [52] | |
| V44I | V | V | V | V | V | Enhance the replicative ability of an H5N1 virus in A549 cells and enhance its pathogenicity in mice | [53] | |
| H266R | R | R | R | R | R | Increased polymerase activity, Increased virulence in mammals and birds | [54] | |
| F277S | S | S | S | S | S | Adapt to mammalian hosts | [55] | |
| C278Q | Q | Q | Q | Q | Q | Adapt to mammalian hosts | ||
| E382D | E | E | D | E | D | Human host marker | [38,56] | |
| N383D | D | D | D | D | D | Enhanced the pathogenicity and viral replication of H5N1 virus in mice | [57,58] | |
| S/A515T | T | T | T | T | T | Increased polymerase activity, Increased virulence in mammals and birds | [54] | |
| L653P | P | P | P | P | P | Adapt to mammalian hosts | [55] | |
| PA-X | R195K | R | K | Increased virulence in mammals | [28] | |||
| M1 | V15I | V | I | S | I | V | Increase pathogenicity to mice | [59] |
| V15I/T | V | S | V | I | V | Increased virulence in mammals | [28,29,60] | |
| N30D | D | F | D | D | D | Increased virulence in mammals | [61] | |
| V115I | V | V | V | L | - | Human host marker | [62] | |
| T121A | T | T | T | A | - | Human host marker | [61] | |
| M128R | M | M | M | L | - | Increased virulence in mice | [37] | |
| A166V | V | V | A | V | - | Contribute to the increased virulence of the H9N2. | [50] | |
| S183A | S | S | S | T | - | Resulted in the failure of virus production | [62] | |
| T185A | T | T | T | K | - | Resulted in the failure of virus production | ||
| T215A | A | A | A | R | - | Increased virulence in mammals | ||
| M2 | L55F | L | L | F | Y | L | Enhanced Transmission | [56] |
| L26F | L | L | L | I | L | Adamantine resistance mutation | [45] | |
| S31N | S | S | N | I | S | Adamantine resistance mutation/Antiviral resistance S31 (amanta) | [59] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trinh, T.-T.T.; Tiwari, I.; Durairaj, K.; Duong, B.T.; Nguyen, A.T.V.; Tuong, H.T.; Hoang, V.T.; Than, D.D.; Nam, S.; Yeo, S.-J.; et al. Genetic Characterization and Pathogenesis of Avian Influenza Virus H7N3 Isolated from Spot-Billed Ducks in South Korea, Early 2019. Viruses 2021, 13, 856. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050856
Trinh T-TT, Tiwari I, Durairaj K, Duong BT, Nguyen ATV, Tuong HT, Hoang VT, Than DD, Nam S, Yeo S-J, et al. Genetic Characterization and Pathogenesis of Avian Influenza Virus H7N3 Isolated from Spot-Billed Ducks in South Korea, Early 2019. Viruses. 2021; 13(5):856. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050856
Chicago/Turabian StyleTrinh, Thuy-Tien Thi, Indira Tiwari, Kaliannan Durairaj, Bao Tuan Duong, Anh Thi Viet Nguyen, Hien Thi Tuong, Vui Thi Hoang, Duong Duc Than, SunJeong Nam, Seon-Ju Yeo, and et al. 2021. "Genetic Characterization and Pathogenesis of Avian Influenza Virus H7N3 Isolated from Spot-Billed Ducks in South Korea, Early 2019" Viruses 13, no. 5: 856. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050856