
Bat Flies of the Family Streblidae (Diptera: Hippoboscoidea) Host Relatives of Medically and Agriculturally Important “Bat-Associated” Viruses

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
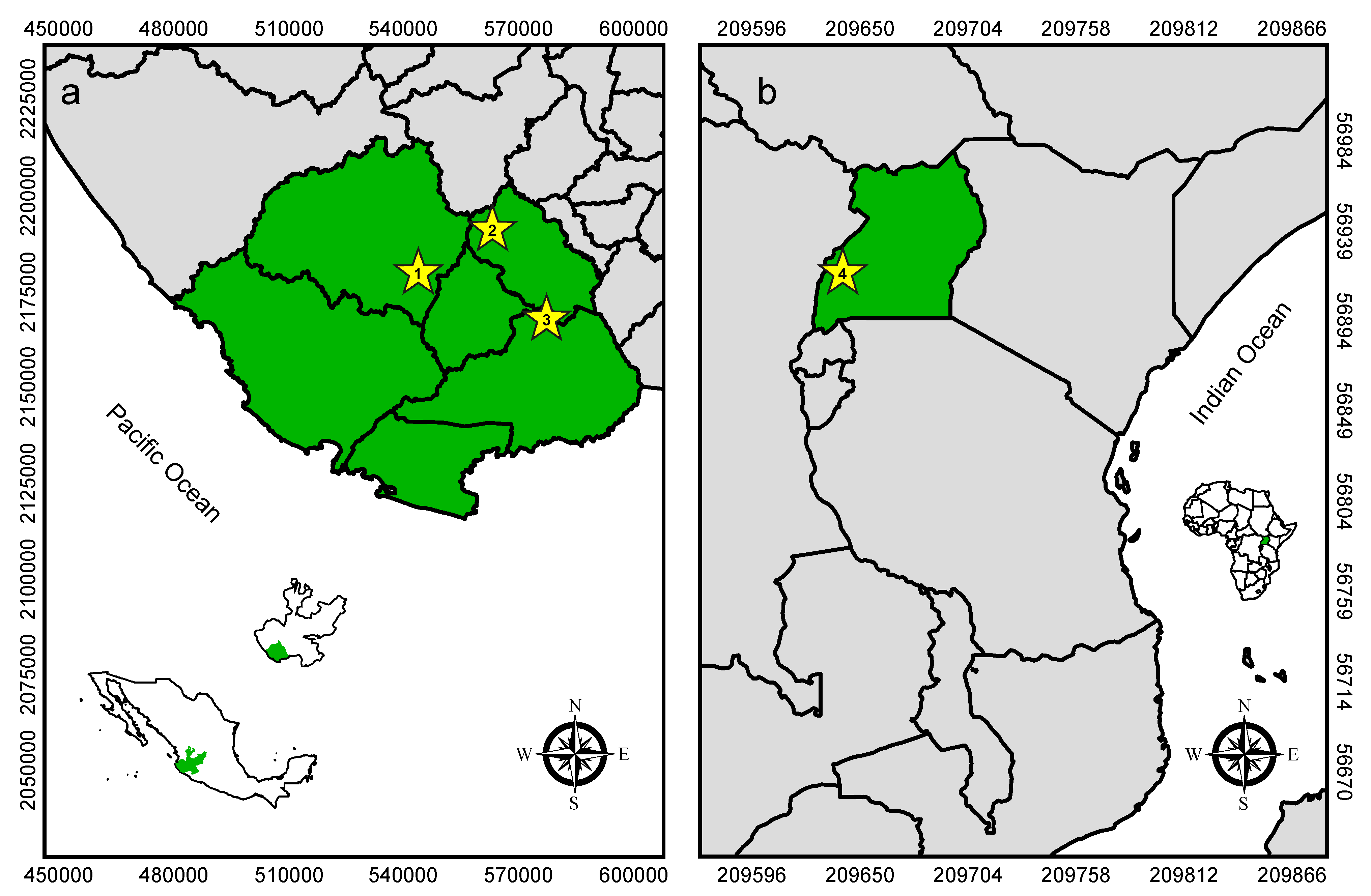
2.1. Study Sites and Sampling
2.2. Imaging of Bat Flies
2.3. DNA Barcoding of Bat Flies
2.4. Metagenomics and Phylogenetics
3. Results
3.1. Bat Fly Identification
3.2. Virus Characterization
3.3. Virus Prevalence and Load
4. Discussion
Supplementary Materials
Author Contributions
Funding
Animal Use Statement
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dick, C.W.; Patterson, B.D. Bat flies: Obligate ectoparasites of bats. In Micromammals and Macroparasites; Morand, S., Krasnov, B.R., Poulin, R., Eds.; Springer: New York, NY, USA, 2006. [Google Scholar]
- Peterson, B.V.; Wenzel, R.L. Nycteribiidae. In Manual of Nearctic Diptera; McAlpine, J.F., Peterson, B.V.E., Shewell, G.E., Teskey, H.J., Vockeroth, J.R., Wood, D.M., Eds.; Research Branch, Agriculture Canada: Ottawa, ON, Canada, 1987; Volume 2, pp. 1283–1291. [Google Scholar]
- Graciolli, G.; Dick, C.W.; Guerrero, R. Family Nycteribiidae. Zootaxa 2016, 4122, 780–783. [Google Scholar] [CrossRef] [PubMed]
- Dick, C.W.; Graciolli, G.; Guerrero, R. Family Streblidae. Zootaxa 2016, 4122, 784–802. [Google Scholar] [CrossRef]
- Dittmar, K.; Morse, S.F.; Dick, C.W.; Patterson, B.D. Bat fly evolution from the Eocene to the present (Hippoboscoidea, Streblidae and Nycteribiidae). In Parasite Diversity and Diversification: Evolutionary Ecology Meets Phylogenetics; Morand, S., Krasnov, B.R., Littlewood, T., Eds.; Cambridge University Press: Cambridge, UK, 2015; pp. 246–264. [Google Scholar]
- Obame-Nkoghe, J.; Rahola, N.; Bourgarel, M.; Yangari, P.; Prugnolle, F.; Maganga, G.D.; Leroy, E.M.; Fontenille, D.; Ayala, D.; Paupy, C. Bat flies (Diptera: Nycteribiidae and Streblidae) infesting cave-dwelling bats in Gabon: Diversity, dynamics and potential role in Polychromophilus melanipherus transmission. Parasit. Vectors 2016, 9, 333. [Google Scholar] [CrossRef] [Green Version]
- Gardner, R.A.; Molyneux, D.H. Polychromophilus murinus: A malarial parasite of bats: Life-history and ultrastructural studies. Parasitology 1988, 96, 591–605. [Google Scholar] [CrossRef]
- Morse, S.F.; Olival, K.J.; Kosoy, M.; Billeter, S.; Patterson, B.D.; Dick, C.W.; Dittmar, K. Global distribution and genetic diversity of Bartonella in bat flies (Hippoboscoidea, Streblidae, Nycteribiidae). Infect. Genet. Evol. 2012, 12, 1717–1723. [Google Scholar] [CrossRef] [PubMed]
- Billeter, S.A.; Hayman, D.T.; Peel, A.J.; Baker, K.; Wood, J.L.; Cunningham, A.; Suu-Ire, R.; Dittmar, K.; Kosoy, M.Y. Bartonella species in bat flies (Diptera: Nycteribiidae) from western Africa. Parasitology 2012, 139, 324–329. [Google Scholar] [CrossRef]
- Xu, Z.; Yang, W.; Feng, Y.; Li, Y.; Fu, S.; Li, X.; Song, J.; Zhang, H.; Zhang, Y.; Liu, W.J.; et al. Isolation and Identification of a highly divergent Kaeng Khoi virus from bat flies (Eucampsipoda sundaica) in China. Vector Borne Zoonotic Dis. 2019, 19, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Van Vuren, P.J.; Wiley, M.R.; Palacios, G.; Storm, N.; Markotter, W.; Birkhead, M.; Kemp, A.; Paweska, J.T. Isolation of a novel orthobunyavirus from bat flies (Eucampsipoda africana). J. Gen. Virol. 2017, 98, 935–945. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Li, Y.; Fu, S.; Li, X.; Song, J.; Zhang, H.; Yang, W.; Zhang, Y.; Pan, H.; Liang, G. Isolation of Kaeng Khoi virus (KKV) from Eucampsipoda sundaica bat flies in China. Virus Res. 2017, 238, 94–100. [Google Scholar] [CrossRef]
- Van Vuren, P.J.; Wiley, M.; Palacios, G.; Storm, N.; McCulloch, S.; Markotter, W.; Birkhead, M.; Kemp, A.; Paweska, J.T. Isolation of a novel fusogenic orthoreovirus from Eucampsipoda africana bat flies in South Africa. Viruses 2016, 8, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, A.J.; Paskey, A.C.; Kuhn, J.H.; Bishop-Lilly, K.A.; Goldberg, T.L. Diversity, transmission, and cophylogeny of ledanteviruses (Rhabdoviridae: Ledantevirus) and nycteribiid bat flies parasitizing Angolan soft-furred fruit bats in Bundibugyo District, Uganda. Microorganisms 2020, 8, 750. [Google Scholar] [CrossRef]
- Moskaluk, A.E.; Stuckey, M.J.; Jaffe, D.A.; Kasten, R.W.; Aguilar-Setien, A.; Olave-Leyva, J.I.; Galvez-Romero, G.; Obregon-Morales, C.; Salas-Rojas, M.; Garcia-Flores, M.M.; et al. Molecular detection of Bartonella species in blood-feeding bat flies from México. Vector Borne Zoonotic Dis. 2018, 18, 258–265. [Google Scholar] [CrossRef]
- Abundes-Gallegos, J.; Salas-Rojas, M.; Galvez-Romero, G.; Perea-Martinez, L.; Obregon-Morales, C.Y.; Morales-Malacara, J.B.; Chomel, B.B.; Stuckey, M.J.; Moreno-Sandoval, H.; Garcia-Baltazar, A.; et al. Detection of Dengue virus in bat flies (Diptera: Streblidae) of common vampire bats, Desmodus rotundus, in Progreso, Hidalgo, Mexico. Vector Borne Zoonotic Dis. 2018, 18, 70–73. [Google Scholar] [CrossRef]
- Goldberg, T.L.; Bennett, A.J.; Kityo, R.; Kuhn, J.H.; Chapman, C.A. Kanyawara virus: A novel Rhabdovirus infecting newly discovered nycteribiid bat flies infesting previously unknown pteropodid bats in Uganda. Sci. Rep. 2017, 7, 5287. [Google Scholar] [CrossRef] [Green Version]
- Bennett, A.J.; Paskey, A.C.; Ebinger, A.; Pfaff, F.; Priemer, G.; Hoper, D.; Breithaupt, A.; Heuser, E.; Ulrich, R.G.; Kuhn, J.H.; et al. Relatives of rubella virus in diverse mammals. Nature 2020, 586, 424–428. [Google Scholar] [CrossRef]
- Bennett, A.J.; Goldberg, T.L. Pteropine orthoreovirus in an Angolan soft-furred fruit bat (Lissonycteris angolensis) in Uganda dramatically expands the global distribution of an emerging bat-borne respiratory virus. Viruses 2020, 12, 740. [Google Scholar] [CrossRef]
- Binetruy, F.; Dupraz, M.; Buysse, M.; Duron, O. Surface sterilization methods impact measures of internal microbial diversity in ticks. Parasit. Vectors 2019, 12, 268. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Löytynoja, A. Phylogeny-aware alignment with PRANK. Methods Mol. Biol. 2014, 1079, 155–170. [Google Scholar]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Abascal, F.; Zardoya, R.; Telford, M. TranslatorX: Multiple alignment of nucleotide sequences guided by amino acid translations. Nucleic Acids Res. 2010, 38, W7–W13. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: Smart model selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [Green Version]
- Toohey-Kurth, K.; Sibley, S.D.; Goldberg, T.L. Metagenomic assessment of adventitious viruses in commercial bovine sera. Biol. J. Int. Assoc. Biol. Stand. 2017, 47, 64–68. [Google Scholar] [CrossRef]
- Negrey, J.D.; Thompson, M.E.; Langergraber, K.E.; Machanda, Z.P.; Mitani, J.C.; Muller, M.N.; Otali, E.; Owens, L.A.; Wrangham, R.W.; Goldberg, T.L. Demography, life-history trade-offs, and the gastrointestinal virome of wild chimpanzees. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20190613. [Google Scholar] [CrossRef]
- Richard, J.C.; Leis, E.; Dunn, C.D.; Agbalog, R.; Waller, D.; Knowles, S.; Putnam, J.; Goldberg, T.L. Mass mortality in freshwater mussels (Actinonaias pectorosa) in the Clinch River, USA, linked to a novel densovirus. Sci. Rep. 2020, 10, 14498. [Google Scholar] [CrossRef]
- Vasilakis, N.; Widen, S.; da Rosa, A.P.T.; Wood, T.G.; Walker, P.J.; Holmes, E.C.; Tesh, R.B. Malpais spring virus is a new species in the genus Vesiculovirus. Virol. J. 2013, 10, 69. [Google Scholar] [CrossRef] [Green Version]
- Tesh, R.; Saidi, S.; Javadian, E.; Loh, P.; Nadim, A. Isfahan virus, a new vesiculovirus infecting humans, gerbils, and sandflies in Iran. Am. J. Trop. Med. Hyg. 1977, 26, 299–306. [Google Scholar] [CrossRef]
- De Souza, W.M.; Acrani, G.O.; Romeiro, M.F.; Júnior, O.R.; Tolardo, A.L.; de Andrade, A.A.; da Silva Gonçalves Vianez Júnior, J.L.; de Almeida Medeiros, D.B.; Nunes, M.R.; Figueiredo, L.T. Complete genome sequence of Piry vesiculovirus. Arch. Virol. 2016, 161, 2325–2328. [Google Scholar] [CrossRef]
- Sapkal, G.N.; Sawant, P.M.; Mourya, D.T. Chandipura viral encephalitis: A brief review. Open Virol. J. 2018, 12, 44–51. [Google Scholar] [CrossRef]
- Rodriguez, L.L. Emergence and re-emergence of vesicular stomatitis in the United States. Virus Res. 2002, 85, 211–219. [Google Scholar] [CrossRef]
- Kumar, B.; Manuja, A.; Gulati, B.R.; Virmani, N.; Tripathi, B.N. Zoonotic viral diseases of equines and their impact on human and animal health. Open Virol. J. 2018, 12, 80–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maclachlan, N.J.; Zientara, S.; Wilson, W.C.; Richt, J.A.; Savini, G. Bluetongue and epizootic hemorrhagic disease viruses: Recent developments with these globally re-emerging arboviral infections of ruminants. Curr. Opin. Virol. 2019, 34, 56–62. [Google Scholar] [CrossRef]
- Alkhamis, M.A.; Aguilar-Vega, C.; Fountain-Jones, N.M.; Lin, K.; Perez, A.M.; Sanchez-Vizcaino, J.M. Global emergence and evolutionary dynamics of bluetongue virus. Sci. Rep. 2020, 10, 21677. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Mohd Jaafar, F. Zoonotic and emerging orbivirus infections. Rev. Sci. Tech. 2015, 34, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Ruder, M.G.; Lysyk, T.J.; Stallknecht, D.E.; Foil, L.D.; Johnson, D.J.; Chase, C.C.; Dargatz, D.A.; Gibbs, E.P. Transmission and epidemiology of bluetongue and epizootic hemorrhagic disease in North America: Current perspectives, research gaps, and future directions. Vector Borne Zoonotic Dis. 2015, 15, 348–363. [Google Scholar] [CrossRef]
- Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef]
- Evans, A.B.; Peterson, K.E. Throw out the map: Neuropathogenesis of the globally expanding California serogroup of orthobunyaviruses. Viruses 2019, 11, 794. [Google Scholar] [CrossRef] [Green Version]
- Edridge, A.W.D.; van der Hoek, L. Emerging orthobunyaviruses associated with CNS disease. PLoS Negl. Trop. Dis. 2020, 14, e0008856. [Google Scholar] [CrossRef]
- Hughes, H.R.; Adkins, S.; Alkhovskiy, S.; Beer, M.; Blair, C.; Calisher, C.H.; Drebot, M.; Lambert, A.J.; de Souza, W.M.; Marklewitz, M.; et al. ICTV virus taxonomy profile: Peribunyaviridae. J. Gen. Virol. 2020, 101, 1–2. [Google Scholar] [CrossRef]
- Tomori, O.; Monath, T.P.; Lee, V.; Fagbami, A.; Fabiyi, A. Bwamba virus infection: A sero-survey of veterbrates in five ecological zones in Nigeria. Trans. R. Soc. Trop. Med. Hyg. 1974, 68, 461–465. [Google Scholar] [CrossRef]
- Kalunda, M.; Lwanga-Ssozi, C.; Lule, M.; Mukuye, A. Isolation of Chikungunya and Pongola viruses from patients in Uganda. Trans. R. Soc. Trop. Med. Hyg. 1985, 79, 567. [Google Scholar] [CrossRef]
- Shope, R.E.; de Andrade, A.H.; Bensabath, G.; Causey, O.R.; Humphrey, P.S. The epidemiology of EEE WEE, SLE and turlock viruses, with special reference to birds, in a tropical rain forest near Belem, Brazil. Am. J. Epidemiol. 1966, 84, 467–477. [Google Scholar] [CrossRef]
- Schnagl, R.D.; Holmes, I.H. Electron microscopy of Japanaut and Tilligerry viruses: Two proposed members of the orbivirus group. Aust. J. Biol. Sci. 1975, 28, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Kemp, G.E.; Le Gonidec, G.; Karabatsos, N.; Rickenbach, A.; Cropp, C.B. IFE: A new African orbivirus isolated from Eidolon helvum bats captured in Nigeria, Cameroon and the Central African Republic. Bull. Soc. Pathol. Exot. Fil. 1987, 81, 40–48. [Google Scholar]
- Boiro, I.; Fidarov, F.M.; Lomonossov, N.N.; Linev, M.B.; Bachkirsov, V.N.; Inapogui, A. Isolation of the Fomédé virus from Chiroptera, Nycteris nana, in the Republic of Guinea. Bull. Soc. Pathol. Exot. Fil. 1985, 79, 180–182. [Google Scholar]
- Fagre, A.C.; Lee, J.S.; Kityo, R.M.; Bergren, N.A.; Mossel, E.C.; Nakayiki, T.; Nalikka, B.; Nyakarahuka, L.; Gilbert, A.T.; Peterhans, J.K.; et al. Discovery and characterization of Bukakata orbivirus (Reoviridae:Orbivirus), a novel virus from a Ugandan bat. Viruses 2019, 11, 209. [Google Scholar] [CrossRef] [Green Version]
- Miura, T.; Kitaoka, M. Viruses isolated from bats in Japan. Arch. Virol. 1977, 53, 281–286. [Google Scholar] [CrossRef]
- Justines, G.; Kuns, M.L. Matucare virus, a new agent recovered from Ornithodoros (Alectorobius) boliviensis. Am. J. Trop. Med. Hyg. 1970, 19, 697–702. [Google Scholar] [CrossRef]
- Dick, C.W.; Gettinger, D.; Gardner, S.L. Bolivian ectoparasites: A survey of bats (Mammalia: Chiroptera). Comp. Parasitol. 2007, 74, 372–377. [Google Scholar] [CrossRef]
- Whitaker, J.O.J.; Ritzi, C.M.; Dick, C.W. Collecting and preserving ectoparasites for ecological study. In Ecological and Behavioral Methods for the Study of Bats, 2nd ed.; Kunz, T.H., Parsons, S., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2009; pp. 806–827. [Google Scholar]
- Reeves, W.R.; Lloyd, J.E. Louse Flies, Keds, and Bat Flies (Hippoboscoidea). In Medical and Veterinary Entomology, 3rd ed.; Mullen, G.R., Durden, L.A., Eds.; Academic Press: London, UK, 2019; pp. 421–438. [Google Scholar]
- Osborne, J.C.; Rupprecht, C.E.; Olson, J.G.; Ksiazek, T.G.; Rollin, P.E.; Niezgoda, M.; Goldsmith, C.S.; An, U.S.; Nichol, S.T. Isolation of Kaeng Khoi virus from dead Chaerephon plicata bats in Cambodia. J. Gen. Virol. 2003, 84, 2685–2689. [Google Scholar] [CrossRef]
- Williams, J.E.; Imlarp, S.; Top, F.H., Jr.; Cavanaugh, D.C.; Russell, P.K. Kaeng Khoi virus from naturally infected bedbugs (Cimicidae) and immature free-tailed bats. Bull. World Health Organ. 1976, 53, 365–369. [Google Scholar]
- Groseth, A.; Mampilli, V.; Weisend, C.; Dahlstrom, E.; Porcella, S.F.; Russell, B.J.; Tesh, R.B.; Ebihara, H. Molecular characterization of human pathogenic bunyaviruses of the Nyando and Bwamba/Pongola virus groups leads to the genetic identification of Mojui dos Campos and Kaeng Khoi virus. PLoS Negl. Trop. Dis. 2014, 8, e3147. [Google Scholar] [CrossRef]
- Pinheiro, F.P.; Travassos da Rosa, A.P. Mojui dos Campos. In International Catalogue of Arboviruses Including Certain Other Viruses of Vertebrates, 3rd ed.; Karabatsos, N., Ed.; American Society of Tropical Medicine and Hygiene: San Antonio, TX, USA, 1985; pp. 695–696. [Google Scholar]
- Zhang, Y.Z.; Shi, M.; Holmes, E.C. Using metagenomics to characterize an expanding virosphere. Cell 2018, 172, 1168–1172. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Z.; Chen, Y.M.; Wang, W.; Qin, X.C.; Holmes, E.C. Expanding the RNA virosphere by unbiased metagenomics. Annu. Rev. Virol. 2019, 6, 119–139. [Google Scholar] [CrossRef]
- Téllez, H.L.A.; Iñiguez-Dávalos, L.I.; Olvera-Vargas, M.; Vargas-Contreras, J.A.; Herrera-Lizaola, O.A. Bats associated to caves in Jalisco, Mexico. THERYA 2018, 9, 29–40. [Google Scholar]
- Horton, D.L.; Breed, A.C.; Arnold, M.E.; Smith, G.C.; Aegerter, J.N.; McElhinney, L.M.; Johnson, N.; Banyard, A.C.; Raynor, R.; Mackie, I.; et al. Between roost contact is essential for maintenance of European bat lyssavirus type-2 in Myotis daubentonii bat reservoir: ‘The Swarming Hypothesis’. Sci. Rep. 2020, 10, 1740. [Google Scholar]
- Parratt, S.R.; Laine, A.L. The role of hyperparasitism in microbial pathogen ecology and evolution. ISME J. 2016, 10, 1815–1822. [Google Scholar] [CrossRef] [Green Version]
- Haelewaters, D.; Hiller, T.; Dick, C.W. Bats, bat flies, and fungi: A case of hyperparasitism. Trends Parasitol. 2018, 34, 784–799. [Google Scholar]
- Wenzel, R.L.; Tipton, V.J. Some relationships between mammal hosts and their ectoparasites. In Ectoparasites of Panama; Wenzel, R.L., Tipton, V.J., Eds.; Field Museum of Natural History: Chicago, IL, USA, 1966; pp. 677–723. [Google Scholar]
- Watts, D.M.; Pantuwatana, S.; DeFoliart, G.R.; Yuill, T.M.; Thompson, W.H. Transovarial transmission of LaCrosse virus (California encephalitis group) in the mosquito, Aedes triseriatus. Science 1973, 182, 1140–1141. [Google Scholar] [CrossRef]
- Rozo-Lopez, P.; Drolet, B.S.; Londoño-Renteria, B. Vesicular stomatitis virus transmission: A comparison of incriminated vectors. Insects 2018, 9, 190. [Google Scholar] [CrossRef] [Green Version]
- Tesh, R.B.; Boshell, J.; Modi, G.B.; Morales, A.; Young, D.G.; Corredor, A.; de Carrasquilla, C.F.; de Rodriguez, C.; Walters, L.L.; Gaitan, M.O. Natural infection of humans, animals, and phlebotomine sand flies with the Alagoas serotype of vesicular stomatitis virus in Colombia. Am. J. Trop. Med. Hyg. 1987, 36, 653–661. [Google Scholar] [CrossRef]
- Corn, J.L.; Comer, J.A.; Erickson, G.A.; Nettles, V.F. Isolation of vesicular stomatitis virus New Jersey serotype from phlebotomine sand flies in Georgia. Am. J. Trop. Med. Hyg. 1990, 42, 476–482. [Google Scholar] [CrossRef]
- Comer, J.A.; Corn, J.L.; Stallknecht, D.E.; Landgraf, J.G.; Nettles, V.F. Titers of vesicular stomatitis virus, New Jersey serotype, in naturally infected male and female Lutzomyia shannoni (Diptera: Psychodidae) in Georgia. J. Med. Entomol. 1992, 29, 368–370. [Google Scholar] [CrossRef]
- Comer, J.A.; Tesh, R.B.; Modi, G.B.; Corn, J.L.; Nettles, V.F. Vesicular stomatitis virus, New Jersey serotype: Replication in and transmission by Lutzomyia shannoni (Diptera: Psychodidae). Am. J. Trop. Med. Hyg. 1990, 42, 483–490. [Google Scholar] [CrossRef]
- Schmidtmann, E.T.; Craig, M.E.; English, L.M.; Herrero, M.V. Sampling for sand flies (Diptera: Psychodidae) among prairie dog colonies on ranches with histories of vesicular stomatitis in new Mexico and Colorado. J. Med. Entomol. 2002, 39, 680–684. [Google Scholar] [CrossRef] [Green Version]
- Fleming, T.H. Bat Migration. Encycl. Anim. Behav. 2019, 605–610. [Google Scholar] [CrossRef]
- Wiederholt, R.; Lopez-Hoffman, L.; Cline, J.; Medellín, R.A.; Cryan, P.; Russell, A.; McCracken, G.; Diffendorfer, J.; Semmens, D. Moving across the border: Modeling migratory bat populations. Ecosphere 2013, 4, 114. [Google Scholar] [CrossRef]
- Clark, G.G.; Calisher, C.H.; Crabbs, C.L.; Canestorp, K.M.; Tesh, R.B.; Bowen, R.A.; Taylor, D.E. Malpais spring virus: A new vesiculovirus from mosquitoes collected in New Mexico and evidence of infected indigenous and exotic ungulates. Am. J. Trop. Med. Hyg. 1988, 39, 586–592. [Google Scholar] [CrossRef]
- Huang, T.; Liu, Z.; Gong, X.; Wu, T.; Liu, H.; Deng, J.; Zhang, Y.; Peng, Q.; Zhang, L.; Liu, Z. Vampire in the darkness: A new genus and species of land leech exclusively bloodsucking cave-dwelling bats from China (Hirudinda: Arhynchobdellida: Haemadipsidae). Zootaxa 2019, 4560, 257–272. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Virus 1 | Accession | Genome | Closest Match 2 | Family 3 | Genus 3 | % Identity 4 |
|---|---|---|---|---|---|---|---|
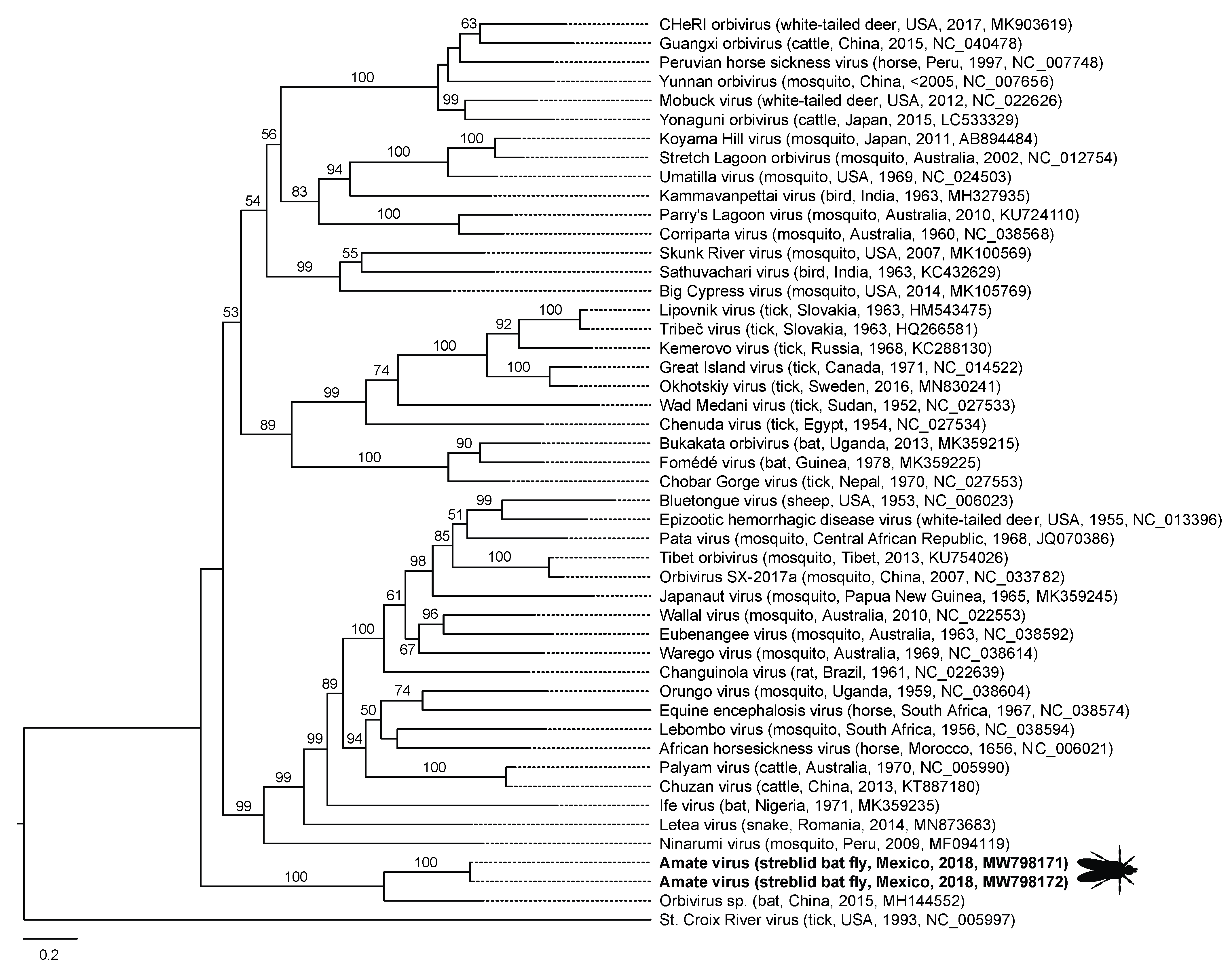
| México | Amate virus | MW798171MW798172 | dsRNA, segmented | Orbivirus sp (Taphozous melanopogon bat, China, 2018, MH144552) | Reoviridae | Orbivirus | 68.65 (78, 0.0) |
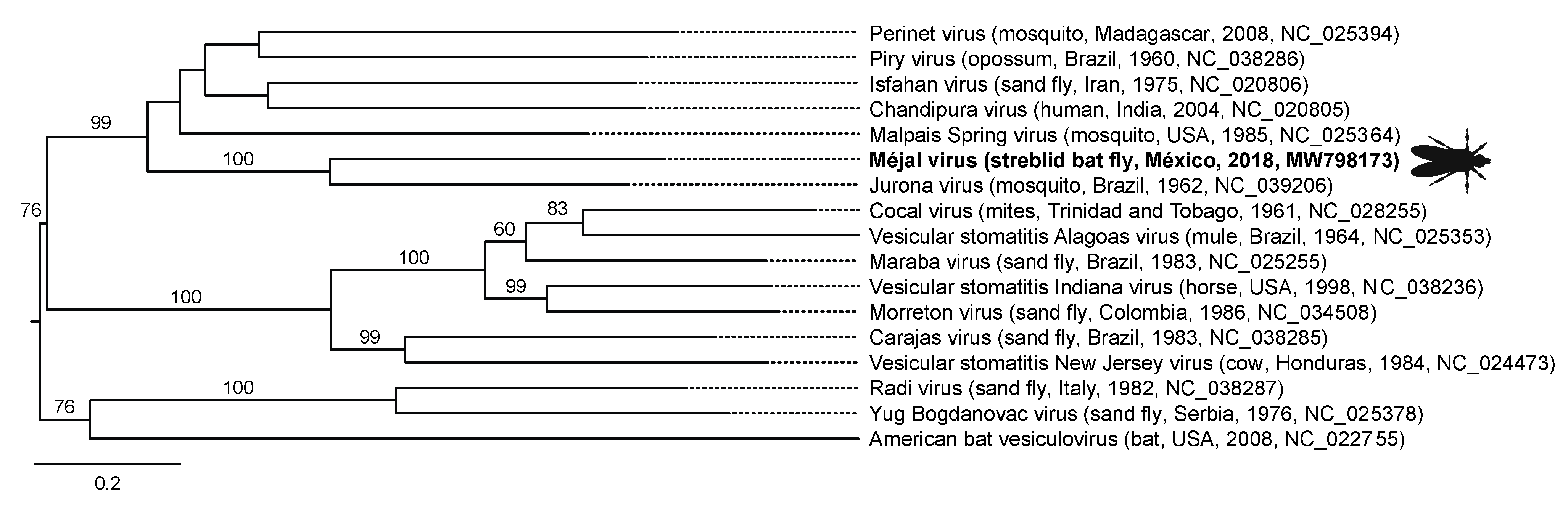
| México | Méjal virus | MW798173 | −ssRNA, linear | Jurona virus (Haemagogus sp. mosquito, Brazil, 1962, NC_039206) | Rhabdoviridae | Vesiculovirus | 68.60 (91, 0.0) |
| México | Jalimé iflavirus 1 | MW798174 | +ssRNA, linear | Varroa destructor virus 2 (Varroa destructor mite, Israel, 2014, NC_040601) | Iflaviridae | unclassified | 72.04 (42, 3e-53) |
| México | Jalimé partiti-like virus 1 | MW798175 | dsRNA | Wuhan insect virus 24 (unspecified insects, China, 2013, KX884086) | unclassified | unclassified | 68.42 (85, 7e-109) |
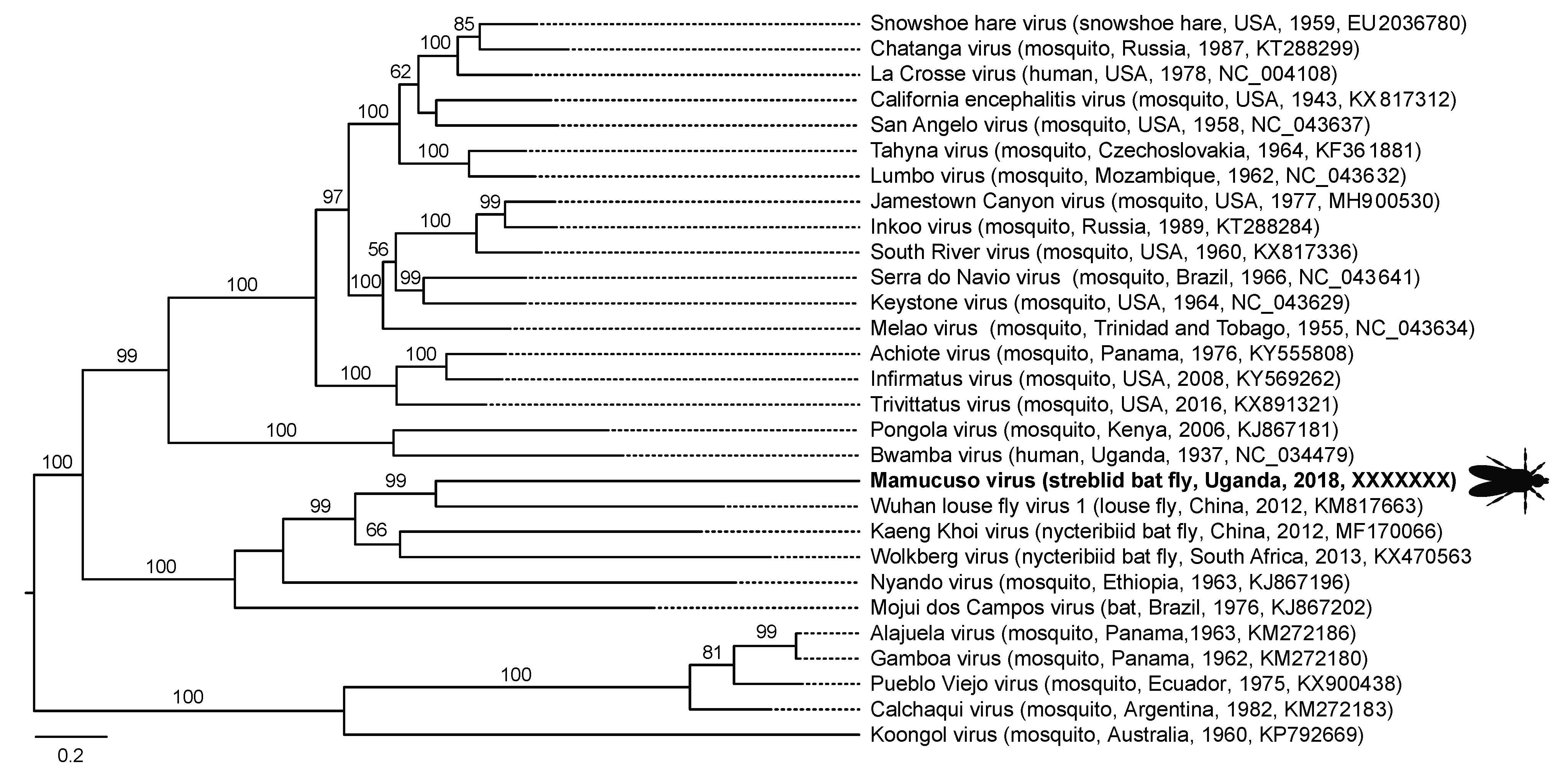
| Uganda | Mamucuso virus | MW798176 | −ssRNA, segmented | Wuhan Louse Fly Virus 1 (unidentified Hippoboscoidea, China, 2012, KM817663) | Peribunyaviridae | Orthobunyavirus | 69.45 (96, 0.0) |
| Uganda | Kamu rhabdovirus 1 | MW798177 | −ssRNA, linear | American bat vesiculovirus (Eptesicus fuscus bat, USA, 2008, JX569193) | Rhabdoviridae | unclassified | 72.90 (22, 9e-13) |
| Uganda | Kamu toti-like virus 1 | MW798178 | dsRNA | Hubei toti-like virus 22 (unspecified insects, China, 2013, KX882954) | unclassified | unclassified | 76.79 (5, 2e-9) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Martínez, M.M.; Bennett, A.J.; Dunn, C.D.; Yuill, T.M.; Goldberg, T.L. Bat Flies of the Family Streblidae (Diptera: Hippoboscoidea) Host Relatives of Medically and Agriculturally Important “Bat-Associated” Viruses. Viruses 2021, 13, 860. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050860
Ramírez-Martínez MM, Bennett AJ, Dunn CD, Yuill TM, Goldberg TL. Bat Flies of the Family Streblidae (Diptera: Hippoboscoidea) Host Relatives of Medically and Agriculturally Important “Bat-Associated” Viruses. Viruses. 2021; 13(5):860. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050860
Chicago/Turabian StyleRamírez-Martínez, María M., Andrew J. Bennett, Christopher D. Dunn, Thomas M. Yuill, and Tony L. Goldberg. 2021. "Bat Flies of the Family Streblidae (Diptera: Hippoboscoidea) Host Relatives of Medically and Agriculturally Important “Bat-Associated” Viruses" Viruses 13, no. 5: 860. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050860